Role of soil fauna in the spatial and temporal variation of humus forms: micromorphological investigation on thin sections and stereoscopic observation
Forest@ - Journal of Silviculture and Forest Ecology, Volume 3, Pages 555-561 (2006)
doi: https://doi.org/10.3832/efor0411-0030555
Published: Dec 18, 2006 - Copyright © 2006 SISEF
Research Articles
Guest Editors: 5° SISEF Congress (Grugliasco, TO - 2005)
« Forests and Society - Changes, Conflicts, Sinergies »
Collection/Special Issue: E. Lingua, R. Marzano, G. Minotta, R. Motta, A. Nosenzo, G. Bovio
Abstract
In a spruce forest of the Autonomous Province of Trento, located on an acid substrate and with a north exposition, within the Village of Pellizzano, a transect has been traced; along this transect 19 humus profiles have been examined, one at every three meters. For each of them, OH and A horizons (respectively the humic organic and the organo-mineral horizon) have been sampled and studied following the methodology formulated by Ponge ([8]) and Bernier & Ponge ([1]). The morphological and semi-quantitative observation of these horizons with the stereoscope and the use of an identification key of soil fauna’s faecal pellets ([3]) allowed us to determine and point out their relative abundance and to formulate interesting remarks on spatial and temporal variability of humus forms in forests. For four profiles of the transect, a micro-morphological study, with the microscope, of the thin sections of every horizon has been carried out as well, in order to observe those more detailed characters such as: the presence of fossilized coprolites, of paleo-aggregates and of those soil fauna’s dung, in particular mites’ faecal pellets, hardly identifiable with the stereoscope. Both methodology of morphological investigation proved to be necessary for an accurate research and to be a substantial aid for a better identification of humus forms, after they have been described in field following the French morphological-genetic approach ([5]).
Keywords
Pedofauna, Forme di humus, Indagine morfologica, Sezioni sottili
Introduzione
Durante gli ultimi tre decenni la letteratura specialistica ha migliorato la conoscenza e la considerazione della funzione degli organismi animali nel suolo: da agenti decompositori in grado di facilitare l’assunzione di nutrienti da parte dei vegetali, hanno assunto il ruolo di veri protagonisti nella costituzione, stabilità ed evoluzione dell’ecosistema forestale.
Nella parte superficiale del suolo, i processi di trasformazione delle sostanze vegetali in humus, attraverso la digestione animale, erano già noti a Müller ([7]) ma solamente Kubiena ([6]) ha potuto confermare queste ricerche su base microscopica.
Quando i resti vegetali passano nell’intestino degli animali del suolo subiscono una demolizione fisica (sminuzzamento) ed una degradazione chimica ad opera degli enzimi intestinali. Negli strati organici ed organo-minerali, i principali protagonisti dell’opera di trasformazione fisica e chimica dei residui vegetali ed animali sono gli organismi decompositori: pedofauna, microflora e funghi si alternano e si susseguono svolgendo le azioni fondamentali di frammentazione, decomposizione e sintesi ([4]). Gli escrementi prodotti dalla pedofauna possono rappresentare sia un aumento della superficie vegetale aggredibile dalle componenti fungina e batterica, sia il primo stadio di alterazione chimica necessaria alla formazione delle sostanze umiche.
Non tutti gli animali però svolgono nel suolo la stessa funzione: alcuni di essi sono specializzati, ad esempio, nella frammentazione della necromassa vegetale, mentre altri sono deputati all’incorporazione più o meno intima dei composti organici con le particelle minerali.
Partendo da questo presupposto, l’approccio morfologico-genetico francese ([5]) descrive i Mull come forme di humus caratterizzate dalla presenza un orizzonte A biostrutturato, ovvero formato dall’attività di lombrichi, nella maggior parte dei casi anecici, che creano un intimo legame tra componente organica e componente minerale I Moder invece derivano principalmente dall’attività degli enchitreidi e l’orizzonte A presenta una struttura giustapposta, ovvero le particelle minerali sono solamente accostate agli escrementi organici, in tal caso la formazione di complessi umo-argillosi è fortemente rallentata.
La ricerca proposta rappresenta uno degli aspetti indagati nel progetto DINAMUS “Forme di humus e dinamica del bosco” ed ha lo scopo di individuare, all’interno di una pecceta altimontana della Provincia Autonoma di Trento, i principali tipi di pedofauna attivi nella creazione degli humus forestali in quattro diverse fasi di sviluppo del bosco e di evidenziarne la variabilità spaziale e temporale. L’analisi morfologica allo stereoscopio e l’osservazione micromorfologica di sezioni sottili hanno confermato la loro validità in questo tipo di indagine.
Materiali e metodi
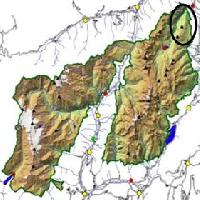
L’area di studio è situata nel Trentino occidentale (Fig. 1), comune di Pellizzano, nei pressi del Parco Naturale Adamello-Brenta, (coordinate Gauss-Boaga / Roma40 16379559; 5127926), limitata a Nord dalla Val di Sole e a Sud dalla Presanella (Cima Presanella 3.500 metri), ad Est dal Gruppo del Brenta e ad Ovest dal Massiccio granitico dell’Adamello. All’intero di una pecceta altimontana, si è scelta un’area di circa 1000 m2 a quota 1740 m s.l.m. su substrato tonalitico coperto da morenico con esposizione Nord nei pressi del Monte Nambino (2678 m s.l.m).
Fig. 1 - Mappa della Provincia Autonoma di Trento e indicazione del sito di studio.
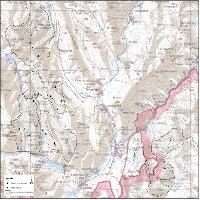
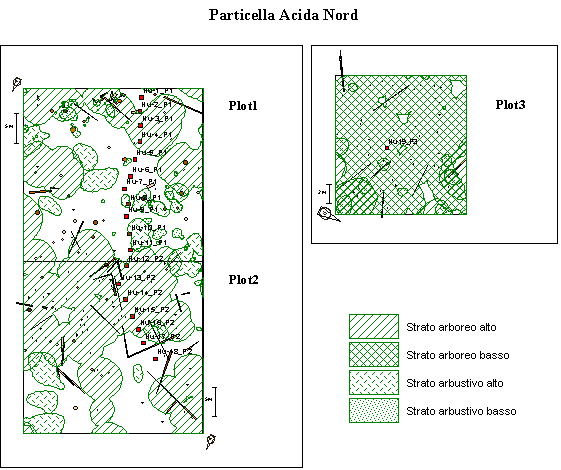
Nell’area prescelta (Fig. 2) si è creato un transetto, disposto spazialmente in modo da attraversare quattro fasi dinamiche del bosco: apertura, fase di rinnovazione, fase intermedia, fase adulta. Lungo il transetto, ogni 3 metri, si sono eseguite le buche per la raccolta del materiale da studiare.
Fig. 2 - Siti di prelievo degli orizzonti umiferi. I profili 5, 13, 18 e 19 sono stati campionati anche per lo studio delle sezioni sottili.
La metodologia di campionamento degli orizzonti per l’indagine morfologica allo stereoscopio è stata descritta da Ponge ([8]) e da Bernier & Ponge ([1]) e adattata allo studio in oggetto.
In campo, con l’ausilio di un coltello, si è creata un’apertura larga circa 40 cm con una profondità tale da riuscire ad osservare completamente gli orizzonti OH ed A, ovvero raggiungendo B o la roccia madre. Si sono quindi prelevati strati di suolo omogenei e possibilmente non manipolati, aventi superficie di 25 cm2 (cm 5 x 5) e profondità uguale allo spessore dell’orizzonte Lo strato di suolo così raccolto è stato posto in un contenitore di polipropilene, riempito con alcool al 95% e chiuso ermeticamente. In laboratorio si è posto ciascun campione in una scatola di Petri del diametro di 15 cm; sopra il suolo si è posto un disco trasparente, dello stesso diametro della Petri, al quale, precedentemente sono stati praticati 157 fori distanti 1 cm l’un l’altro. Dopo aver ricoperto il disco di alcool, per aumentare la nitidezza della visuale, si è eseguita l’osservazione di ciascun campione per mezzo di uno stereoscopio con ingrandimenti fino a 40X. L’annotazione della tipologia e della quantità di escrementi della pedofauna osservati sotto la griglia di punti, eseguita con l’ausilio di una chiave di riconoscimento degli escrementi della pedofauna ([3]), ha permesso di individuare, per ciascun orizzonte, gli animali del suolo in esso attivi.
Quattro dei diciannove profili di humus sono stati inoltre raccolti per le osservazioni micromorfologiche ([2]). I campioni, prelevati mediante scatole di Kubiena (contenitori rettangolari in alluminio), comprendono la parte inferiore degli orizzonti organici (nella maggior parte dei casi OH) ed i primi centimetri dell’orizzonte sottostante (A, E o B). I medesimi sono stati portati in laboratorio dove dopo essere stati essiccati sono stati impregnati sotto vuoto con resina liquida poliestere. I campioni sono stati sezionati e levigati con una troncatrice a mola attraverso diversi cicli di rettifica. Dopo aver ottenuto una superficie perfettamente piana la sezione viene incollata con una resina epossidica su un vetrino portaoggetti delle dimensioni di 90 x 60 mm (sezione 1/4 mammouth). Il campione viene smerigliato sottoponendolo ad un altro ciclo di rettifica fino a raggiungere uno spessore di 30 µm. Attraverso il microscopio polarizzatore si sono osservati caratteri del suolo. In particolare ci si è concentrati soprattutto sulla struttura (dimensione degli aggregati e paleoaggregati) e sul tipo di coproliti presenti.
Risultati
L’osservazione micromorfologica semiquantitativa allo stereoscopio, attraverso l’ausilio di una chiave di riconoscimento degli escrementi della pedofauna ([3]), e l’indagine micromorfologica delle sezioni sottili hanno permesso di rilevare la presenza delle principali categorie di animali del suolo. Si è pertanto dedotto che nella zona indagata la pedofauna è rappresentata principalmente da:
- Enchitreidi: classe Oligochaeta, famiglia Enchytraeidae. Nelle regioni temperate la loro dimensione varia da qualche millimetro fino a 2-3 cm; il loro aspetto è filiforme. Questi anellidi presentano metameria ed appaiono generalmente biancastri e trasparenti, si cibano principalmente di residui vegetali e, in particolare dei tessuti parenchimatici delle lamine fogliari. Generalmente le loro deiezioni, di forma ovoidale con contorni irregolari e di dimensioni inferiori o uguali a qualche decimo di millimetro, si accumulano nell’orizzonte OH e sono costituite dalla mescolanza di residui vegetali ([9], [3]). Nel sito indagato si sono trovati parecchi escrementi definibili come organominerali; ciò è dovuto al fatto che in alcuni casi questi animali si cibano di escrementi già presenti nel suoli. In particolare nel caso in esame si sono cibati di feci organominerali di vermi epigei.
- Vermi epigei: classe Oligochaeta, famiglia Lumbricidae. Hanno dimensioni comprese tra 0.5 e 5 cm, pigmentazione rossa e tollerano bassi valori di pH del suolo. Vivono nella porzione superficiale del terreno, cibandosi principalmente di lettiera; talvolta scendono anche negli orizzonti più profondi dando origine ad escrementi organominerali. Gli escrementi hanno dimensioni variabili da qualche decimo di millimetro a 2 mm ed appaiono cilindrici con punta conica ([9], [3]).
- Vermi anecici: classe Oligochaeta, famiglia Lumbricidae. La loro lunghezza è superiore a 5 cm, la colorazione è rosa bruno e sono molto sensibili ai bassi valori di pH del suolo. Durante la loro attività compiono migrazioni dalla superficie del suolo agli orizzonti minerali, creando gallerie che possono raggiungere profondità notevoli (fino a 1 m). Sono in grado di riciclare la lettiera in modo molto rapido e sono i principali artefici dell’intima unione tra materia minerale e materia organica. Le deiezioni, di un centimetro ed oltre, hanno forma glomerulare ([9], [3]).
- Acari oribatidi: classe Arachnida, sottoclasse Acari, ordine Oribatida. Nelle regioni temperate le loro dimensioni variano da 0.1 a 2 mm. Normalmente si trovano al di sotto della lettiera e si nutrono di materiale vegetale, di ife fungine e di batteri. Il loro apparato boccale è dotato di potenti appendici capaci di triturare il cibo. Tra gli acari del suolo, gli oribatidi assumono una particolare importanza nei processi legati all’umogenesi. Particolare importanza riveste la loro azione nella degradazione degli aghi delle resinose: essi penetrano nell’ago attraverso un minuscolo forellino praticato ad un’estremità, si nutrono dei tessuti parenchimatici e fuoriescono. I loro escrementi presentano colore bruno/nero, forma cilindrica e piccole dimensioni (30-50 µm - [9], [3]).
L’osservazione morfologica allo stereoscopio dei 19 profili ha permesso di giungere alle seguenti considerazioni:
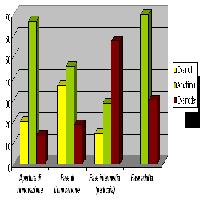
- L’orizzonte OH, nelle fasi di apertura e rinnovazione, è caratterizzato dalla presenza quasi esclusiva di escrementi di verme epigeo olorganici, nelle fasi di bosco maturo e di perticaia invece diventano più abbondanti gli escrementi olorganici di enchitreide (Fig. 3).
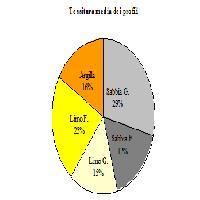
- L’orizzonte A, in fase di apertura è dominato da escrementi di verme epigeo organominerali. Proseguendo lungo il transetto compaiono in questo orizzonte, talvolta in modo quasi esclusivo, gli escrementi organominerali di enchitreidi. La presenza di lombrichi anecici si è desunta nella zona di apertura (profilo 18) e tra alcuni grossi massi, ad una elevata profondità, in un profilo eseguito sotto il bosco maturo (Fig. 4);
- Lo spessore dei profili di humus aumenta gradualmente, anche se non in modo continuo, lungo il transetto, ovvero all’aumentare dell’età del bosco (Fig. 5).
- La fase di crescita che presenta maggiore variabilità di escrementi della pedofauna è il bosco maturo (Fig. 6, Fig. 7, Fig. 8).
Fig. 3 - Variabilità di presenza ed abbondanza degli escrementi della pedofauna nell’orizzonte OH.
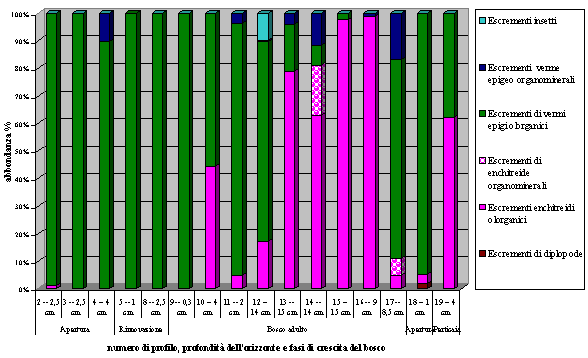
Fig. 4 - Variabilità di presenza ed abbondanza degli escrementi della pedofauna nell’orizzonte A, lungo il transetto.
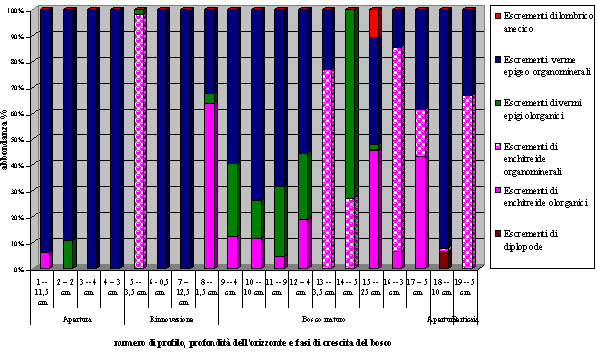
Fig. 5 - Profondità degli orizzonti OH ed A lungo il transetto.
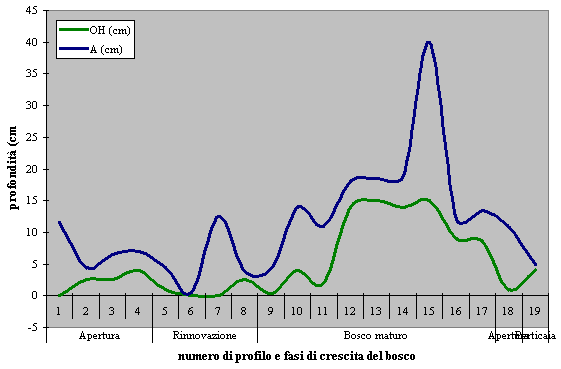
Fig. 6 - Variabilità ed abbondanza degli escrementi della pedofauna nelle 4 fasi dinamiche del bosco.
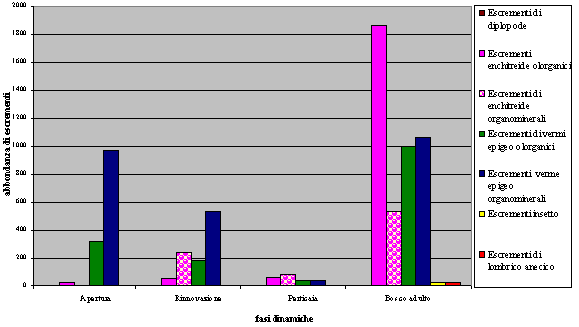
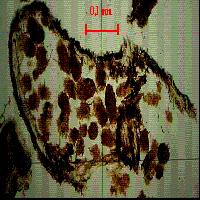
Fig. 7 - Osservazione al microscopio polarizzatore: escrementi di acaro all’interno di una struttura vegetale.

Fig. 8 - Osservazione al microscopio polarizzatore: escremento fossile.

Discussione
Il presente studio ha messo in evidenza la validità dei due approcci utilizzati nell’indagine approfondita della morfologia degli orizzonti umiferi, con particolare riferimento all’abbondanza degli escrementi della pedofauna e alla loro dimensione.
L’osservazione semiquantitativa allo stereoscopio permette, con facilità, buona precisione e discreta velocità, di ottenere interessanti informazioni. L’indagine micromorfologica di sezioni sottili si può ben affiancare al metodo precedente, soprattutto quando le dimensioni degli elementi da osservare sono inferiori a 0.5 mm. A causa degli elevati costi e dei lunghi tempi di preparazione del campione e di esecuzione delle sezioni, questa tecnica dovrebbe infatti essere utilizzata nel caso siano necessari studi di dettaglio da effettuarsi solo su alcuni campioni ben selezionati.
Ringraziamenti
La Dott.ssa Sylvaine Camaret del Laboratoire d’Écologie Alpine, Université de Savoie, Chambery (France) ha fornito un valido supporto scientifico per lo studio della vegetazione. Il presente lavoro è stato svolto nell’ambito del Progetto di Ricerca “DINAMUS. Forme di humus e dinamica del bosco”, diretto dal Centro di Ecologia Alpina del Monte Bondone (TN) e finanziato dal Fondo per i Progetti di Ricerca della Provincia Autonoma di Trento con delibera n. 437 dd. 08/03/2002.
References
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar