Dynamics of natural regeneration in Pinus laricio stands from southern Apennines (Italy)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 3, Pages 380-386 (2006)
doi: https://doi.org/10.3832/efor0401-0030380
Published: Sep 20, 2006 - Copyright © 2006 SISEF
Research Articles
Guest Editors: 5° SISEF Congress (Grugliasco, TO - 2005)
« Forests and Society - Changes, Conflicts, Sinergies »
Collection/Special Issue: E. Lingua, R. Marzano, G. Minotta, R. Motta, A. Nosenzo, G. Bovio
Abstract
The seedlings establishment was studied in small (380 m2), medium (855 m2) and large (1520 m2) gaps created in calabrian pine (Pinus laricio Poiret) stands (mean height 22 m) in the Southern Apennine. After three growing seasons first results put in evidence: a) no significant differences of water soil content were observed between gap sizes; b) transmittance was higher in large and medium gaps than in small ones and in the centre and northern sides in both gap sizes; c) calabrian pine seedling density was higher in large gaps than in medium and small ones and namely in the centre positions; silver fir seedlings appear after the second growing season in small and medium gaps; d) seedling mortality of calabrian pine was relevant in small and medium gaps in the edge and silver fir seedling mortality in the centre of the large ones; e) in these first years the ground vegetation (bramble and bracken) represents a moderate detrimental effect on seedlings establishment that could be removed by partial cuttings.
Keywords
Renaturalization, Regeneration, Conifer stand, Gaps, Microclimate
Introduzione
In Italia sono stati eseguiti numerosi rimboschimenti con conifere, la maggior parte con pini, a scopo essenzialmente protettivo. Molti di questi rimboschimenti mostrano gravi problemi di stabilità (meccanica e biologica) di fronte agli agenti meteorici (venti, tempeste) e suscettibilità ad attacchi di funghi patogeni e insetti.
La rinaturalizzazione delle monocolture di conifere (qui intesa come la trasformazione in popolamenti con una maggiore diversità compositiva e strutturale) rappresenta uno dei principali obiettivi della selvicoltura in Italia e in molti Paesi europei ([12], [18], [14], [6], [28], [17]), soprattutto all’interno delle aree protette.
Tra i vari strumenti selvicolturali vengono sempre più presi in considerazione quelli basati sul principio che l’evoluzione naturale delle foreste è legata all’apertura di vuoti (buche, gaps) nella copertura arborea per la morte di uno o più individui ([25], [21], [27], [23], [19]). L’apertura di vuoti nella copertura arborea consente inoltre di incrementare la diversità specifica ([5], [24]).
Gugliotta & Mercurio ([11]) hanno dimostrato che l’apertura di buche in rimboschimenti di pino nero in Abruzzo consente l’insediamento di numerose latifoglie e quindi di avviare dinamiche evolutive verso popolamenti misti. Il ruolo delle dimensioni delle buche e dei fattori ecologici sull’insediamento di specie forestali è stato studiato in molte foreste ([9], [10], [16], [3], [4], [14], [6], [29]).
Lo scopo di questo lavoro è di: 1) valutare se l’applicazione del taglio a buche rappresenta una via appropriata per favorire la rinnovazione naturale di altre specie oltre il pino laricio; 2) analizzare l’influenza dei fattori microambientali in buche di differenti dimensioni sulla rinnovazione naturale e quindi determinare le dimensioni ottimali.
Materiali e metodi
L’area sperimentale è stata realizzata in una pineta artificiale di pino laricio (Pinus laricio Poir.) in località Bufalaria, Comune di Capistrano (VV) sul macroversante est delle Serre vibonesi (Appennino meridionale ? 38°42’N, 16°20’E) a 900 m, con pendenza del terreno del 5%.
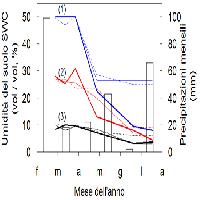
La precipitazione media annua è di 1848 mm, quella estiva di 102 mm, la temperatura media annua è di 10.8 °C, quella del mese più freddo di 3.1 °C, quella del mese più caldo di 21.5 °C. I suoli classificati come Humic Dystrudept, sono profondi, a tessitura sabbioso-franca e derivano da gneiss e scisti biotitici. La vegetazione naturale potenziale è costituita da faggete (Anemono apenninae-Fagetum Brullo 1984 = Aquifolio-Fagetum Gentile 1969) progressivamente sostituite in passato da colture agricole; l’area è stata poi rimboschita negli anni ’50.
Durante la primavera 2003 sono stati realizzati tagli a buche di forma circolare in condizioni di sostanziale omogeneità per quanto riguardava altitudine, esposizione e pendenza. Sono state considerate le seguenti dimensioni: tagli a buche "piccole" (380 m2), "medie" (855 m2) e "grandi" (1520 m2) con due repliche ciascuna. Il rapporto tra il diametro della buca e l’altezza delle piante più alte (D/H) era rispettivamente di 1.0, 1.5 e 2.0 (altezza media di 22 m). Dopo le operazioni di taglio e esbosco è stato rimosso l’orizzonte Oi del suolo.
All’interno di ogni buca sono state realizzate 9 microaree circolari di 3.14 m2 (raggio 100 cm) posizionate una nel centro della buca e le altre nelle quattro direzioni cardinali, alla distanza di un terzo del raggio, partendo dal centro della buca, per campionare le posizioni CENTRO, e, quattro, sempre nelle quattro direzioni cardinali, in prossimità dell’area di insidenza della chioma delle piante al limite della buca per campionare le posizioni di ORLO. Ciò allo scopo di valutare quali sono le posizioni all’interno della buca più favorevoli per l’insediamento e l’affermazione delle varie specie forestali.
Misure ambientali
Umidità del suolo
Sono stati prelevati quattro campioni di suolo con una sonda pedologica (8 cm in diametro x 15 cm in profondità) nella parte centrale e nell’orlo di ogni buca (al di fuori delle microaree ma nella loro prossimità per evitare di danneggiare i semenzali) per determinare il contenuto idrico del suolo con il metodo gravimetrico. Un analogo numero di campioni di suolo è stato prelevato anche all’interno del popolamento non trattato adiacente ogni buca. I campionamenti sono stati eseguiti, mensilmente, alla distanza di 48 ore dall’ultima precipitazione. È stato campionato un solo livello di profondità in quanto i semenzali concentrano la maggior parte del proprio apparato radicale nello strato superficiale del suolo. Ogni campione è stato pesato immediatamente dopo la raccolta (peso fresco). In seguito, in laboratorio, i campioni sono stati essiccati in stufa ventilata a 105 ± 2 °C per almeno 24 ore fino a raggiungere il peso costante e quindi pesati di nuovo (peso secco). Il contenuto idrico del suolo è stato espresso come percentuale del peso secco (eqn. 1):
Temperatura del suolo
Misure della temperatura del suolo sono state eseguite, in ogni microarea, alla profondità di 10 cm, con la terra accostata al termometro per circa 1 minuto, alle ore 12.00, durante il mese di agosto ogni 10 giorni. I valori di temperatura che si raggiungono in questo mese possono infatti correlarsi alla mortalità dei semenzali.
Radiazione
Sono state eseguite misure di radiazione fotosinteticamente attiva (PAR) (radiazione 0.4 - 0.7 m della banda) con un ceptometro (AccuPAR, Degagon Devices Inc., Pullman, WA, USA), a 1.00 m da terra, con lo strumento in orizzontale. In ogni punto di campionamento sono state eseguite quattro misure istantanee secondo le direzioni cardinali.
Le misure sono state eseguite mensilmente, durante la stagione vegetativa (maggio-settembre), in giornate con cielo sereno alle 12.00 (ora solare) per rilevare i valori massimi. Le misure di radiazione sono state eseguite:
- all’aperto in una radura, in prossimità delle buche
- all’interno della pineta
- in corrispondenza di ogni microarea.
Dopo ogni ciclo di misure è stata calcolata la trasmittanza con la seguente formula (eqn. 2):
Rilievi sui semenzali
Tutti i semenzali, nati dopo l’apertura della buca, all’interno di ogni microarea sono stati identificati per specie, è stata misurata l’altezza, l’etàè stata determinata mediante il conteggio dei verticilli. Il censimento dei semenzali in ogni microarea è stato eseguito alla fine di ogni stagione vegetativa. Sono stati quindi calcolati: densità, altezza media, età, mortalità.
La percentuale di copertura della vegetazione erbacea e arbustiva è stata stimata a livello di microarea e di buca.
Le differenze tra le diverse dimensioni delle buche (piccole, medie, grandi) e la posizione all’interno di ogni buca rispetto ai vari parametri microambientali e la rinnovazione sono state testate per verificarne la significatività statistica con analisi della varianza per ranghi a un criterio di classificazione con il test di Kruskal-Wallis (Sigmastat 3.1, SYSTAT software Inc., USA).
Risultati e discussione
Vengono esposti i risultati relativi alla terza stagione vegetativa (2005), mentre per quanto riguarda la mortalità si riportano anche quelli della seconda stagione vegetativa (2004).
Umidità del suolo
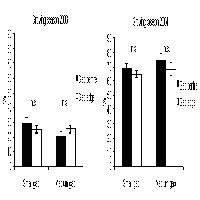
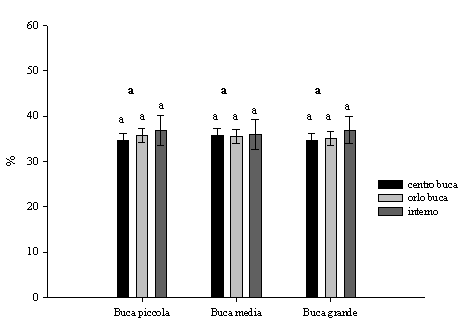
Le differenze del contenuto idrico del suolo tra centro e orlo non sono statisticamente significative sia rispetto alle diverse dimensioni che alle posizioni delle buche (Fig. 1). Anche McGuire et al. ([15]) in buche su pinete di Pinus palustris dove il rapporto D/H variava da 0.75 e 3 non hanno osservato differenze tra centro e orlo. I valori rilevati all’interno del bosco sono stati superiori in confronto con le buche ma non sono stati statisticamente significativi (Fig. 1).
Fig. 1 - Variazioni dell’umidità del suolo in relazione alle dimensioni delle buche, alla posizione all’interno delle buche e all’interno del bosco (± ES). Lettere differenti indicano variazioni significative (p<0.05), quelle in grassetto si riferiscono alle dimensioni delle buche, quelle in carattere normale sono relative alle diverse posizioni.
Trasmittanza
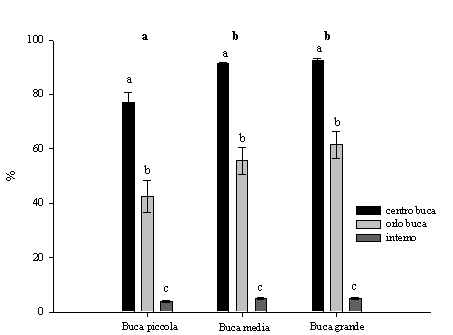
La trasmittanza è stata più elevata nelle buche grandi e medie rispetto a quelle piccole. Le differenze non sono significative tra le buche medie e grandi ma lo sono rispetto alle piccole, i valori abbastanza simili che sono stati rilevati nelle buche grandi e medie e in minor misura in quelle piccole sono dovuti al campionamento della radiazione nell’ora centrale del giorno. I valori sono più elevati e statisticamente significativi nel centro rispetto all’orlo in tutte le dimensioni analizzate (Fig. 2). Così come osservato da McGuire et al. ([15]) in buche dove il rapporto D/H variava da 0.75 e 3. I valori rilevati all’interno del bosco sono decisamente inferiori rispetto a quelli rilevati all’interno delle buche (Fig. 2).
Fig. 2 - Variazioni della trasmittanza (h 12.00, ± ES) in relazione alle dimensioni delle buche, alla posizione all’interno delle buche e all’interno del bosco. Lettere differenti indicano variazioni significative (p<0.05), quelle in grassetto si riferiscono alle dimensioni delle buche, quelle in carattere normale sono relative alle diverse posizioni.
Temperatura del suolo
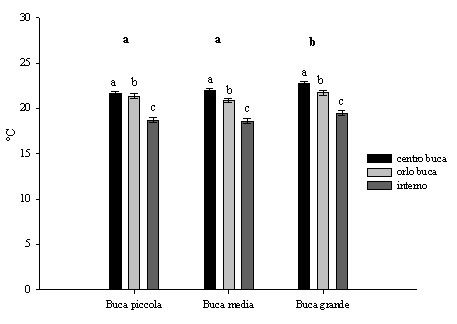
La temperatura del suolo non raggiunge valori elevati (< 25 °C), come era da aspettarsi in suoli a tessitura sabbioso-franca, in quanto mitigata dalla copertura erbacea e arbustiva. Quindi questo fattore non può essere direttamente associato alla mortalità dei giovani semenzali ([10]). I valori differiscono in maniera significativa tra le buche grandi rispetto a quelle medie e piccole. Inoltre sono superiori nel centro rispetto all’orlo delle buche con significative differenze in tutte le dimensioni delle buche. Analogamente sono maggiori e significativamente differenti all’interno delle buche rispetto all’interno del bosco (Fig. 3) a causa dell’incremento della radiazione.
Fig. 3 - Variazioni della temperatura del suolo (agosto, h 12.00, ± ES) in relazione alle dimensioni delle buche, alla posizione all’interno delle buche e all’interno del bosco. Lettere differenti indicano variazioni significative (p<0.05), quelle in grassetto si riferiscono alle dimensioni delle buche, quelle in carattere normale sono relative alle diverse posizioni.
Densità dei semenzali
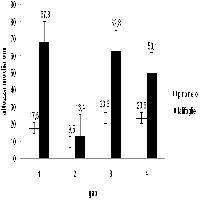
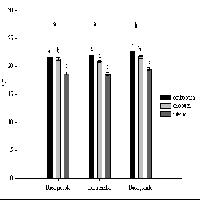
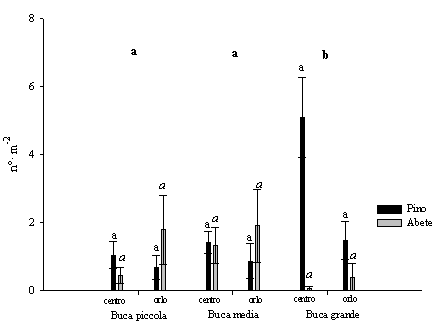
La densità dei semenzali di pino laricio è stata significativamente più elevata nelle buche grandi rispetto a quelle medie e piccole (nelle buche medie e piccole si è verificata una alta mortalità come viene specificato in seguito). I valori sono inoltre maggiori nel centro ma non statisticamente significativi rispetto all’orlo (Fig. 4). Analogo comportamento è stato riscontrato in buche naturali in popolamenti di Pinus palustris in Florida (USA) ([2]). A partire dal 2° e poi nel 3° anno si è osservato l’ingresso dei semenzali di abete bianco con densità maggiori nelle buche piccole e medie, sia nell’orlo che nel centro (Fig. 4). Confermando un comportamento su quanto osservato in precedenti esperienze sui tagli a buche in abetine artificiali nell’Appennino meridionale ([1]). Nella prima stagione vegetativa la rinnovazione naturale era costituita, in tutte le buche, dal 100% di semenzali di pino laricio dell’anno. Nella terza stagione vegetativa diminuisce sensibilmente il numero dei semenzali di pino laricio di 1 anno e aumenta considerevolmente quello dei semenzali di abete bianco che potrebbe indicare che si stanno creando condizioni più favorevoli per quest’ultima specie (Tab. 1). Infatti i semenzali di abete bianco possono insediarsi e sopravvivere anche a bassi livelli radiativi ([20], [22], [13]) sotto parziale copertura delle erbe.
Fig. 4 - Variazioni della densità dei semenzali (± ES) in relazione alle dimensioni delle buche e alla posizione all’interno delle buche. Lettere differenti indicano variazioni significative (p<0.05), quelle in grassetto si riferiscono alle dimensioni delle buche (limitatamente al pino), quelle in carattere normale sono relative al pino nelle diverse posizioni, quelle in corsivo sono relative all’abete nelle diverse posizioni.
Tab. 1 - Distribuzione percentuale dei semenzali di pino laricio e di abete bianco in relazione alle dimensioni delle buche, alla specie e all’età.
| Anno | Età | Buca piccola | Buca media | Buca grande | |||
|---|---|---|---|---|---|---|---|
| Pino | Abete | Pino | Abete | Pino | Abete | ||
| 2004 | 1 anno | 62 | 100 | 66 | 100 | 74 | 100 |
| 2004 | 2 anni | 38 | - | 34 | - | 26 | - |
| 2005 | 1 anno | 6 | 57 | 5 | 58 | 1 | 46 |
| 2005 | 2 anni | 47 | 43 | 56 | 42 | 71 | 54 |
| 2005 | 3 anni | 47 | - | 39 | - | 28 | - |
Mortalità dei semenzali
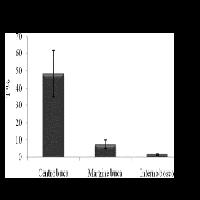
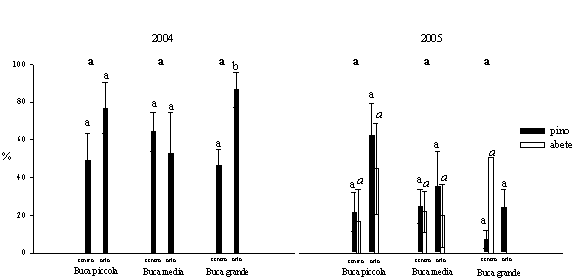
La mortalità dei semenzali di pino laricio diminuisce nel 2005 rispetto al 2004; è stata maggiore nelle buche medie e piccole rispetto a quelle grandi ma senza significative differenze. Inoltre è stata più rilevante nell’orlo rispetto al centro verosimilmente dovuta all’ombreggiamento delle chiome degli alberi circostanti. La mortalità dei semenzali di abete bianco assume invece valori più rilevanti nel centro delle buche grandi (Fig. 5).
Fig. 5 - Variazioni della mortalità dei semenzali (± ES) in relazione alle dimensioni delle buche e alla posizione all’interno delle buche. Lettere differenti indicano variazioni significative (p<0.05), quelle in grassetto si riferiscono alle dimensioni delle buche (limitatamente al pino), quelle in carattere normale sono relative al pino nelle diverse posizioni, quelle in corsivo sono relative all’abete nelle diverse posizioni.
Copertura erbacea e arbustiva
La copertura della vegetazione all’interno delle buche è in media dell’80% (a prevalenza di rovo e felce aquilina), valori leggermente inferiori (65%) si hanno nelle microaree poste nell’orlo. Malcolm et al. ([14]) ritengono che il numero dei semenzali può essere più basso dove rovo e felce aquilina sono dominanti. In particolare la felce aquilina può svolgere un’azione di competizione per la luce, l’umidità e i nutrienti, e può avere un effetto fitotossico sulla germinazione e il primo insediamento nei semenzali di pino silvestre ([7]). Il rovo svolge il ruolo di specie miglioratrice del suolo, un’azione complessa sui semenzali sia di protezione che di competizione ([26]), ma i semenzali di abete possono comunque attraversare la copertura del rovo ([8]). In questo caso, la vegetazione erbacea e arbustiva non impedisce in senso assoluto lo sviluppo dei semenzali. Del resto Gray & Spies ([10]) hanno rilevato che in buche con rapporto D/H di 1.0 Abies amabilis può insediarsi anche con un grado di copertura del 90%.
Considerazioni conclusive
I risultati dopo tre stagioni vegetative non consentono di trarre ancora delle conclusioni circa la dinamica della rinnovazione. Subito dopo l’apertura delle buche ha prevalso la rinnovazione del pino laricio, in particolar modo nel centro delle buche grandi, dopodiché è cominciato l’insediamento progressivo dell’abete bianco. Il pino è probabilmente destinato a scomparire con il tempo soprattutto nelle buche piccole e medie, mentre le specie arboree dell’Anemono apenninae-Fagetum = Aquifolio-Fagetum sono destinate ad avere, in seguito, un ruolo più rilevante. Il parziale ombreggiamento di erbe e arbusti può facilitare l’insediamento dei semenzali durante le estati siccitose e provvedere micrositi umidi e temperati più favorevoli per i semenzali delle specie tolleranti bassi livelli radiativi. In ogni caso la parziale eliminazione della copertura erbacea e arbustiva può con il tempo favorire l’affermazione dei semenzali.
Allo stesso modo non è ancora possibile stabilire quali siano le dimensioni ottimali delle buche per assicurare la definitiva affermazione delle varie specie. Tuttavia alcune considerazioni possono aiutare a dare una prima indicazione: le buche grandi con rapporto D/H (2.0) favoriscono l’affermazione del pino laricio ([14]), ma gli attacchi di Traumatocampa pytiocampa Den et Schiff. sono maggiormente frequenti negli alberi di margine nelle buche grandi mentre sono quasi inesistenti in quelle medie e piccole in quanto questo insetto preferisce le posizioni illuminate. La rinnovazione delle specie autoctone (in questo caso abete bianco) sembra essere favorita dalle buche di dimensioni più piccole. Ciò potrebbe far pensare che, nei casi come quello esaminato, potrebbero essere più appropriate le buche di piccole dimensioni se l’obiettivo è la rinaturalizzazione dei rimboschimenti.
Ringraziamenti
Questo lavoro è stato eseguito con fondi della Regione Calabria nell’ambito del progetto “La rinaturalizzazione dei rimboschimenti”. Si ringrazia il Dott. Antonio Poletto dell’Azienda Agricola S. Maria di Serra San Bruno (VV) per il supporto logistico.
References
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar