Dynamics and management of silver fir stand in Tuscany: a review of long-term research
Forest@ - Journal of Silviculture and Forest Ecology, Volume 5, Pages 122-130 (2008)
doi: https://doi.org/10.3832/efor0517-0050122
Published: May 21, 2008 - Copyright © 2008 SISEF
Research Articles
Abstract
The paper aims to outline the guidelines for the management of silver fir stands in Tuscan Apennines, according to a review of research activities carried out by the Silviculture Institute of Florence, nowadays Department of Forest Environmental Technologies (DISTAF), from 1960 up to today. The study cases are placed in the Vallombrosa Forest Reserve (province of Florence), in the National Park of Foreste Casentinesi Monte Falterona e Campigna (province of Arezzo) and in Amiata Mountain (province of Siena). The main research activities concern different issues linked to the dynamics of silver fir stands: 1) ecology of natural regeneration, 2) stands dynamics in time, 3) sapling growth rates in relation with different canopy cover, 4) impact of wildlife on regeneration storey, 5) forest typology according to vegetation dynamics. As regards management aspects, the decrease of the importance of silver fir pure stands for timber production, as well as the increase of their role from naturalistic, recreation and landscape point of view, encourage close-to-nature sylviculture interventions aimed to promote stand dynamics towards mixed forest. Selective cuttings are proposed in order to progressively reduce the tree canopy cover and to favour the growth of regeneration layer. Nevertheless, wherever silver fir artificial stands play an important role from an historical point of view, the conservation of pure stand by means of clear cutting and artificial regeneration is recommended. Finally, the paper outlines the necessity to face the interactions between forest regeneration and wild ungulate populations, which currently represents one of the most dramatic problems of sustainable management.
Keywords
Abies alba, natural regeneration, forest stand dynamic, silviculture, forest typologies, wild ungulate
Introduzione
L’obiettivo del lavoro è l’individuazione delle linee guida per la gestione sostenibile delle abetine di abete bianco della Toscana, proponendo una sintesi dell’attività di ricerca condotta dalla seconda metà degli anni ’60 ad oggi. Gli studi presero il via nell’Istituto di Selvicoltura dell’Università di Firenze per iniziativa del prof. Ezio Magini, e sono proseguiti fino ai nostri giorni grazie a un gruppo di ricerca del DISTAF della medesima Università. Le indagini sono inquadrabili in tre filoni di ricerca principali, condotti in buona parte in parallelo:
- ecologia della rinnovazione naturale dell’abete bianco, con particolare riferimento all’influenza del microclima luminoso ([24], [23], [27]);
- dinamiche evolutive dei popolamenti, con particolare riferimento alle caratteristiche fisionomiche dei soprassuoli, al ritmo di accrescimento del novellame di abete e all’influenza della pressione esercitata dagli ungulati ([24], [23], [35], [8], [10], [11]);
- aspetti selvicolturali e gestionali, con la messa a punto di una tipologia delle abetine casentinesi su base evolutiva ([9], [4]).
Quanto esposto di seguito è una revisione critica dei risultati ottenuti fino ad oggi, anche alla luce di studi affini condotti in altre zone d’Italia e all’estero.
Sintesi dei risultati
Rinnovazione e accrescimento giovanile
Le prime esperienze di Magini ([24]), concentrate nella foresta di Vallombrosa, miravano allo studio dei problemi della rinnovazione naturale dell’abete bianco, in soprassuoli di composizione specifica variabile. Lo studio, condotto in una quarantina di aree, mise in evidenza che la rinnovazione della conifera era favorita in zone di margine, mentre i tratti di soprassuolo più densi e le ampie radure si rivelavano sfavorevoli. La spiegazione dei fenomeni fu attribuita, in massima parte, al microclima luminoso: furono messe in luce correlazioni positive e molto significative fra irradianza relativa (I.R.) e indice di rinnovazione ([24]). Dove i valori di I.R. scendevano sotto il 2% il novellame di abete tendeva a scomparire, per valori compresi fra il 2 e il 70%, la densità e le dimensioni dei semenzali di abete crescevano con la disponibilità di luce, mentre per valori superiori al 70% il novellame veniva sopraffatto dalla vegetazione nitrofila. Restò aperto il problema dell’assenza di rinnovazione dell’abete in bosco puro, anche in condizioni d’illuminazione favorevoli; successivamente gli studi di Becker & Drapier ([5]), avrebbero in parte spiegato il fenomeno mettendo in luce il ruolo inibitorio svolto dalle sostanze fitotossiche contenute negli aghi dell’abete.
Nel 1988, ricalcando la metodologia del lavoro precedente, i rilievi furono ripetuti in 20 aree, in modo da verificare le tendenze evolutive dei processi segnalati un ventennio prima. Le nuove esperienze confermarono quanto anticipato da Magini sugli effetti della luce nei confronti della rinnovazione dell’abete ([27]). Soprattutto, fu evidenziato lo stato di sofferenza del novellame di abete nei soprassuoli in cui, nel corso del ventennio, il grado di copertura arboreo era aumentato ([23]). Con questo studio si spostò l’attenzione più sui processi successionali nel loro insieme che sulla rinnovazione dell’abete in sé: l’obiettivo prioritario era ormai l’individuazione delle linee guida per la gestione dei soprassuoli in fase di rinaturalizzazione (nei casi in cui si ritenesse opportuna la conservazione dell’abetina pura, si procedeva comunque con la rinnovazione artificiale). Il problema della rinnovazione naturale dell’abete, che aveva ancora significato scientifico e sperimentale, sotto l’aspetto gestionale rivestiva sempre meno importanza.
Nel 2004, a distanza di quasi 40 anni dalle prime osservazioni, i rilievi furono ripetuti in sei delle aree storiche di Magini ([8], [10]). I principali risultati si possono così riassumere:
- In abetine fortemente diradate per cause naturali, la rinnovazione delle latifoglie (fortemente cresciuta nel periodo 1966-1988) nel 2004 ha subito un vistoso regresso: le ragioni sono legate in parte ai danni da daino, in parte alle carenze luminose dovute alla parziale chiusura del piano superiore delle chiome dell’abete.
- La rinnovazione di abete, accompagnata da quella di latifoglie, è risultata quantitativamente apprezzabile solo in poche situazioni caratterizzate da aperture frequenti e di piccole dimensioni (< 100 m2), dovute a cause naturali. Ne risultano mosaici strutturali in cui si alternano tratti a copertura più o meno densa a tratti scoperti (grande ricchezza di margini): a partire dal 1988, in tali situazioni il novellame di abete ha cominciato a manifestare i sintomi del prolungato aduggiamento (cimale mal conformato o, a volte, addirittura biforcato, formazione del nido di cicogna, scarsa vigoria generale).
- In una fustaia transitoria di castagno, in cui 1966 era stato segnalato un vistoso insediamento dell’abete bianco, si è registrato un sensibile regresso delle condizioni vegetative dell’abete, a causa del lungo ombreggiamento subito. Dove la copertura è stata interrotta da attacchi parassitari, vegetano tuttavia gruppi di novellame più promettenti. Gli accrescimenti e lo stato vegetativo indicano una marcata differenza tra gli abeti presenti nel piano inferiore (nido di cicogna, accrescimenti longitudinali degli ultimi 5 anni limitatissimi, frequenza di fusti policormici) e quelli del piano intermedio (portamento piramidale, accrescimenti sostenuti).
I risultati hanno suggerito, a questo punto della ricerca, di studiare i modelli di accrescimento giovanile dell’abete bianco, in modo da valutarne l’influenza sul processo di rinnovazione.
Una prima serie di indagini è stata condotta in aree non interessate dalla presenza degli ungulati selvatici: si tratta di tre aree “storiche” di Vallombrosa, nelle quali è presente un denso e affermato piano di rinnovazione di abete bianco ([10]). Attraverso indagini dendrocronologiche è stato valutato il ritmo di accrescimento longitudinale di piante con altezza compresa fra 0.5 e 5 m, espresso in termini di tempo di passaggio (Tp), che nel caso in questione è il numero di anni necessario per crescere di 0.5 m in altezza. L’indagine ha evidenziato che, in tutte le aree e indipendentemente dall’attuale altezza degli individui, il Tp medio è significativamente superiore (p < 0.01) nel primo mezzo metro di altezza (circa 14 anni), diminuendo progressivamente negli intervalli successivi, in funzione delle caratteristiche del soprassuolo (grado di copertura).
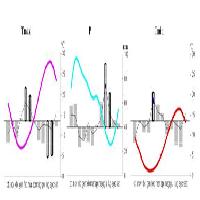
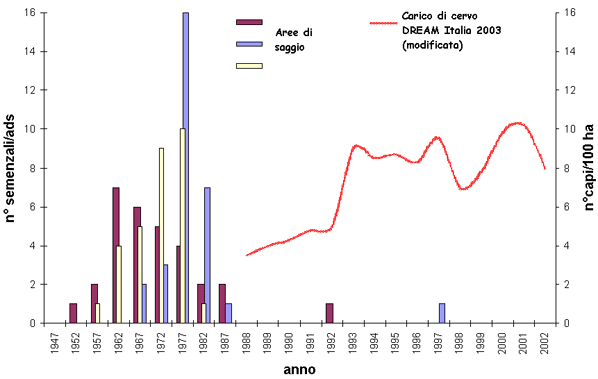
Una seconda serie di indagini è stata successivamente condotta in Casentino, in condizioni di forte pressione della fauna selvatica ([11]). I risultati evidenziano che i danni da ungulati riguardano oltre la metà delle piante del piano di rinnovazione e che la conifera è la specie maggiormente interessata, soprattutto per le piante di altezza inferiore a 3 m. I ritmi di accrescimento longitudinale sono paragonabili a quelli registrati a Vallombrosa, tuttavia è emersa una maggiore lentezza di crescita per le piante alte meno di 1.5 m. Questo fenomeno è attribuibile al fatto che le piante di taglia inferiore, insediatesi più recentemente (in concomitanza con l’inizio dell’incremento del carico degli ungulati, avvenuto all’inizio degli anni ’90 con l’istituzione del Parco), avevano all’epoca un’altezza tale da essere facilmente danneggiate dal morso degli animali: ciò ha ulteriormente rallentato il loro accrescimento. Infatti, confrontando la dinamica di insediamento del novellame di abete bianco con la curva che esprime l’esplosione della densità della popolazione di cervo (Fig. 1), è evidente la quasi totale mancanza di semenzali insediati dopo la fine degli anni ’80.
Fig. 1 - Insediamento del novellame di abete bianco (si sono tenute distinte le aree di studio) e dinamica della popolazione di cervo in Casentino. In ascissa sono indicati gli anni, in ordinata il numero semenzali campionati (a sinistra) e il numero di capi di cervo stimati per 100 ha (a destra, dati [16]).
Dinamiche strutturali dei soprassuoli e tipologie
A seguito dell’attuale tendenza alla gestione di soprassuoli in rinaturalizzazione con criteri riconducibili alla selvicoltura naturalistica ([34]), sono state condotte ricerche inerenti alla valutazione delle dinamiche strutturali nelle abetine. Gli studi riferibili a questo filone sono stati concentrati nel bosco della SS. Trinità presso Santa Fiora, situato sulle pendici del Monte Amiata ([35]), e nelle abetine del Casentino ([8]). Le analisi sono state condotte valutando le relazioni tra la diversità specifica e strutturale dei soprassuoli da una parte, e i caratteri quantitativi e qualitativi del piano di rinnovazione dall’altra. Per approfondire l’analisi della struttura verticale dei popolamenti è stato sperimentato un apposito indice che valuta il livello di stratificazione (indice strutturale, Is).
Il risultato di maggiore spicco, comune alle due zone di studio e attestato da alti valori di correlazione, è che sia la densità sia la diversità specifica del novellame sono positivamente influenzate da diversità specifica e complessità strutturale del soprassuolo.
Per le abetine casentinesi è stato possibile approfondire aspetti relativi alle dinamiche strutturali ([10]). Si è osservato che i soprassuoli si mantengono monoplani fino a 70-80 anni (soglia di età che tende a diminuire con l’abbassarsi della quota), salvo perdurare in tale struttura nelle stazioni più scadenti. A questa fase segue una stratificazione del soprassuolo, legata a una diminuzione del grado di copertura del piano superiore. Il novellame di abete (raro) e di latifoglie (originate da seme, assieme ai polloni originati da ceppaie di piante preesistenti all’impianto e agli individui piantati) va a costituire lo strato inferiore. Si avvia così una trasformazione che porta la struttura spaziale a una minore omogeneità sia orizzontale, in conseguenza delle aperture, sia verticale, come risultato dell’affermazione di uno strato di rinnovazione (Fig. 2). La struttura biplana che ne deriva è una tappa di passaggio in direzione di quella pluristratificata, possibile grazie a un piano intermedio discontinuo e non troppo denso, che permette l’insediamento di un piano inferiore (la probabilità di perdurare in una struttura biplana è tanto maggiore quanto minore è la capacità di affermazione della rinnovazione naturale da seme). In Casentino, le abetine rimangono a lungo in fase di struttura biplana (pochissime abetine sono classificabili come “pluristratificate”), fenomeno legato, oltre che alle caratteristiche stazionali, alla pressione esercitata degli ungulati selvatici.
Fig. 2 - Rappresentazione grafica semplificata delle strutture osservate: A = monoplana, B = biplana, C = pluristratificata.
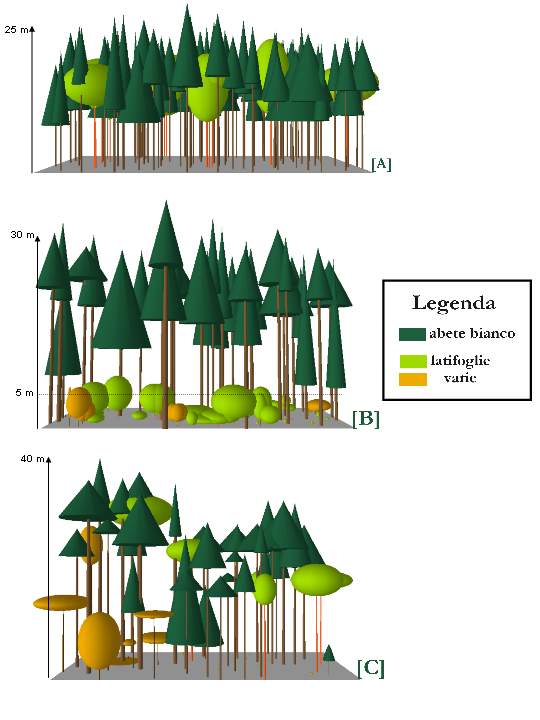
Va aggiunto che in una stessa particella possono coesistere vari livelli di stratificazione in funzione delle caratteristiche stazionali e di fattori di disturbo (biotici o abiotici). Infatti, con l’aumento dell’età del soprassuolo principale, aumenta la possibilità di osservare, all’interno di una stessa particella (se di superficie sufficientemente ampia), diversi tipi strutturali (“mosaico di strutture”).
I risultati delle indagini condotte hanno originato studi tipologici, che prendono spunto dalla tipologia dei boschi della Toscana ([30]). In particolare sono state oggetto di studio le zone situate all’interno delle Riserve Naturali Biogenetiche Statali (Camaldoli, Badia Prataglia e Scodella), appartenenti al Demanio Statale e in gestione all’Ufficio Territoriale per la Biodiversità di Pratovecchio (ex-ASFD), e alcuni territori del Demanio Regionale gestiti dalla Comunità Montana del Casentino (Complesso Foreste Casentinesi).
La classificazione tipologica iniziale, basata su caratteri fisionomici, è stata successivamente modificata in base ai risultati di un’analisi discriminante, che ha così permesso di valutare quali variabili differenziassero, meglio di altre, le fisionomie individuate.
Data l’origine artificiale delle abetine e la sostanziale omogeneità delle caratteristiche geo-pedologiche della zona, si è considerato un unico tipo (abetina di origine artificiale a faggio), all’interno del quale si sono individuati sottotipi. Ne risulta il seguente schema ([4]).
Le abetine altimontane, poste oltre 1300 m s.l.m., in stazioni con pendenza accentuata, attualmente ospitano popolamenti di abete bianco su ex-pascoli, di fertilità medio-bassa. È forte l’incidenza di piante mal conformate, schiantate e sradicate a causa delle avverse condizioni meteoriche (neve, galaverna, vento). I processi evolutivi, influenzati dal breve periodo vegetativo, sono molto lunghi, la copertura dello strato arboreo oscilla dal 65 all’85%, la struttura è tendenzialmente biplana.
Il sottotipo più rappresentato è quello delle abetine montane, dislocate fra 800 e 1300 m s. l. m. Esse si distinguono dalle precedenti per il dinamismo più marcato, dovuto a un periodo vegetativo più lungo e una generale maggiore fertilità stazionale (si tratta di abetine spesso impiantate su ex-coltivi), che determinano un evidente sviluppo dei piani inferiori. Nelle interruzioni della copertura, dovute a tagli intercalari o fitosanitari, sradicamenti e schianti, si osserva l’insediamento di latifoglie (soprattutto faggio), mentre sporadica è la rinnovazione di abete bianco. La struttura di riferimento è biplana per gruppi, anche se alle quote inferiori è possibile imbattersi in strutture multiplane. All’interno del sottotipo si distinguono due varianti: a faggio e a latifoglie mesofile (quest’ultima diffusa a quote inferiori).
Un sottotipo abetine non evolute, trasversale ai precedenti, è stato distinto in quanto degno di una gestione a parte. Generalmente si tratta di abetine di età inferiore a 70 anni, con un grado di copertura molto elevato (85-95%), tale da ostacolare evoluzioni, fenomeno particolarmente evidente dove i tagli intercalari non sono stati effettuati o hanno avuto bassa intensità. Ne risulta una struttura di riferimento monoplana. Nelle abetine più vecchie l’insediamento dei piani inferiori è ostacolato, secondo i casi, dalla scarsa fertilità stazionale o dalla pressione della fauna ungulata.
Indirizzi gestionali
Le abetine pure toscane sono il risultato di modificazioni fisionomiche, floristiche e provvigionali degli originari consorzi misti del cingolo Fagus-Abies, in cui l’abete era mescolato, per singole piante o a gruppi, al faggio e a latifoglie mesofile ([36]). Negli ultimi decenni, nella maggior parte di questi soprassuoli, le pratiche selvicolturali si sono progressivamente ridotte in termini di frequenza, intensità e superficie di taglio: oggi le utilizzazioni sono per lo più limitate al recupero di piante schiantate o a diradamenti dal basso molto moderati. Il trattamento tradizionale - il taglio raso con rinnovazione artificiale posticipata (turno di 90-100 anni) in superfici di 1-3 ha - non è più praticato. In particolare, sono cambiate le funzioni attribuite a questi boschi: hanno acquistato maggiore importanza gli aspetti turistico-ricreativi, paesaggistici, protettivi e di conservazione, a scapito di quelli produttivi. Visto che buona parte di tali soprassuoli ricade nell’ambito di aree protette (Riserve Biogenetiche, Parchi ecc.), la gestione delle abetine mira ad incoraggiare l’evoluzione verso il bosco misto di abete e latifoglie - cui si accorda una sostanziale maggiore stabilità - in linea con quanto accade nel resto dell’Europa ([44], [41]).
In questo senso, le osservazioni nelle aree permanenti di Vallombrosa nel periodo 1966-2005 suggeriscono alcune riflessioni. L’ingresso di latifoglie sotto la copertura dell’abete bianco, fenomeno segnalato già a partire dalla metà degli anni ’60 e progredito fino al 1988, appare, secondo i casi, stabile o in regresso. Anche buona parte del novellame di abete manifesta oggi sintomi di sofferenza indotti dal prolungato ombreggiamento. Appare invece stabile, nel tempo, la situazione in condizioni estreme di copertura arborea. Infatti, sia nelle ampie radure invase dalla flora nitrofila, sia sotto la densa copertura all’interno delle abetine, a distanza di quarant’anni non si ha traccia di rinnovazione di specie arboree. Fra i numerosi fattori chiamati in causa, due assumono maggior rilievo: la disponibilità di luce e la pressione degli ungulati selvatici.
Per quanto concerne la radiazione luminosa, è nota l’elevata efficienza dei semenzali di abete bianco a sfruttare bassi livelli di tale fattore ecologico ([23], [18], [33], [19], [22]). Questo adattamento permette all’abete di evitare la concorrenza della vegetazione erbaceo-arbustiva ([24], [37]), come di quella arborea ([30], [32]). Ciò si traduce in una tendenza ad affermarsi nelle zone dove sussistono condizioni di margine: l’abete bianco si comporta cioè come una specie opportunista ([40]) che combina la tolleranza dell’ombra con la capacità di riprendere l’accrescimento appena si verificano adeguati livelli radiativi.
Quindi, volendo favorire la rinnovazione dell’abete, interruzioni localizzate della copertura potrebbero essere efficaci nel promuovere il processo. Nello specifico, potrebbe trattarsi di tagli marginali, tagli a scelta ([41]) o tagli successivi su piccole superfici ([7]). Tuttavia, visto il particolare ambito in cui si realizzano le condizioni di nicchia per l’insediamento del novellame, è opportuno iniziare gli interventi a partire da nuclei di semenzali con buone prospettive di affermazione. Questi trattamenti, essendo condizionati dalle dinamiche del piano di rinnovazione, prescindono da un turno prestabilito e determinano la formazione di soprassuoli irregolari. Dal punto di vista operativo è importante stabilire cosa intendere con il termine “semenzali affermati”. Studi condotti in Polonia ([43]) evidenziano che l’età in cui i semenzali di abete iniziano a crescere rapidamente in altezza è 15 anni: a questa soglia, in base ai risultati ottenuti, corrisponde un’altezza di circa 50 cm che può costituire, in assenza di disturbi da parte di ungulati, la dimensione minima di riferimento.
Il taglio a buche, recentemente riconsiderato in Italia ([29], [15]) e all’estero ([25]), può favorire la rinnovazione naturale dell’abete, anche se non sempre il successo è garantito: tagli eseguiti nel 1982 nelle abetine di Camaldoli hanno messo in evidenza, a sedici anni di distanza, dinamiche ancora in una fase arbustiva pre-forestale ([29]). Anche in relazione a quanto suggerito da esperienze recenti ([2]), le dimensioni delle buche dovrebbero essere comprese tra 500 e 1000 m2, sebbene nelle condizioni stazionali più favorevoli l’estensione possa essere anche leggermente superiore (fino a 1500 m2).
Nella maggior parte dei soprassuoli, tuttavia, gli interventi selvicolturali saranno indirizzati ad assecondare le dinamiche evolutive in direzione del bosco misto di latifoglie e abete. I trattamenti suggeriti si basano su interventi localizzati, tempestivi e d’intensità pari alla capacità di affermazione dei piani inferiori: il dinamismo strutturale dei soprassuoli andrebbe favorito con “tagli di liberazione”, mirati a ridurre progressivamente la copertura del soprassuolo adulto allo scopo di favorire l’accrescimento del novellame insediato in seguito al processo successionale (Fig. 3).
Fig. 3 - Simulazione di taglio di liberazione in favore del piano di rinnovazione sotto l’abetina.
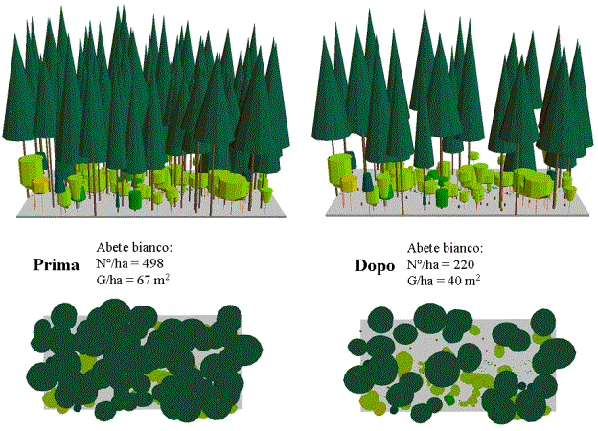
In ogni caso, prima di applicare qualsiasi intervento selvicolturale occorre affrontare il problema della pressione degli ungulati selvatici ([39], [31]).
Vari fenomeni (ripopolamento di cervo, daino e capriolo, spesso associato alla riduzione della pressione venatoria) hanno portato, negli ultimi anni, all’incremento demografico delle popolazioni di ungulati, oggi fattore cruciale per lo sviluppo e l’affermazione della rinnovazione naturale e artificiale di molte specie forestali. Come è stato anticipato, la brusca interruzione del processo di rinnovazione dell’abete bianco osservata in Casentino a partire dalla fine degli anni ’80 va messa in relazione soprattutto all’attività degli ungulati. L’abete bianco è la specie che più di altre soffre gli effetti della brucatura ([17], [28]), per quanto anche molte latifoglie, fra cui soprattutto l’acero di monte, subiscano gli effetti del morso.
La situazione Toscana in questo senso è critica ma non singolare, visto che il problema dei danni a carico del novellame di abete bianco è grave anche in altre aree italiane ed estere ([42], [13], [6], [20], [21], [38]).
La selettività dei danni di origine alimentare mette a repentaglio, nel lungo periodo, la possibilità di affermazione delle specie più appetite: possono risultarne modifiche nella struttura e nel dinamismo evolutivo degli ecosistemi forestali ([3], [31]).
Alla luce di quanto detto, si schematizzano le linee-guida per la gestione delle abetine toscane:
- assecondare le dinamiche evolutive dei popolamenti in direzione del bosco misto a struttura irregolare: il risultato atteso è un incremento di complessità e stabilità dei sistemi;
- favorire il mantenimento di una copertura continua al suolo, avvalendosi di rinnovazione naturale eventualmente intergrata con piantagioni (in particolar modo nelle fasce di protezione e di maggiore pendenza);
- assicurare la stabilità meccanica dei soprassuoli puri attraverso tagli intercalari;
- individuare, valorizzare e preservare le abetine pure di particolare interesse storico, culturale e paesaggistico (ad es., abetine nei pressi del Monastero di Camaldoli e Vallombrosa). A tal fine, non si può prescindere dal trattamento classico che prevede il taglio a raso e la rinnovazione artificiale posticipata ([23]), intervento previsto anche nel Piano di Gestione di Vallombrosa ([14]).
- controllare la fauna selvatica, in particolare gli ungulati.
Considerazioni conclusive
Il progressivo ingresso di latifoglie nelle abetine toscane esprime la tendenza successionale in direzione del bosco misto: le modificazioni fisionomiche che ne derivano variano in funzione delle caratteristiche della stazione e dell’intensità e frequenza dei fattori di disturbo. In un quadro così complesso, le tipologie possono essere di aiuto ai fini gestionali. In particolare, l’individuazione delle dinamiche strutturali appare decisiva ai fini di una corretta applicazione degli interventi selvicolturali. Come regola generale sarebbe opportuno assecondare le dinamiche spontanee dei popolamenti, in considerazione sia dei costi necessari a mantenere le abetine pure sia della maggiore stabilità garantita da popolamenti misti. La diversità tuttavia, ad ampia scala può essere interpretata nel senso della conservazione di tessere rare: in questo senso può essere opportuna la conservazione delle abetine pure storiche ([1]).
La soluzione del problema della pressione della fauna ungulata, come ormai sostenuto da molti studiosi in Italia e all’estero ([26], [39], [12]), è una gestione integrata, che a misure faunistico-venatorie affianchi misure di gestione forestale e ambientale.
References
Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar