Effect of stand structure on models for volume and aboveground biomass assessment (Castelfusano pinewood, Roma)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 6, Pages 75-84 (2009)
doi: https://doi.org/10.3832/efor0569-006
Published: Mar 25, 2009 - Copyright © 2009 SISEF
Research Articles
Abstract
The main purpose of this research was to analyse the effects of stand structure on biomass allocation and on the accurancy of estimation models for volume and aboveground biomass of Italian stone pine (Pinus pinea L.). Although the species is widely distributed on Mediterranean coasts, few studies on forest biomass estimation have focused on pinewoods. The research was carried out in the Castelfusano’s pinewood (Rome) and concerned the two most common structural types: (a) 50 years-old pinewood originated by broadcast seeding; and (b) 62 years-old pinewood originated by partial seeding alternating worked strips to firm strips. Some 83 sample trees were selected for stem volume estimation and a subset of 32 trees used to quantify the total epigeous biomass, the wooden biomass compartment, including stem and big branches (diameter > 3 cm) and the photosynthetic biomass, including thin branches (diameter < 3 cm) and needles. Collected data were used to elaborate allometric relations for stem volume, total biomass and specific relations for both compartments, based on one (d2) or two (d2h) indipendent variables, for both structural types. Furthermore, pinewood specific biomass expansion factors (BEF) - indexes used to estimate carbon stocks starting from stem biomass data - were obtained. The achieved estimation models were subjected to both parallelism and coincidence tests, showing significant effects of stand structure on the accurancy of the allometric relations. The effects of stand structure and reliability of tree height curves on the accurancy of estimation models for volume and aboveground biomass and on biomass allocation in different compartments are analysed and discussed.
Keywords
Pinus pinea L., Structural types, Allometric relations, Biomass allocation, Biomass expansion factors
Introduzione
L’importanza degli ecosistemi forestali nel mitigare gli effetti dei cambiamenti globali mediante la fissazione e lo stoccaggio di grandi quantità di carbonio è ampiamente riconosciuta ([14], [6]). Si stima che le foreste, pur ricoprendo circa il 30% delle terre emerse, contengano più dell’80% della biomassa terrestre ([14], [15]).
Con la ratifica del protocollo di Kyoto il nostro paese si è impegnato in una notevole riduzione delle emissioni dei gas a effetto serra ([10], [9]). Si tratta di una sfida rilevante che, oltre a riflessioni di politica forestale, necessita di un’attendibile valutazione e quantificazione del carbonio stoccato dalle foreste. In ragione di ciò la stima della biomassa - variabile il cui valore è direttamente proporzionale alla quantità di carbonio contenuta all’interno delle foreste - rappresenta una priorità per il mondo scientifico.
I metodi di stima in linea di principio sono due: diretti e indiretti. I primi presupporrebbero l’abbattimento di tutti gli alberi di un popolamento o, per lo meno, di aree rappresentative dello stesso per poi procedere a misure dirette ([21]). Si tratta di un approccio non solo distruttivo ma anche estremamente dispendioso e quindi, di fatto, inapplicabile. Di norma si procede quasi sempre attraverso metodi indiretti basati sull’elaborazione di relazioni allometriche fra la biomassa e alcuni parametri dendrometrici facilmente misurabili come il diametro, l’altezza e, in alcuni casi, il diametro della chioma ([21], [30], [4]). In genere, con l’abbattimento di un numero contenuto di alberi modello si riesce a disporre di modelli di stima piuttosto accurati.
Tra i fattori in grado di condizionare il livello di accuratezza di questi modelli rientrano tutti quelli che interagiscono con l’accrescimento dei popolamenti forestali. Fra questi, oltre ai fattori di disturbo di natura diretta e indiretta ([6]), giocano un ruolo di primo piano le caratteristiche strutturali del soprassuolo che, esprimendo la disposizione spazio-temporale degli alberi ([24]), condizionano i processi di competizione interindividuale. Nel caso di popolamenti artificiali le combinazioni fra le possibili densità, modalità e sesti di impianto rivestono un ruolo importante per l’accrescimento dei soprassuoli e la ripartizione della sostanza organica nei vari comparti. Ciò vale soprattutto per il pino domestico (Pinus pinea L.), specie ampiamente diffusa lungo i tratti costieri della nostra penisola e la cui architettura è sensibilmente condizionata dalla struttura del popolamento, con forme variabili dalla caratteristica chioma ad ombrello, nel caso di popolamenti radi, alla forma a candelabro, nelle condizioni di maggiore densità ([8]).
A seguito del vasto incendio che ha interessato la pineta di pino domestico di Castelfusano (Roma) nel luglio 2000, il Centro di Ricerca per la Selvicoltura del C.R.A. ha intrapreso un ampio programma di studi finalizzato a individuare forme di trattamento in grado di favorire una gestione razionale sotto il profilo bio-ecologico di questo importante complesso forestale ([1], [2], [3], [13]). In questo contesto, tenuto conto che il pino domestico è specie diffusa nel bacino del Mediterraneo ma per la quale esistono solo studi sporadici sulla biomassa ([7], [25]), sono state avviate indagini per elaborare relazioni allometriche del volume e della biomassa epigea e valutare l’influenza della struttura arborea sia sui modelli di previsione sia sull’allocazione della biomassa nei vari comparti. Lo studio ha interessato i due tipi strutturali più diffusi all’interno della pineta e ha posto a confronto modelli di previsione basati su una (diametro) e due (diametro e altezza) variabili indipendenti. Nel contempo, la disponibilità di stime accurate della biomassa epigea ha anche consentito di calcolare i valori del biomass expansion factors (BEF), indice in grado di fornire, per una data specie e su larga scala, stime speditive dello stock di carbonio a partire dai dati disponibili a livello di inventari forestali ([20], [26], [16], [5], [23]).
Materiali e Metodi
L’area di studio
Lo studio è stato condotto nella pineta di Castelfusano, gestita dal Comune di Roma. La pineta, dal 1996 inclusa nella Riserva Naturale Statale del “Litorale Romano”, si estende su di un’area di circa 1100 ettari, a ridosso della fascia costiera fra Ostia e la Tenuta Presidenziale di Castelporziano. Il substrato geologico è rappresentato da formazioni dunali derivate dal trasporto fluviale di materiale solido da parte del Tevere che dà origine a suoli a tessitura grossolana, non calcarei in superficie, riferibili agli Orhtieutric Arenosol (Carta dei suoli del Comune di Roma).
I rilievi sono stati condotti nei due tipi strutturali più diffusi, che si differenziano sia per l’età che per la diversa tecnica di impianto utilizzata:
- pineta di 62 anni originata da semina su fasce lavorate alternate a fasce salde (Tipo B).
- pineta di 50 anni originata da semina andante ad integrazione dei nuclei di leccio (Quercus ilex L.) e macchia mediterranea preesistenti (Tipo C);
I due tipi presentano differenze sotto il profilo strutturale, conseguenti alla differente modalità di impianto. L’assenza di un organico trattamento selvicolturale ha contributo a mantenere e incrementare nel tempo le diversità e le anomalie strutturali. Nel Tipo B le chiome del pino sono fortemente asimmetriche, compresse e con ridotta porzione verde per l’elevata concorrenza laterale; nel Tipo C le modalità di semina hanno portato ad una aggregazione delle piante di pino per ciuffi o per gruppi compatti, limitando lo spazio individuale a disposizione e determinando chiome scadenti. Le principali caratteristiche dei due tipi strutturali sotto il profilo dendrometrico sono riassunte nella Tab. 1, mentre le distribuzioni delle piante in classi diametriche sono riportate in Fig. 1. In entrambi i casi i dati derivano da rilievi condotti in aree di ricerca permanenti, di forma rettangolare e superficie pari a un ettaro, prima degli interventi sperimentali di diradamento (autunno-inverno 2002-2003). Per maggiori dettagli sulle caratteristiche dei popolamenti studiati si rimanda a Amorini et al. ([1], [2]).
Tab. 1 - Principali indicatori dendrometrici relativi a ciascun tipo strutturale.
| Tipo strutturale |
Età | Densità (n ha-1) |
Area basimetrica (m2 ha-1) |
Diametro medio (cm) |
Altezza media (m) |
|---|---|---|---|---|---|
| B | 62 | 338 | 38.83 | 38.2 | 18.0 |
| C | 50 | 370 | 33.08 | 33.7 | 15.2 |
Fig. 1 - Distribuzione di frequenza del pino domestico per i tipi strutturali B e C.
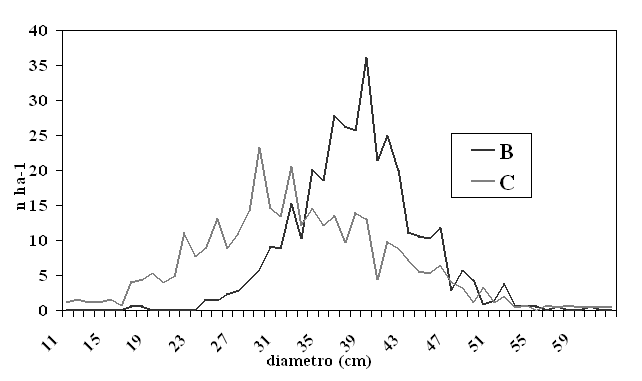
Le differenze osservate, tenuto conto dell’omogeneità dei suoli e della prossimità delle due aree di ricerca, sono da attribuire alla differenza di età e alla diversa modalità di impianto.
Il campionamento
In concomitanza con gli interventi sperimentali di diradamento sono stati abbattuti 83 alberi modello (Tipo B: 44 piante; Tipo C: 39 piante) distribuiti in maniera proporzionale e ponderata per ogni classe diametrica.
Dopo l’abbattimento di ogni albero modello è stato misurato il diametro a 1.30 metri, il diametro ogni metro a partire dalla sezione a 0.50 metri, l’altezza totale e l’altezza di inserzione della chioma. I dati raccolti hanno consentito di calcolare il volume cormometrico comprensivo di cimale utilizzando la formula di Heyer di cubatura per sezioni e di elaborare curve ipsometriche distinte per ciascun tipo strutturale.
La misura della biomassa è stata invece eseguita su un sottocampione di 32 alberi (15 piante del Tipo B e 17 del Tipo C) considerando, oltre alla biomassa totale, due componenti principali: la biomassa legnosa, composta da fusto e rami grossi (diametro > 3 cm), e la biomassa fotosintetizzante, composta da rami fini (diametro < 3 cm) e aghi. Sia problemi logistici che di autorizzazioni (la pineta è inserita nella Riserva del Litorale Romano) non hanno consentito di misurare la biomassa ipogea. Oltre al volume del fusto è stato determinato, direttamente in campo, il peso fresco totale dei rami grossi e dei rami fini. Sono stati poi prelevati campioni legnosi di fusto (in prossimità della base, a metà e in prossimità dell’inserzione chioma), rami grossi e rami fini per la determinazione della densità basale (ρ) del fusto e del rapporto peso fresco/peso secco (θ) per i rami grossi e i rami fini. La densità basale di ciascun campione è stata calcolata misurando con il metodo dell’immersione il volume fresco dei campioni raccolti e, successivamente, determinando il loro peso secco ponendoli in stufa ventilata a 80 ± 2 °C fino al raggiungimento del peso costante. Le variabili densità basale e rapporto peso fresco/ peso secco state utilizzate per la stima della biomassa totale e delle singole componenti. In particolare, per il fusto è stata applicata la formula Psf = Vff · ρ (Psf = peso secco del fusto in Kg, ρ = densità basale in Kg·m-3 e Vff = volume fresco del fusto in m3), mentre per i rami grossi e i rami fini la formula Psr = Pf · θ (Psr e Pf rispettivamente peso secco e peso fresco dei rami grossi/fini in Kg e θ = rapporto tra peso secco e fresco per rami grossi/fini). Dalla somma dei pesi secchi dei vari comparti è stata ottenuta la biomassa totale di ciascuna pianta.
I modelli di previsione
Per l’elaborazione dei modelli di previsione si è proceduto sia con il metodo della stepwise regression sia costruendo una matrice di correlazione tra le variabili dipendenti volume e peso secco (totale e per componenti) e le variabili indipendenti diametro (d), altezza totale (h), d², h², dh, d²h, dh² e d²h². Si è così potuto discriminare le variabili indipendenti maggiormente correlate con le variabili dipendenti analizzate e, di conseguenza, scegliere quelle più idonee alla costruzione di modelli sia a una che a due entrate.
Per verificare gli effetti delle differenze strutturali sui modelli di previsione del volume e della biomassa è stata eseguita sia l’analisi grafica, mettendo a confronto l’andamento delle curve relative al tipo strutturale B con quelle relative al tipo strutturale C, sia una dettagliata analisi statistica attraverso il test F e i test di coincidenza e parallelismo ([11], [12], [28]). Visti i risultati delle matrici di correlazione, i test sono stati eseguiti utilizzando d2 e d2h come covariate, il tipo strutturale come variabile indipendente e i valori di volume e peso secco come variabili dipendenti. I valori di densità basale (ρ), relativi a ciascun tipo strutturale, sono stati sottoposti al test t di Student per escludere una eventuale influenza di questa variabile sui risultati ottenuti. Per l’elaborazione dei modelli e i relativi test statistici è stato utilizzato il software Systat (Systat Software Inc., San Jose, California, USA).
Biomass expansion factor (BEF)
Avendo a disposizione i dati relativi alle caratteristiche dendrometriche dei popolamenti e modelli accurati di stima della biomassa è stato possibile calcolare i valori del BEF per ciascun tipo strutturale. Come è noto il BEF è il risultato di un rapporto fra due variabili, la variabile “target” oggetto di studio al numeratore e quella invece da “espandere” (variabile nota) al denominatore. In letteratura sono stati proposti diversi tipi di BEF in funzione della variabile target e della variabile nota ([20], [26], [16], [22]). In questo studio, secondo la linea recentemente più adottata ([18], [17]), si è scelto di calcolare il BEF come rapporto tra la biomassa totale e la biomassa dei fusti ad ettaro (BEF = biomassa totale / biomassa fusti).
Risultati
Nella Tab. 2 sono riportati il numero, le caratteristiche dendrometriche e i valori di volume del fusto e di biomassa epigea degli alberi modello campionati distinti per tipo strutturale. Il set di dati relativo agli alberi modello ha consentito non solo di elaborare accurati modelli di previsione del volume del fusto e della biomassa ma anche di fare alcune considerazioni sull’allocazione della biomassa tra i vari comparti.
Tab. 2 - Principali caratteristiche degli alberi modello di pino domestico utilizzati per la stima del volume e della biomassa, distinti per tipo strutturale.
| Tipo strutturale |
N. oss. |
Diametro (cm) | Altezza (m) | Volume (m3) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Med | Min | Max | Med | Min | Max | Med | ||
| B | 44 | 24.0 | 49.0 | 35.9 | 16.3 | 19.3 | 18.0 | 0.423 | 1.566 | 0.959 |
| C | 39 | 15.5 | 48.0 | 29.9 | 13.0 | 17.8 | 14.9 | 0.122 | 1.380 | 0.531 |
| Tipo strutturale |
N. oss |
Diametro (cm) | Altezza (m) | Biomassa totale (kg) | ||||||
| Min | Max | Med | Min | Max | Med | Min | Max | Med | ||
| B | 15 | 27.3 | 49.0 | 40.4 | 16.3 | 18.9 | 17.9 | 266.1 | 1714.1 | 844.1 |
| C | 17 | 15.5 | 45.8 | 31.0 | 14.1 | 16.7 | 14.9 | 66.0 | 910.7 | 423.6 |
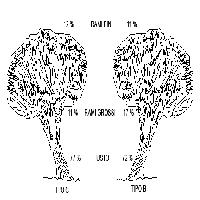
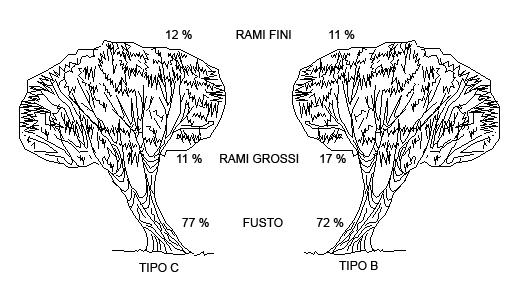
Ponendo a confronto la percentuale di biomassa allocata nel fusto, nei rami grossi e nei rami fini e aghi nei due tipi strutturali emergono alcune differenze (Fig. 2). Se la percentuale allocata nella parte fotosintetizzante è praticamente identica, maggiori scarti si osservano a livello di fusto e rami grossi, con valori in quest’ultimo caso significativamente differenti (test t = -2.965; p = 0.0058). Il test t di Student sui valori di densità basale non ha fatto emergere differenze significative tra i due tipi strutturali (Tipo B = 0.53 ± 0.1; Tipo C = 0.49 ± 0.1).
Fig. 2 - Allocazione della biomassa epigea del pino domestico nei comparti fusto, rami grossi e rami fini.
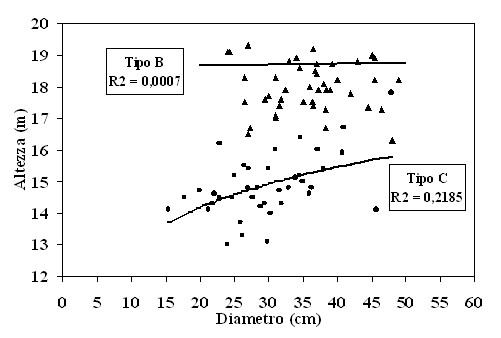
I valori di altezza, misurati a terra sul campione di alberi modello, hanno evidenziato differenze ipso-diametriche tra i due tipi strutturali (Fig. 3). Nel Tipo C, la minore età e un livello di competizione meno omogeneo hanno condotto ad una più marcata differenziazione delle altezze e, di conseguenza, ad una maggiore pendenza della curva. Il valore del coefficiente di determinazione R2, benché non raggiunga valori elevati (R2 = 0.22), è risultato comunque significativo al test di p di Pearson. Al contrario, nel Tipo B la relazione ipso-diametrica è aleatoria come dimostra il forte appiattimento della curva.
Fig. 3 - Curve ipsometriche per i tipi strutturali B [triangoli: y = 0.1089 · Ln(x)+ 18.384] e C [cerchi: y = 1.8399 · Ln(x) + 8.6621].
Dall’analisi della matrice di correlazione emerge uno stretto rapporto tra gran parte delle variabili indipendenti testate e i valori di volume e biomassa epigea (Tab. 3). Fanno eccezione h² e dh² che a causa dei bassi valori di r non vengono riportate in tabella. Tale analisi, ancor più dell’approccio stepwise, consente in maniera efficace e sintetica di apprezzare, non solo il legame tra la singola variabile indipendente e la singola variabile dipendente, ma anche di fornire un quadro di insieme sulle capacità di ciascuna variabile indipendente di essere un buon predittore.
Tab. 3 - Matrice di correlazione fra variabili predittrici e biomassa, nelle sue varie componenti, e volume del fusto. Con (*) sono evidenziate le variabili con la correlazione più elevata. Il livello di significatività per tutte le correlazioni è di p < 0.001.
| Variabile indipendente |
Biomassa legnosa |
Biomassa fotosintetizzante |
Biomassa totale | Volume | |
|---|---|---|---|---|---|
| 1 entrata | d | 0.923 | 0.844 | 0.919 | 0.944 |
| h | 0.668 | 0.479 | 0.646 | 0.644 | |
| d2 | 0.939* | 0.885* | 0.939* | 0.945* | |
| 2 entrate | dh | 0.936 | 0.820 | 0.927 | 0.977 |
| d2h | 0.957* | 0.880* | 0.954* | 0.979 | |
| d2h2 | 0.948 | 0.853 | 0.942 | 0.980* | |
Entrando più nello specifico, nel caso dell’utilizzo di una sola variabile indipendente, quella maggiormente correlata è il d²; tra le possibili combinazioni di due variabili indipendenti, d²h è invece quella con R2 più elevato. A fronte di ciò l’elaborazione dei possibili modelli di previsione del volume e della biomassa, basati su una variabile indipendente (una entrata) e su due variabili indipendenti (due entrate), ha preso in considerazione rispettivamente d2 e d2h.
Gli effetti della struttura del popolamento sull’accuratezza dei modelli di previsione è messa in evidenza dai risultati del test F (Tab. 4). La scelta della covariata, ad una o due entrate, è determinante ai fini della significatività della differenza fra i due tipi di pineta. Selezionando come variabile indipendente il d2, si ottengono differenze significative per tutti i comparti, con le ipotesi di coincidenza e parallelismo sempre rifiutate. Utilizzando come variabile indipendente d2h non emergono invece differenze significative fra il Tipo B e il Tipo C per la biomassa totale e per quella legnosa (ipotesi di coincidenza accettata), mentre queste permangono per la biomassa fotosintetizzante ed in parte per il volume (ipotesi di coincidenza rifiutata ma ipotesi di parallelismo accettata).
Tab. 4 - Verifica delle ipotesi di coincidenza e parallelismo per discriminare gli effetti della struttura sui modelli lineari predittivi prescelti.
| Covariata | Variabile dipendente |
N. campione |
F | Ipotesi | |
|---|---|---|---|---|---|
| Coincidenza | Parallelismo | ||||
| d2 | Volume | 83 | 53.75 | Rifiutata | Rifiutata |
| d2 | Biomassa totale | 32 | 4.27 | Rifiutata | Rifiutata |
| d2 | Biomassa legnosa | 32 | 3.49 | Rifiutata | Rifiutata |
| d2 | Biomassa fotosintetizzante | 32 | 8.46 | Rifiutata | Rifiutata |
| d2h | Volume | 83 | 4.65 | Rifiutata | Accettata |
| d2h | Biomassa totale | 32 | 2.27 | Accettata | Accettata |
| d2h | Biomassa legnosa | 32 | 1.14 | Accettata | Accettata |
| d2h | Biomassa fotosintetizzante | 32 | 7.41 | Rifiutata | Rifiutata |
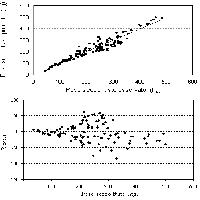
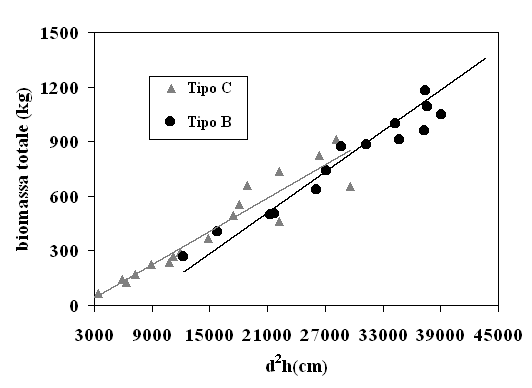
Quanto ora esposto trova conferma anche nell’analisi grafica delle trendline del volume e della biomassa in funzione delle variabili indipendenti selezionate. A titolo di esempio si riportano gli andamenti della biomassa totale in funzione di d2 (Fig. 4) e di d2h (Fig. 5). Si nota che, con d2, i modelli relativi ai due tipi strutturali tendono a divergere in maniera più netta, al contrario di quanto si osserva utilizzando d2h.
Fig. 4 - Biomassa totale del pino domestico in funzione della variabile d2 e del tipo strutturale.
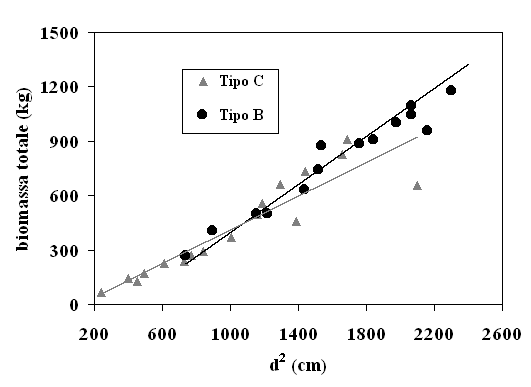
Fig. 5 - Biomassa totale del pino domestico in funzione della variabile d2 h e del tipo strutturale.
Ciò premesso sono stati elaborati una serie di modelli di previsione del volume del fusto e della biomassa epigea mediante regressioni allometriche ad una e due entrate (d2 e d2h). Le funzioni predittrici sono state tenute distinte per i due tipi strutturali. Come tipo di funzione è stata scelta quella lineare y = a + bx che, tenendo costante la forma dell’equazione, garantisce l’additività tra le componenti rispetto al totale. In Tab. 5 vengono riportati i coefficienti a e b della funzione predittrice, i coefficienti di determinazione corretti (adjusted R 2) e l’errore standard delle stime (ESS).
Tab. 5 - Coefficienti e indici statistici dei modelli di previsione a una (d2) e a due entrate (d2h) del volume del fusto, della biomassa totale e delle sue due principali componenti.
| Modello | Comparto | Tipo strutturale |
a | b | R2 | ESS |
|---|---|---|---|---|---|---|
| y = a + d2 * b | Volume (dm3) | B | -9.484 | 0.732 | 0.997 | 0.057 |
| Volume (dm3) | C | -20.623 | 0.584 | 0.992 | 0.071 | |
| Biomassa totale (kg) | B | -198.236 | 0.620 | 0.993 | 0.064 | |
| Biomassa totale (kg) | C | -54.463 | 0.462 | 0.986 | 0.070 | |
| Biomassa fotosintetizzante (kg) | B | -61.386 | 0.095 | 0.965 | 0.016 | |
| Biomassa fotosintetizzante (kg) | C | -6.723 | 0.054 | 0.983 | 0.009 | |
| Biomassa legnosa (kg) | B | -136.850 | 0.526 | 0.993 | 0.055 | |
| Biomassa legnosa (kg) | C | -47.740 | 0.408 | 0.985 | 0.065 | |
| y = a + d2h* b | Volume (dm3) | B | -14.214 | 0.041 | 0.998 | 0.003 |
| Volume (dm3) | C | -4.198 | 0.038 | 0.995 | 0.004 | |
| Biomassa totale (kg) | B | -170.371 | 0.034 | 0.993 | 0.004 | |
| Biomassa totale (kg) | C | -41.906 | 0.030 | 0.989 | 0.004 | |
| Biomassa totale (kg) | B+C | -42.009 | 0.029 | 0.990 | 0.004 | |
| Biomassa fotosintetizzante (kg) | B | -54.357 | 0.005 | 0.959 | 0.001 | |
| Biomassa fotosintetizzante (kg) | C | -5.102 | 0.003 | 0.987 | 0.001 | |
| Biomassa legnosa (kg) | B | -116.014 | 0.029 | 0.994 | 0.003 | |
| Biomassa legnosa (kg) | C | -36.805 | 0.026 | 0.988 | 0.004 | |
| Biomassa legnosa (kg) | B+C | -36.305 | 0.026 | 0.990 | 0.003 |
La Tab. 6, oltre alcune variabili dendrometriche qualificanti i popolamenti oggetto di studio, riporta i valori di BEF relativi a ciascun tipo strutturale. Da notare come il Tipo B presenti valori superiori al Tipo C, mentre la media dei due valori è pari a 1.36.
Tab. 6 - - Caratteristiche dei popolamenti studiati e expansion factor per la biomassa epigea (BEF).
| Tipo strutturale |
Età | Densità (n ha-1) |
Comparto | Biomassa secca (Mg*ha-1) |
B.E.F. |
|---|---|---|---|---|---|
| B | 62 | 338 | Fusto | 173.1 | 1.39 |
| Totale | 240.8 | ||||
| C | 50 | 370 | Fusto | 132.4 | 1.33 |
| Totale | 176.1 |
Discussioni e Conclusioni
I risultati ottenuti e, in particolare la serie di modelli di previsione della biomassa totale e per componenti, seppure relativi ad uno specifico caso di studio, costituiscono non solo uno strumento di notevole utilità sotto il profilo tecnico, ma anche un contributo non trascurabile in termini conoscitivi. La rilevanza di ciò è accresciuta sia dalla sporadicità degli studi e dall’esiguità di strumenti di stima ad oggi disponibili per il pino domestico, che dalla diffusione assolutamente non trascurabile di questa specie nel bacino del Mediterraneo.
Detto ciò, i risultati ottenuti consentono di fare alcuni approfondimenti sulle relazioni tra struttura del popolamento e allocazione della biomassa da un lato, e sull’accuratezza dei modelli di previsione dall’altro.
La differente pressione competitiva, evidenziata dagli indicatori dendrometrici relativi ai due tipi strutturali esaminati, ha condizionato in primo luogo l’accrescimento longitudinale e, in secondo luogo, l’allocazione della biomassa. Nella pineta caratterizzata da semina su fasce lavorate alternate a fasce salde (Tipo B) si è instaurata nel tempo una pressione competitiva più uniforme rispetto al Tipo C, dove la giustapposizione di nuclei di pino e di leccio ha determinato una competizione spazialmente più disomogenea. Questo ha indotto uno sviluppo in altezza più uniforme, peraltro tipico della specie allorché allevata in condizioni di elevata densità. Al contrario nel Tipo C, dove il pino domestico è stato seminato in maniera andante e a integrazione di nuclei di latifoglie preesistenti, con conseguente maggiore variabilità da punto a punto dei livelli di competizione, si è registrata una maggiore differenziazione delle altezze, con valori distribuiti in un intervallo di circa quattro metri (13-17 m).
Quanto sopra descritto ha trovato puntuale riscontro nelle relazioni ipsodiametriche relative a ciascun tipo strutturale. Relazione ipsodiametrica praticamente inesistente nel caso del Tipo B e, al contrario, statisticamente significativa nel Tipo C. Risultati questi che, peraltro, pongono non pochi problemi sotto il profilo metodologico, dal momento che l’impiego di modelli di previsione del volume e della biomassa a due entrate, notoriamente ritenuti molto più accurati di quelli a una entrata, presuppongono l’elaborazione o l’utilizzo di relazioni ipsodiametriche. Relazioni ipsodiametriche che, qualora aleatorie come nel Tipo B, possono introdurre errori non trascurabili o comunque superiori all’aumento di accuratezza ottenuto con la scelta del modello a due entrate. La rilevanza di questo risultato è accresciuta dal fatto che questo studio è esente dagli errori che in genere affliggono le misure di altezza su piante in piedi, dal momento che la base dei dati su cui si è operato è costituita da misure di altezze raccolte a terra sugli alberi campione abbattuti. Da ciò ne consegue che nel caso del pino domestico, più che in altre specie, l’opzione a favore dei modelli di previsione a due entrate dovrebbe avvenire non in maniera pregiudiziale, ma dopo un attento esame della robustezza delle relazioni ipsodiametriche disponibili e della loro rispondenza alle reali caratteristiche del popolamento.
Oltre agli effetti sullo sviluppo longitudinale sono emerse alcune differenze tra i due tipi strutturali relativamente all’allocazione della biomassa. Mentre la componente rami fini e aghi ha valori praticamente uguali nei due tipi strutturali, si registrano invece differenze significative tra biomassa del fusto e dei rami grossi. Nel Tipo B il fusto costituisce il 72% del totale, mentre nel Tipo C il 77%. Al contrario la biomassa dei rami grossi è pari al 17% nel Tipo B e all’11% nel Tipo C. Anche in questo caso è lecito ritenere che le differenze nei livelli di competizione, conseguenza delle iniziali modalità di impianto e dell’assenza di un organico trattamento nelle fasi successive, abbiano condizionato l’occupazione dello spazio da parte del popolamento, con un conseguente diverso sviluppo delle chiome.
Tutto ciò spiega gli esiti dei test statistici sui modelli di previsione del volume e della biomassa. Test che hanno messo in luce effetti significativi della struttura del popolamento sui modelli di previsione, sia a una che a due entrate. Nel caso dei modelli a una entrata gli effetti sono tali da suggerire sempre l’utilizzo di modelli di previsione specifici per ciascun tipo strutturale. Nel caso dei modelli a due entrate l’inserimento dell’altezza come variabile indipendente mitiga gli effetti della struttura: per la stima della biomassa legnosa e totale è infatti statisticamente accettabile l’utilizzo di un’unica relazione allometrica, ovviamente tenendo distinte le equazioni ipsometriche per ciascun tipo strutturale.
Inevitabilmente quanto sopra riportato ha condizionato anche il BEF, che peraltro, nel caso di Castelfusano, mostra valori in linea con quelli riportati per il genere Pinus in diversi paesi europei ([23]). Le differenze osservate tra i due tipi strutturali vanno a sostegno e integrazione di quanto emerso in studi recenti, che hanno evidenziato differenze nei valori di BEF non solo a livello di specie ma anche di età del popolamento ([22], [19]) e, quindi, di caratteristiche strutturali.
Nel complesso i modelli di previsione del volume e della biomassa e i valori di BEF proposti rappresentano, oltre che strumenti di interesse tecnico-applicativo e una valida base per le ricerche attualmente in corso nella pineta di Castelfusano, un contributo utile a colmare l’attuale vuoto di conoscenze sul pino domestico e su formazioni di notevole valenza come le pinete litoranee. Ben lungi da mettere in dubbio l’utilità di strumenti di stima validi su ampia scala e gli sforzi per arrivare ad una semplificazione dei modelli predittivi ([29], [27]), quanto emerso nel presente studio richiama la necessità di non sottovalutare gli effetti della struttura sullo sviluppo longitudinale e, quindi, sulle relazioni ipsodiametriche, sull’allocazione della biomassa nei vari comparti, sull’accuratezza dei modelli di previsione e sui valori di BEF, soprattutto laddove si miri a stime particolarmente accurate o distinte per i vari comparti. Le peculiarità del pino domestico, fanno si che una superficiale analisi della struttura, più che per altre specie, possa indurre errori rilevanti sotto il profilo scientifico e assolutamente non trascurabili anche sotto il profilo tecnico.
Ringraziamenti
La ricerca è stata finanziata dal Comune di Roma, Dipartimento X Politiche Ambientali e Risorse Agricole nell’ambito della convenzione “Gestione delle pinete di pino domestico (Pinus pinea L.) della Riserva Naturale Statale di Castelfusano” e dal Mi.P.A.F. nell’ambito del progetto Ri.SELV.ITALIA - sottoprogetto 3.1 - Selvicoltura, funzionalità e difesa degli ecosistemi forestali - 3.1.1 Conservazione e miglioramento delle pinete litoranee.
Si ringrazia sentitamente il dott. Alessandro De Michelis, direttore della Tenuta Presidenziale di Castelporziano, per la disponibilità assicurata nel corso della ricerca.
Si ringrazia altresì il personale del CRA - Centro di Ricerca per la Selvicoltura che a vario titolo ha collaborato alla realizzazione della ricerca, con una menzione particolare per i collaboratori Umberto Cerofolini, Andrea Ciofini, Luigi Mencacci, e Maurizio Piovosi.
Particolarmente sentito è il ringraziamento che va al dott. Giovanni Tabacchi per i consigli, l’assistenza e l’aiuto per l’elaborazione delle relazioni allometriche e la metodologia ad esse correlata.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar