Birds and structure in the Ragabo pine forest (Linguaglossa, CT - Italy)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 7, Pages 223-233 (2010)
doi: https://doi.org/10.3832/efor0633-007
Published: Oct 13, 2010 - Copyright © 2010 SISEF
Research Articles
Guest Editors: 7° SISEF Congress (Pesche, IS - 2007)
« Development, adaptation, naturality and conservation »
Collection/Special Issue: Marco Marchetti, Roberto Tognetti
Abstract
Structural analysis of a forest ecosystem may allow to assess the spatial heterogeneity and temporal dynamics of the vegetation, the factors related to tree recruitment and the dynamics of canopy gaps, as well as to define the niche of wildlife. The study of the ecosystem response to different silvicultural treatments may also provide hints of the bio-ecological functionality and forest ecosystem stability. In this work, we analyzed the most representative structural types within the Ragabo pine wood (monoplane forest and multilayered forest), and their relationships with the specific composition of the local breeding bird community. The results show the strong sensitivity of birds to environmental changes and their large selectivity in the choice of habitats. In particular uneven-aged forests remain particularly suited to maintain over time a bird community richer and more stable than the even-aged forests.
Keywords
Calabrian pine, Bird community, Stand structure, Sustainable management, Ragabo wood
Introduzione
Il pino laricio (Pinus laricio Poiret), sottospecie endemica dell’Appennino Calabro e dell’Etna in Sicilia, è una delle specie più importanti della selvicoltura dell’Italia meridionale.
In Calabria le pinete naturali si trovano in Sila, dove caratterizzano in modo significativo il paesaggio forestale tra 1200 e 1600 m di quota e coprono una superficie di circa 97000 ettari ([4]), e sui versanti meridionali dell’Aspromonte, dove interessano circa 3000 ettari tra 1250 e 1600 m di quota. In Sicilia vegeta quasi esclusivamente sull’Etna e caratterizza alcuni ambiti territoriali distribuiti in modo non uniforme da 1000 a 2000 m di quota. Interessa una superficie di 5654 ettari ([10]), 1200 dei quali nel solo demanio del Comune di Linguaglossa. Si tratta per la maggior parte (87%) di popolamenti puri ([10]).
L’importanza delle pinete di laricio della Sila, dell’Aspromonte e dell’Etna è accresciuta dal loro corteggio floristico costituito in buona parte da specie endemiche o esclusive di queste formazioni e dalle specie animali ad esse legate. La fauna invertebrata presenta interessanti endemismi ([22]). Molte specie trovano in queste aree il limite meridionale di diffusione. Tra gli uccelli vanno menzionate alcune specie, la cui presenza è strettamente legata alle pinete di laricio, quali: il regolo (Regulus regulus), segnalato in Calabria esclusivamente nelle pinete di laricio della Sila e dell’Aspromonte; il lucherino (Carduelis spinus) che in Sicilia si riproduce proprio nelle pinete dell’Etna ([12]); il crociere (Loxia curvirostra) la cui presenza di nuclei stabili, a queste latitudini, è vincolata alle formazioni di laricio.
Nel passato la sostenibilità ambientale della gestione di queste formazioni non era considerata un aspetto primario. L’istituzione di aree protette che inglobano la quasi totalità delle pinete naturali di laricio, ha rilanciato il problema della gestione sostenibile e della conservazione o dell’aumento della biodiversità di questi sistemi.
L’applicazione di forme di trattamento basate sui modelli prescritti dalla selvicoltura classica ha portato ad una estrema semplificazione delle condizioni strutturali e ad una riduzione della biodiversità di gran parte delle pinete di laricio. In alcuni ristretti ambiti si sono conservate delle forme di “selvicoltura tradizionale”, descritte per la Sila da Ciancio et al. ([5]), che hanno garantito il mantenimento di una struttura disetanea di tipo atomistico salvaguardando l’efficienza del bosco attraverso la rinnovazione naturale e il mantenimento, sull’unità di superficie, di una provvigione minima, garanzia contro i rischi di degrado del suolo e di depauperamento dell’ecosistema.
La struttura verticale del bosco è, uno dei fattori che maggiormente influisce sulla composizione e sulla densità delle comunità ornitiche ([6]). La diversità specifica delle comunità aumenta in misura proporzionale alla complessità degli habitat. I boschi maturi, caratterizzati da una maggiore complessità strutturale rispetto a quelli giovani (maggior numero di strati, più abbondante sottobosco, ecc.), favoriscono la presenza di popolamenti ornitici più ricchi di specie ([23]). Il numero di specie di uccelli presenti e l’abbondanza degli effettivi di popolazione sono strettamente correlati alla biodiversità complessiva dell’ambiente.
Con il presente lavoro si riportano i primi risultati di uno studio volto alla valutazione degli effetti, sulla struttura dei soprassuoli e sulla composizione specifica della comunità ornitica, di due forme di gestione tradizionalmente applicate nella selvicoltura mediterranea: il taglio a raso con rilascio di riserve e il taglio a scelta.
Si fa presente che trattasi di risultati preliminari che in un successivo contributo andranno ampliati e corroborati.
Area di studio
I popolamenti oggetto di studio fanno parte del demanio del comune di Linguaglossa (CT). La pineta è interamente compresa entro i confini del parco regionale dell’Etna e nel SIC ITA070013 “Pineta di Linguaglossa”.
I valori di temperatura e precipitazioni di riferimento per l’area oggetto di studio sono quelli ricavati da Santocono ([20]) per la stazione Rifugio Conti, posta a 1589 m s.l.m., in posizione baricentrica rispetto alla pineta Ragabo. Per questa stazione l’Autore ha individuato un valore di temperatura media annua di 8.3°C; le precipitazioni presentano il tipico andamento del clima mediterraneo e ammontano complessivamente a 1200 mm annui. Secondo la classificazione fitoclimatica di Pavari, l’area rientra nella zona del Fagetum, sottozona calda con transizione verso quella fredda; secondo Schmidt ricade nel cingolo Fagus - Abies (F.A. - [20]). Secondo Rivas Martinez, il bioclima è supramediterraneo umido.
L’indice di aridità di De Martonne per la stazione Rifugio Conti è superiore a 60, pertanto la zona è da considerarsi perumida.
Dal punto di vista geologico, l’area è interessata da colate laviche dell’ellittico (Mongibello antico, Pleistocene - [18]), da cui hanno avuto origine suoli riferibili, secondo la Soil Taxonomy, all’associazione dei regosuoli, litosuoli e suoli bruni andici ([8]). Tipico di quest’area è anche il continuo apporto di ceneri e lapilli dovuto all’attività vulcanica.
Dal punto di vista fitosociologico, le pinete di laricio dell’Etna sono riferibili allo Junipero hemisphaericae - Pinetum calabricae, associazione esclusiva di questa zona della Sicilia ([3]). I condizionamenti ecologici conseguenti all’attività del vulcano (deposizione di strati di lapilli, incendi, ecc.) non permettono l’evoluzione dei suoli e la pineta assume il significato di un climax edafico stabile nel tempo, con un ruolo ecologico oltre che paesaggistico di grande rilievo ([17]).
Nelle condizioni favorevoli per l’evoluzione del suolo, la pineta rappresenta uno stadio dinamico che evolve, a seconda dell’altitudine, verso i boschi di querce caducifoglie a quote più basse o popolamenti di faggio a quote più elevate ([14]).
Il soprassuolo è costituito da pino laricio allo stato puro. Alle quote inferiori al pino, si associano il castagno (Castanea sativa, Mill.) e la roverella (Quercus pubescens Willd.); verso l’alto il faggio (Fagus sylvatica L.) e il pioppo tremulo (Populus tremula, L.) e, ai limiti superiori della vegetazione forestale, la betulla dell’Etna (Betula aetnensis Raf.).
In queste pinete fino alla metà del secolo scorso veniva effettuata, sui fusti di maggiori dimensioni, la resinazione, pratica che anticipa la morte delle piante. Dagli anni ’70 del secolo scorso gli interventi sul soprassuolo sono stati limitati all’asportazione delle piante morte o deperienti di dimensioni medio-grandi, la ramaglia e i fusti di piccole dimensioni venivano lasciati in bosco.
Materiali e metodi
All’interno della pineta sono state scelte due zone le cui differenze, per quanto riguarda la struttura e i parametri dendrometrici, sono attribuibili alle due forme di trattamento applicate. Nel primo caso il trattamento applicato è il taglio a raso con riserva di 50-60 piante a ettaro, nel secondo caso una forma di gestione tradizionale riconducibile al taglio a scelta a piccoli gruppi.
Nelle due zone sono state individuate 25 stazioni di ascolto per il rilievo dei parametri necessari alla caratterizzazione della comunità ornitica nidificante.
Rilievi
Per la descrizione delle caratteristiche dendrometriche e strutturali dei soprassuoli sono state delimitate 25 aree di saggio di forma circolare e diametro variabile in funzione della densità del soprassuolo. La superficie media delle aree è 2928 m2 e l’area campionata interessa complessivamente una superficie di 73199 m2.
In ogni area sono stati rilevati i diametri di tutte le piante a partire da una soglia minima di cavallettamento di 2.5 cm. Inoltre, è stato misurato un congruo numero di altezze, distribuite in tutte le classi di diametro, utilizzate per la costruzione della curva ipsometrica.
Per la caratterizzazione della struttura verticale e l’individuazione degli strati verticali con il metodo proposto da Latham et al. ([11]), all’interno di transect di 500 m2 (50 × 10 m) ricadenti nelle aree di saggio circolari, sono stati rilevati il diametro, l’altezza totale e di inserzione della chioma verde di tutte le piante.
Per la rappresentazione grafica del bosco, in ciascuno dei transect è stata rilevata la posizione di tutte le piante mediante le coordinate polari riferite a un vertice, ne è stato misurato il diametro a 1.30 m da terra (a partire da una soglia minima di cavallettamento di 2.5 cm), l’altezza totale e quella del punto di inserzione della chioma verde. Di ciascuna chioma sono stati misurati anche i quattro raggi secondo i punti cardinali.
Per la determinazione dell’età convenzionale delle piante, nel popolamento coetaneo, per ogni classe di diametro, è stato individuato un campione di alberi, rappresentativi delle condizioni medie del soprassuolo, da ciascuno dei quali è stata prelevata, a 1.30 m da terra, una carotina. Nei soprassuoli disetanei sono state analizzate tutte le piante con diametro superiore ai 7.5 cm, prelevando una carotina, mentre, per le piante con diametro compreso tra 2.5 e 7.5 cm sempre riunite in gruppi, sono state prelevate delle rotelle da alcuni individui rappresentativi del gruppo stesso. I dati, ottenuti dalla lettura delle carotine e delle rotelle, sono stati utilizzati per la costruzione dei modelli di struttura cronologica del soprassuolo.
Per quanto riguarda i rilievi sulla rinnovazione (sono state classificate come rinnovazione tutte le piante che non raggiungevano il diametro di 2.5 cm a 1.30 m da terra), all’interno di ogni transect è stato contato il numero di individui presenti, distintamente per specie, ne è stata misurata l’altezza e sono stati localizzati sul terreno.
Il rilevamento dell’avifauna è stato effettuato applicando il metodo dei punti d’ascolto ([1]). Il punto di ascolto coincide con il centro delle aree di saggio circolari, la durata dell’ascolto è di 10’ per ogni osservazione, con una distanza minima di 200 m tra due punti successivi. Per ridurre al minimo l’effetto “margine”, i punti sono stati individuati a una distanza minima di 100 m dai margini del bosco e dalla strada provinciale che lo attraversa.
Ai fini dell’elaborazione dei dati raccolti e per la valutazione della presenza dell’avifauna sono stati presi in considerazione solamente i contatti avvenuti entro un raggio di 100 m dal punto di ascolto. Nel periodo di osservazione dal 17/04/2009 al 26/05/2009 sono state effettuate 2 ripetizioni per poter individuare sia le specie nidificanti precoci che quelle tardive, condotte dallo stesso rilevatore, in un intervallo di tempo di 3.5 ore a partire dalle prime luci dell’alba e mai oltre le ore 9.00 orario solare. Inoltre, i rilievi sono stati effettuati in assenza di precipitazioni e con condizioni di vento favorevoli (< 4 della scala di Beaufort). La posizione di tutte le stazioni di ascolto è stata registrata mediante un ricevitore GPS.
Sulla base dei dati raccolti è stata redatta la check-list delle specie nidificanti e, cumulando i dati delle osservazioni, è stato calcolato, per ciascuna specie, il valore di frequenza percentuale (FP) in tutte le stazioni visitate e il valore di dominanza (pi) ottenuto dividendo FP per la sommatoria delle frequenze percentuali di tutte le specie.
Le specie presenti in almeno metà delle stazioni sono state considerate comuni, mentre quelle aventi pi*100 > 5 sono state considerate dominanti ([21]).
Per la descrizione della struttura della comunità ornitica sono stati utilizzati i seguenti parametri:
- S = ricchezza;
- s = ricchezza media per stazione;
- H’ = diversità specifica (indice di Shannon);
- NP% = percentuale di specie non-passeriformi;
- J = equiripartizione (H’/H’ max, dove H’ max = lnS, [16]);
- NC = numero di specie comuni;
- ND = numero di specie dominanti.
Tali parametri, recentemente applicati in studi analoghi ([7], [9], [19], etc.), consentono una immediato confronto tra le caratteristiche di diverse comunità ornitiche.
Risultati
Fustaia coetanea
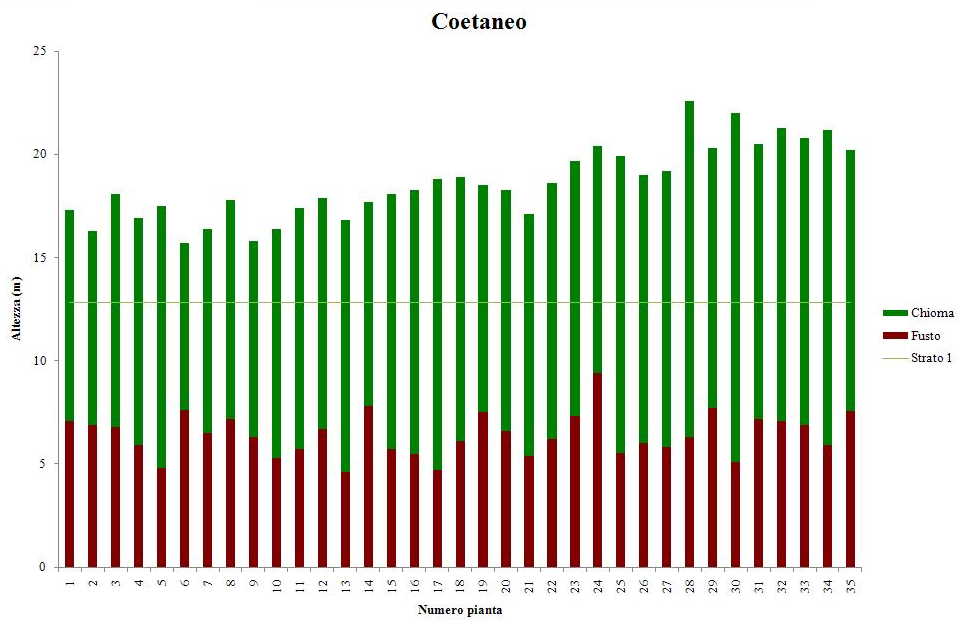
Si tratta di una fustaia coetanea di 110 - 140 anni di età, trattata a taglio raso con riserva di 50 - 60 piante ad ettaro. Il soprassuolo è stato sottoposto a interventi di diradamento di tipo basso e di grado debole, ripetuti ogni 25 anni, e finalizzati a regolare la densità del popolamento. I fusti presentano una forma da buona a discreta, la chioma è inserita nel terzo superiore. Il sottobosco è formato quasi esclusivamente da felce aquilina (Pteridium aquilinum (L.) Kuhn) e brachipodio pinnato (Brachypodium pinnatum (L.) P. Beauv.).
Il soprassuolo è costituito mediamente da 388 piante a ettaro, di cui il 95% pino laricio e il restante 5% costituito da roverella e betulla dell’Etna. L’area basimetrica complessiva ammonta a 64.36 m2 a ettaro e il volume a 831.1 m3. Il diametro della pianta di pino laricio di dimensioni medie misura 47.76 cm e 27.4 m di altezza (Tab. 1). Il contributo in termini di area basimetrica e volume delle latifoglie è trascurabile (<1%).
Tab. 1 - Parametri dendrometrici fustaia coetanea. (*): il valore Min è uguale a zero in quanto la roverella non era presente in tutte le aree di saggio.
| Specie | Parametri statistici |
N piante |
Dg (m) |
Hg (m2) |
G (m2) |
V (m3) |
|---|---|---|---|---|---|---|
| Pino laricio | Media | 367 | 47.76 | 27.4 | 64.11 | 830.1 |
| Dev st | 79 | 5.46 | 1.2 | 8.79 | 124.2 | |
| Min | 247 | 38.06 | 25.0 | 53.14 | 683.2 | |
| Max | 527 | 56.31 | 29.2 | 82.84 | 1069.2 | |
| Roverella | Media | 21 | 5.09 | 2.1 | 0.25 | 1.1 |
| Dev st | 34 | 6.91 | 2.7 | 0.43 | 1.9 | |
| Min* | 0 | 0 | 0 | 0 | 0 | |
| Max | 95 | 16.25 | 5.7 | 1.32 | 5.9 | |
| Totale | Media | 388 | - | - | 64.36 | 831.1 |
| Dev st | 71 | - | - | 8.69 | 123.9 | |
| Min | 310 | - | - | 53.33 | 684.0 | |
| Max | 559 | - | - | 82.84 | 1069.2 |
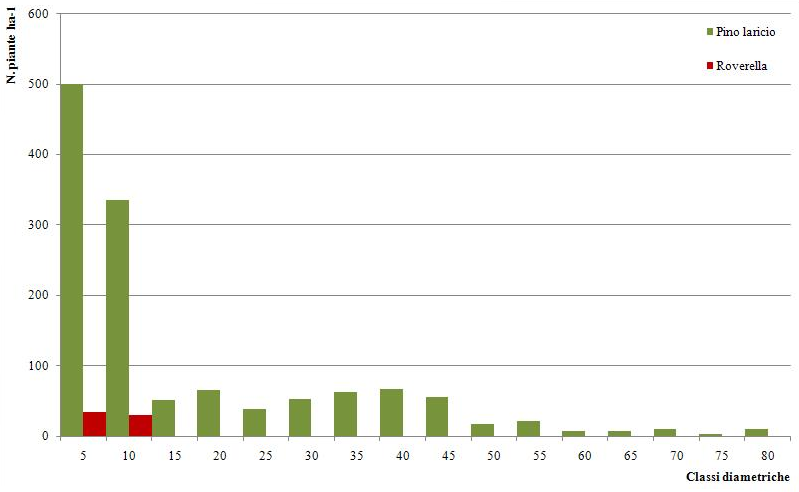
Il campo di variazione dei diametri è tra 5 e 85 cm. La distribuzione delle piante in classi di diametro presenta andamento a campana con asimmetria a sinistra e moda nella classe di 45 cm di diametro (Fig. 1).
Fig. 1 - Distribuzione delle piante in classi diametriche, fustaia coetanea.
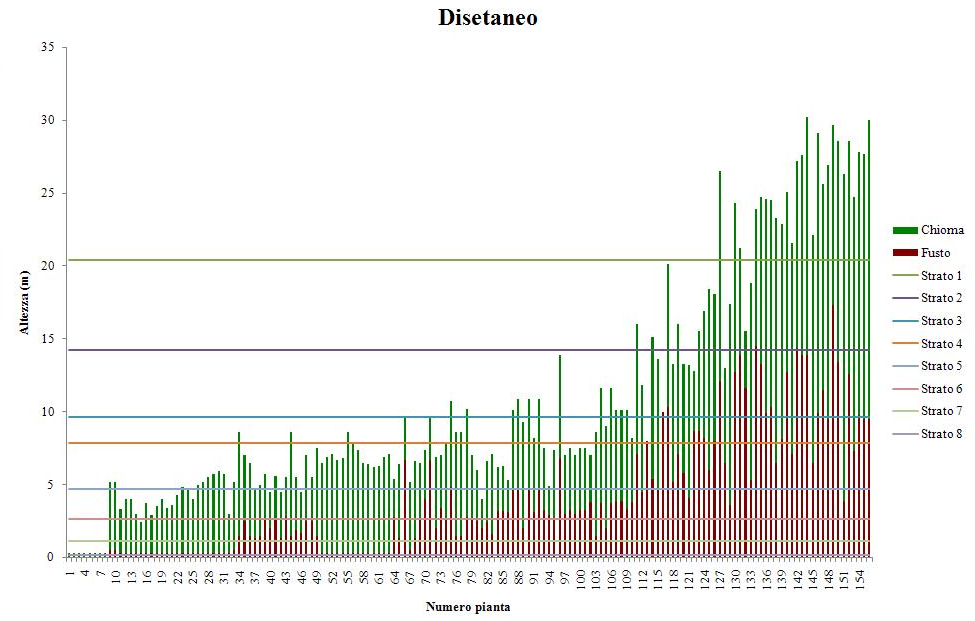
L’analisi della struttura verticale, attraverso l’applicazione dell’indice di Latham, evidenzia una ridotta articolazione del soprassuolo (Fig. 2) che è in genere monoplano ed, in alcuni casi, ovvero nei soprassuoli di maggiore età, comincia a differenziarsi in un piano dominante e uno dominato. La presenza di un terzo strato, sotto copertura, è legata ad occasionali fenomeni di insediamento del novellame.
Fig. 2 - Stratificazione verticale secondo Latham, fustaia coetanea.
L’analisi delle carotine prelevate conferma la coetaneità di questi boschi; le età convenzionali lette sono comprese tra i 110 ed i 140 anni, valori che coincidono con quelli riportati nel piano di assestamento redatto per la pineta ([15]).
Allo stato attuale, in assenza di interventi colturali, i processi di evoluzione naturale procedono molto lentamente a causa della concorrenza esercitata dal piano dominante. Le latifoglie presenti all’interno del popolamento non sembrano avere capacità di ulteriore accrescimento. I piccoli gaps che si aprono casualmente per la caduta di singole piante sono favorevoli all’insediamento e alla rinnovazione del pino laricio. Le occasionali interruzioni della copertura sono causate dal crollo di singole piante di grandi dimensioni dalle quali, fino alla metà del secolo scorso, veniva estratta la resina. Tale pratica, infatti, anticipa la morte della pianta.
La necromassa a terra è costituita quasi esclusivamente da ramaglia minuta, più raramente da giovani piante morte per aduggiamento, oltre che dagli aghi che annualmente cadono al suolo. I processi di decomposizione procedono abbastanza rapidamente per le favorevoli condizioni microclimatiche che si instaurano sotto copertura. Raramente si osservano piante di dimensioni medie e grosse morte in piedi o cadute a terra.
Fustaia disetanea
L’applicazione di una forma di trattamento riconducibile al taglio a scelta, con la quale venivano selezionate e prelevate le piante di maggiori dimensioni, ha consentito di ottenere popolamenti disetanei a struttura complessa. Patrone ([15]) ha definito questi boschi disformi nei riguardi sia della densità che della struttura a causa dei tagli effettuati.
L’insediamento della rinnovazione era legato all’esecuzione degli interventi di utilizzazione e alle condizioni dei suoli, che in quest’area risultano poco evoluti e ricchi di scheletro. Già nella seconda metà del secolo scorso erano presenti piante con età comprese tra 5 e oltre 200 anni. Inoltre, i processi di rinnovazione sono proseguiti fino a circa 20 anni fa. L’assenza o la scarsità di soggetti con età inferiore ai 20 anni è, infatti, da imputare principalmente alla sospensione degli interventi di utilizzazione o di asportazione delle piante morte avvenuta a seguito dell’istituzione del Parco Regionale dell’Etna. La rinnovazione costituisce dei piccoli gruppi, piuttosto densi, irregolarmente distribuiti all’interno del popolamento in rapporto a quelli che sono stati gli interventi di utilizzazione.
Mediamente il soprassuolo è costituito da 955 piante a ettaro; considerando solo i soggetti con diametro superiore a 17.5 cm, la densità si riduce a 400 piante a ettaro. Complessivamente l’area basimetrica ammonta a 49.15 m2 ad ettaro e il volume a 453.6 m3. L’altezza media delle piante di pino (16.8 m) è fortemente condizionata dalla elevata densità del soprassuolo e dai diametri modesti delle piante (Tab. 2). La presenza di latifoglie è molto scarsa e si tratta sempre di piante di piccole dimensioni localizzate in prossimità dei fossi o sotto la copertura delle piante di dimensioni più grandi.
Tab. 2 - Parametri dendrometrici fustaia disetanea. (*): il valore Min è uguale a zero in quanto non erano presenti roverelle o altre specie in tutte le aree di saggio.
| Specie | Parametri statistici |
N piante |
Dg (m) |
Hg (m2) |
G (m2) |
V (m3) |
|---|---|---|---|---|---|---|
| Pino laricio | Media | 924 | 26.69 | 16.8 | 48.87 | 452.6 |
| Dev st | 924 | 26.69 | 16.8 | 48.87 | 452.6 | |
| Min | 527 | 19.48 | 14.7 | 39.22 | 346.2 | |
| Max | 1467 | 33.19 | 18.4 | 67.23 | 644.6 | |
| Roverella | Media | 32 | 6.50 | 2.9 | 0.30 | 1.1 |
| Dev st | 44 | 6.29 | 2.6 | 0.37 | 1.4 | |
| Min* | 0 | 0 | 0 | 0 | 0 | |
| Max | 159 | 17.73 | 5.87 | 0.70 | 4.26 | |
| Altre specie | Media | 2 | 1.27 | 0.6 | 0.02 | 0.1 |
| Dev st | 6 | 3.57 | 1.7 | 0.05 | 0.2 | |
| Min* | 0 | 0 | 0 | 0 | 0 | |
| Max | 18 | 12.75 | 5.29 | 0.18 | 0.68 | |
| Totale | Media | 955 | - | - | 49.15 | 453.6 |
| Dev st | 247 | - | - | 7.91 | 83.8 | |
| Min | 552 | - | - | 39.39 | 346.8 | |
| Max | 1467 | - | - | 67.23 | 644.6 |
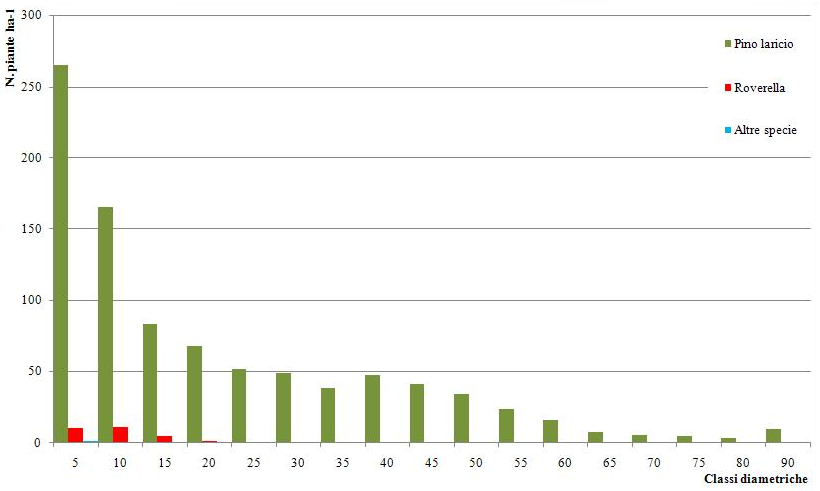
Il campo di variazione dei diametri è tra 5 e 90 cm. La distribuzione delle piante in classi di diametro evidenzia un andamento decrescente all’aumentare del diametro (Fig. 3). Considerando solamente le piante con diametro superiore a 17.5 cm, si ottiene una distribuzione piuttosto irregolare che evidenzia la presenza di gruppi di piante di diverse dimensioni fra loro giustapposti. Il sottobosco è abbastanza uniforme e risulta costituito soprattutto da graminacee.
Fig. 3 - Distribuzione delle piante in classi diametriche, fustaia disetanea.
Le analisi della struttura verticale, condotte attraverso l’indice di Latham, evidenziano la presenza di un numero elevato di strati, compreso fra 4 e 8 (Fig. 4). In tutti i casi, la struttura somatica risulta piuttosto eterogenea già su piccole superfici. La differenziazione tra gli strati superiori è sempre netta e legata alla presenza di gruppi di piante con differenti età. Negli strati inferiori la differenziazione si attenua in quanto la disponibilità di spazio si riduce e c’è una maggiore concorrenza all’interno di ciascun gruppo. La sospensione degli interventi selvicolturali ha certamente condizionato anche la crescita delle piante nel gruppo e bloccato i processi di insediamento della rinnovazione. La densità all’interno dei gruppi risulta sempre elevata, per cui anche la mortalitàè alta. Inoltre, la presenza degli strati superiori, nei quali non vengono più effettuati interventi, limita l’accrescimento dei soggetti più vigorosi presenti nei gruppi sottostanti.
Fig. 4 - Stratificazione verticale secondo Latham, fustaia disetanea.
Il range di variazione dell’età convenzionale è compreso tra 20 e 140 anni. Sulla base dei rilievi effettuati è stato possibile distinguere 13 classi di età, di ampiezza pari a 10 anni. Le frequenze più elevate si osservano nelle classi cronologiche di 30 e 40 anni (Fig. 5 e Fig. 6). Poche sono le piante che rientrano nella classe cronologica di 20 anni, mentre sono completamente assenti quelle di età inferiore a 20 anni. Dalla lettura degli anelli nella zona del colletto è emerso che anche i soggetti che non raggiungono la soglia minima di cavallettamento di 2.5 cm hanno comunque più di 20 anni di età.
Fig. 5 - Distribuzione delle piante in classi cronologiche.
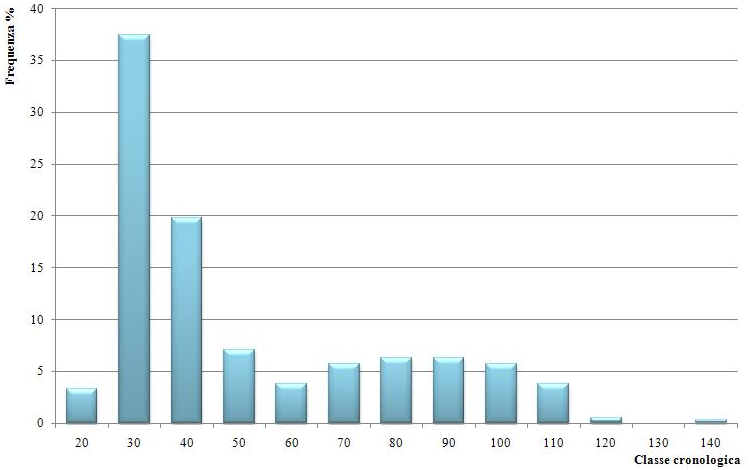
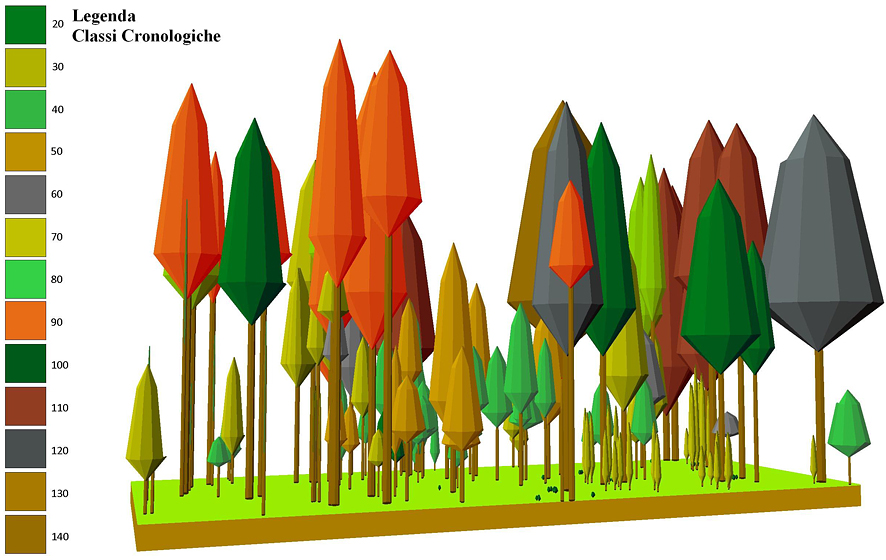
Fig. 6 - Modello di struttura cronologica.
Avifauna
Nelle 25 stazioni di ascolto sono state censite un totale di 23 specie nidificanti. Nei conteggi non sono stati inseriti i rapaci diurni e notturni rilevati.
Per la classificazione dell’avifauna presente è stata utilizzata la check-list degli uccelli italiani di Brichetti & Massa ([2]). Le specie nidificanti nella Pineta Ragabo appartengono a quattro ordini e a dodici famiglie; quattro sono le specie di Non-Passeriformi.
Cumulando i dati dei due rilevamenti effettuati, è stato calcolato per tutte le specie il valore di frequenza percentuale nelle stazioni visitate. Sulla base dei risultati ottenuti, emerge che 3 specie sono presenti in più del 90% delle stazioni. Inoltre altre 8 specie sono presenti in più di metà delle stazioni (Tab. 3).
Tab. 3 - Frequenza percentuale (F%), dominanza (pi·100) e relativa guild di appartenenza delle specie nidificanti.
| Specie | F% | F% Coetaneo |
F% Disetaneo |
pi*100 | Guilds |
|---|---|---|---|---|---|
| Columba palumbus | 92 | 90 | 93 | 9.31 | GF |
| Cuculus canorus | 12 | 0 | 20 | 1.06 | CF |
| Jynx torquilla | 12 | 0 | 20 | 0.80 | GF |
| Dendrocopos major | 88 | 80 | 93 | 8.24 | BF |
| Troglodytes troglodytes | 16 | 0 | 27 | 1.33 | GF |
| Erithacus rubecula | 64 | 10 | 100 | 6.65 | GF |
| Turdus merula | 12 | 0 | 20 | 1.33 | GF |
| Turdus viscivorus | 44 | 50 | 40 | 3.99 | GF |
| Sylvia atricapilla | 56 | 40 | 67 | 5.85 | CF |
| Phylloscopus collybita | 24 | 0 | 40 | 2.13 | CF |
| Regulus ignicapillus | 28 | 10 | 40 | 2.93 | CF |
| Parus ater | 100 | 100 | 100 | 9.84 | CF |
| Parus caeruleus | 72 | 50 | 87 | 6.38 | CF |
| Parus major | 60 | 10 | 93 | 5.32 | CF |
| Sitta europea | 28 | 20 | 33 | 3.72 | BF |
| Certhia brachydactyla | 56 | 70 | 47 | 5.05 | BF |
| Garrulus glandarius | 68 | 40 | 87 | 5.85 | CF |
| Pica pica | 4 | 0 | 7 | 0.53 | GF |
| Corvus cornix | 16 | 0 | 27 | 1.33 | GF |
| Sturnus unicolor | 4 | 10 | 0 | 0.27 | CF |
| Fringilla coelebs | 96 | 100 | 93 | 9.57 | GF |
| Serinus serinus | 4 | 10 | 0 | 0.80 | GF |
| Loxia curvirostra | 80 | 80 | 80 | 7.71 | CF |
I valori di frequenza percentuale, dominanza e guild di appartenenza di tutte le specie, sono riportati nella Tab. 3. I parametri della comunità ornitica sono riportati in Tab. 4.
Tab. 4 - Parametri di struttura della comunità ornitica.
| Parametri | Coetaneo | Disetaneo |
|---|---|---|
| N. stazioni | 20 | 20 |
| S | 16 | 21 |
| s | 7.70 | 12.13 |
| H’ | 2.34 | 2.74 |
| J | 0.84 | 0.90 |
| NP% | 12.50 | 14.29 |
| NC | 8 | 10 |
| ND | 10 | 10 |
Le specie rilevate sono state suddivise in guilds in base alle strategie alimentari. Sono state identificate tre guilds: GF (ground-feeders) costituito da specie che si nutrono sul terreno; CF (canopy-feeders) formato da specie che si nutrono sulla chioma; BF (bark-feeders) caratterizzato da specie che si nutrono sulla corteccia.
I valori della frequenza relativa calcolata per ciascun guild (Tab. 5) evidenziano basse densità per le specie che si alimentano sui tronchi, mentre i valori relativi alle specie che utilizzano il terreno e la copertura vegetale praticamente si equivalgono.
Tab. 5 - Frequenze percentuali per le guilds individuate.
| Guilds | Coetaneo | Disetaneo |
|---|---|---|
| GF | 26.67 | 42.86 |
| CF | 53.33 | 42.86 |
| BF | 20.00 | 14.29 |
Confrontando i dati relativi alle due tipologie strutturali risulta evidente come i popolamenti a struttura disetanea siano caratterizzati da una maggiore ricchezza di specie (21), rispetto ai popolamenti coetanei (16 specie), così come è anche nettamente superiore la presenza media per punto di ascolto con un valore di 12.10 specie per punto, a fronte di sole 7.70 specie riscontrate nei popolamenti coetanei.
Discussione e conclusioni
L’elevata articolazione della stratificazione verticale delle pinete a struttura disetanea è il fattore chiave per il mantenimento di una più ricca comunità ornitica. Infatti, un sistema in cui lo spazio verticale è completamente saturato dagli strati di vegetazione è in grado di offrire un elevato numero di nicchie ecologiche e di soddisfare le esigenze di un maggiore numero di specie di uccelli rispetto a sistemi la cui struttura è estremamente semplificata. Con i primi risultati ottenuti, le differenze rilevate, in termini di numero totale di specie ornitiche, sembrano essere attribuibili alla diversità strutturale dei due sistemi.
Nelle pinete trattate a taglio raso con riserve, i fenomeni di competizione spingono le piante a un rapido accrescimento in altezza e a uno sviluppo contenuto della chioma, la cui parte attiva si concentra nella porzione apicale dei fusti solo nei soprassuoli di maggiore età, quando l’accrescimento in altezza culmina e la copertura vegetale comincia a differenziarsi in piani si ha l’insediamento della rinnovazione. Questo fenomeno, che in natura si verifica a seguito di disturbi su ampie superfici (incendi o schianti da neve), determina una estrema omogeneità e semplificazione della struttura dei soprassuoli che occupano le superfici bruscamente denudate. L’omogeneità delle condizioni ecologiche ha ripercussioni anche sulle comunità animali che occupano il popolamento vegetale. Nelle pinete trattate con il taglio a scelta già su superfici di limitata estensione sono state rilevate tutte le fasi cronologiche caratterizzanti il ciclo di sviluppo del pino, la seriazione diametrica rilevata è sempre più ampia e sono presenti esemplari di elevate dimensioni; ciò corrisponde ad una eterogeneità, oltreché nella struttura verticale, anche in quella orizzontale. Questo fattore, in accordo con quanto riportato in letteratura ([13], [6], [23]) verificato anche in ambienti urbani ([19]), potrebbe spiegare il maggior valore del numero medio di specie di uccelli per punto di ascolto rilevato nell’area sottoposta a taglio a scelta rispetto a quella trattata con il taglio a raso con riserve.
I risultati delle analisi strutturali e faunistiche effettuate su questi soprassuoli consentono di avanzare delle ipotesi sugli effetti delle forme di trattamento applicate e i cui effetti sono ancora oggi chiaramente visibili. In un contributo successivo sarà interessante verificare se tali ipotesi verranno confermate o meno.
L’applicazione di forme di trattamento basate su schemi e modelli prestabiliti, nel caso specifico quello del bosco normale coetaneo trattato a taglio raso con riserve, ha causato un estrema semplificazione della struttura del bosco e una conseguente riduzione della biodiversità. Il taglio a raso, la cui applicazione non risponde più alle attuali esigenze di gestione dei boschi, determina una elevata azione di disturbo del sistema i cui effetti permangono per l’intero ciclo vitale delle piante e si estendendo anche alle comunità animali che questi boschi ospitano.
La forma tradizionale di utilizzazione, finalizzata esclusivamente al prelievo del materiale necessario a soddisfare le esigenze della popolazione locale, comportava limitati disturbi facilmente riassorbili dal sistema che, con una capillare applicazione, hanno favorito una grande articolazione della struttura del soprassuolo, superiore rispetto alle capacità che si riteneva potessero avere sistemi edificati da specie eliofile come il pino laricio. La struttura di sistemi così gestiti ha offerto alla comunità ornitica la possibilità di sfruttare a pieno tutto lo spazio tridimensionale offerto dal bosco.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar