Influence of forest fires on insect diversity: a case study in Apulia
Forest@ - Journal of Silviculture and Forest Ecology, Volume 8, Pages 13-21 (2011)
doi: https://doi.org/10.3832/efor0648-008
Published: Feb 17, 2011 - Copyright © 2011 SISEF
Research Articles
Abstract
In this paper we analyze the relationship between forest fires and insect diversity in the Mediterranean environment. Using “distance from the ignition point” as explanatory variable, we developed a number of regression models predicting insect abundance and family richness, as well as some of the most common measures of biodiversity (e.g., Shannon index, Simpson index, Margalef index). We report a case study in a forest area (Bosco “Il Quarto” - 600 ha) located in the National Park of Alta Murgia, Apulia Region (southern Italy). Results show a significant influence of forest fire on insect abundance and family richness and a positive linear relationship between biodiversity and distance from the point of ignition for Coleoptera. Simpson’s index, in particular, appears the most suitable indicator for measuring the effects of fires on insect biodiversity in the short term.
Keywords
Introduzione
Gli incendi costituiscono un importante fattore ecologico che condiziona l’evoluzione degli ecosistemi forestali di molte regioni biogeografiche ([10]). L’ambiente mediterraneo, in particolare, è sottoposto al verificarsi di numerosi incendi boschivi, specie nel periodo estivo, con conseguenze importanti sui processi di evoluzione e conservazione della biodiversità vegetale ed animale.
In ambito forestale, numerosi studi hanno dimostrato le conseguenze degli incendi boschivi su micro e macro-fauna, quali ad esempio: gasteropodi, artropodi ([25], [22]), mammiferi e ungulati. Le specie caratterizzate da scarsa mobilità, soprattutto terrestri e/o terricole, sono quelle maggiormente influenzate dall’azione delle fiamme, dal calore e dal fumo generato durante la combustione ([3]). A questo riguardo, Robbins & Myers ([22]) hanno dimostrato come nelle praterie nord americane l’abbondanza di insetti terricoli decresca dopo l’incendio per poi riprendersi con il riaffermarsi della vegetazione.
In alcuni casi gli incendi possono determinare cambiamenti strutturali piuttosto marcati nella struttura e nella funzionalità degli ecosistemi forestali, tali da alterare i cicli biologici di molte specie vegetali ed animali ([1]).
Le conseguenze degli incendi boschivi sulla biodiversità sono molteplici e difficili da quantificare. La stessa letteratura di settore riporta risultati non generalizzabili e per certi versi contrastanti. Alcuni autori attribuiscono agli incendi effetti positivi sul mantenimento della diversità biologica e l’eterogeneità degli habitat naturali. Altri autori considerano gli incendi in termini negativi per le conseguenze sulle specie endemiche a bassa capacità di dispersione, a vantaggio di specie esotiche e/o invasive altamente competitrici. Sulle conseguenze del passaggio del fuoco gioca un ruolo cruciale il regime degli incendi, definito da intensità, periodo dell’anno, tempo di ritorno.
In questo lavoro si presentano i primi risultati di uno studio dell’influenza degli incendi boschivi sulla biodiversità entomatica in ambiente forestale mediterraneo. Lo studio si è posto l’obiettivo di analizzare l’influenza dell’incendio sulla biodiversità dell’entomofauna in termini di abbondanza di individui e ricchezza di famiglie ad un anno dall’evento. L’analisi congiunta di queste variabili assume una rilevanza particolare se si considera la carenza di studi su questo argomento e la necessità di definire modelli predittivi della biodiversità in funzione di una o più variabili spaziali (ad es., distanza dai margini di un bosco, distanza dai centri abitati e/o dalle strade principali, distanza dal punto d’innesco dell’incendio, ecc.).
Sulla base di queste premesse, lo studio ha inteso: (1) analizzare l’influenza degli incendi boschivi sulla ricchezza e abbondanza di alcune famiglie di insetti anche attraverso l’uso di indici di biodiversità presenti in letteratura; (2) valutare la risposta di insetti appartenenti a taxa diversi. Come indicazione generale, i risultati di questo studio, seppur limitati dal breve periodo di campionamento dell’entomofauna, lasciano intendere una influenza negativa tra incendi in bosco e biodiversità entomatica ad un anno dall’evento. Nello studio si evidenzia come l’incendio riduce nel breve periodo il numero e l’abbondanza di famiglie di insetti con conseguenze negative su tutta una serie di processi ecologici legati alla presenza di questi taxa ([4]).
Materiali e metodi
Area di studio
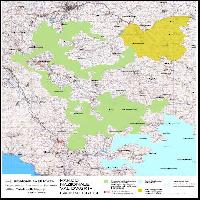
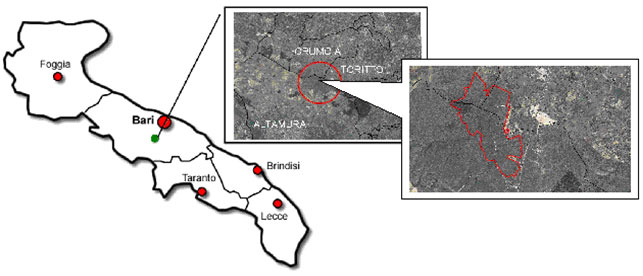
Lo studio ha riguardato un’area boschiva denominata “Il Quarto” (40° 55’ 44.08” N; 16° 36’ 52.00” E) posta all’interno del Parco Nazionale dell’Alta Murgia, nell’ambito dei comuni di Altamura, Toritto e Grumo Appula (Provincia di Bari - Fig. 1). Il bosco si estende su una superficie di circa 600 ettari ad una altitudine media di circa 400 m s.l.m. ed è governato a ceduo. Dal punto di vista vegetazionale l’area è caratterizzata da un piano dominante costituito esclusivamente da roverella (Quercus pubescens W.). Il popolamento oggetto dello studio è un ceduo invecchiato e fortemente degradato dall’eccesso di pascolamento e dalla probabile immissione precoce del pascolo dopo l’utilizzazione. La scarsa fertilità del suolo e il frequente passaggio del fuoco hanno reso più difficile l’interpretazione delle forme di gestione condotte nel passato. Le ceppaie presentano un numero medio di 4-5 polloni caratterizzati da diametri ed altezze contenuti. La densità delle ceppaie (circa 1300 ha-1) denota le scarse condizioni di fertilità della stazione. Pur essendo elevato il numero delle ceppaie le dimensioni ridotte delle chiome non assicurano la continuità della copertura del suolo che si attesta mediamente sul 50-70% della superficie del bosco. Vi sono numerose piante da rinnovazione naturale che spesso ricadono nelle classi diametriche più piccole e si mescolano ad uno strato arbustivo fitto costituito da esemplari di Pistacia lentiscus L. (lentisco), Pistacia terebinthus L. (terebinto), Rosa canina L. sensu Boulang (rosa canina), Crataegus monogyna Jacq. (biancospino), Phillyrea sp. (fillirea). Sporadicamente sono presenti matricine ravvicinate con chioma ampia alle quali si potrebbe attribuire la funzione di piante ombreggianti utilizzate in passato per il ricovero degli armenti. L’assenza di piste di passaggio del bestiame e di calpestio in prossimità di dette piante lasciano presumere che l’attività del pascolo sia stata abbandonata da circa 15-20 anni.
Fig. 1 - Area boschiva denominata “Il Quarto”, posta all’interno del Parco Nazionale dell’Alta Murgia, nell’ambito dei comuni di Altamura, Toritto e Grumo Appula (Provincia di Bari).
Nell’agosto 2008 quest’area boschiva è stato oggetto di un incendio che ha interessato circa 260 ha (43% superficie). L’incendio è stato caratterizzato da grande intensità ed elevato rilascio energetico, non controllabile con intervento diretto. Attraverso il software Visual Behave sono stati stimati alcuni parametri descrittivi del comportamento del fuoco (modello di combustibile “4” - macchia alta compatta): calore emesso per unità di superficie (29016 kJ m-2); intensità lineare (3812.5 kW m-1); lunghezza di fiamma (3.4 m). Secondo la scala di Salazar ([24]), sulla base dei valori di intensità lineare e velocità di propagazione, l’incendio in questione può essere considerato di classe di severità“estrema”. L’incendio ha infatti provocato una riduzione sensibile della vegetazione erbacea e arbustiva e una mortalità pari al 95.9 % delle piante arboree, scottate per l’intero sviluppo in altezza.
Al fine di stimare la biomassa presente nelle aree circostanti il perimetro dell’incendio, sono state individuate due aree di saggio circolari di 400 m2. All’interno delle suddette aree sono state misurate altezza e diametro di 239 piante. I risultati hanno evidenziato un’altezza media pari a 5.01 ± 1.64 m e 5.25 ± 1.71 m e un diametro medio pari a 7.38 ± 2.70 cm e 6.79 ± 2.86 cm, rispettivamente nella prima e nella seconda area di saggio.
Raccolta dei dati
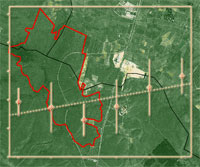
La biodiversità entomatica è stata valutata attraverso la raccolta di insetti per mezzo di trappole a caduta (pit-fall traps). La metodologia impiegata è stata quella descritta in studi analoghi, vedi Rainio & Nimelä ([21]). Nell’area in questione sono state posizionate 6 trappole lungo 3 transetti disposti a circa 120° l’uno dall’altro e secondo un gradiente vegetazionale “interno-esterno” area incendiata. Le trappole sono state disposte ad una distanza regolare di 100 m a partire dal punto presunto di innesco dell’incendio (40° 55’ 43.63” N; 16° 36’ 52.05” E) riportato nelle schede A.I.B. del Corpo Forestale dello Stato. Nell’ambito di ciascun transetto: 3 trappole sono state poste all’interno del perimetro dell’area percorsa, priva di copertura arborea e con presenza di vegetazione erbacea spontanea (distanza compresa tra 0-300 m dal punto d’innesco); 3 trappole sono state collocate all’esterno del perimetro, in aree caratterizzate da un fitto querceto (Q. pubescens) e zone di macchia mediterranea alta (300-600 m).
La raccolta degli insetti caduti nelle trappole è stato effettuato ad intervalli di 7-10 giorni, nel periodo ottobre-dicembre 2009, per un totale di sette rilievi in campo. L’entomofauna raccolta è stata posta in appositi contenitori di plastica tramite un imbuto per polveri con imboccatura larga (2-3 cm di diametro). Nei contenitori di plastica è stato aggiunto alcool 70° per conservare il materiale raccolto fino al momento della classificazione.
Classificazione dell’entomofauna
La classificazione ha previsto alcune fasi preliminari di pulizia del materiale raccolto nelle trappole e separazione degli insetti da altri invertebrati e detriti di vario tipo. Successivamente si è passati ad una prima separazione dei taxa. L’operazione è stata condotta separando, per ciascun campionamento, prima gli individui di dimensioni maggiori e poi quelli più piccoli.
Per la classificazione dei taxa si è provveduto all’individuazione dell’ordine e della famiglia di appartenenza di ciascun esemplare raccolto mediante osservazione con microscopio ottico binoculare. In questo modo è stato possibile determinare la ricchezza di famiglie e l’abbondanza di individui in ciascuna famiglia di insetti per trappola e transetto.
Indici di biodiversità
Per quantificare la biodiversità entomatica sono stati determinati alcuni dei principali indici di biodiversità riportati in letteratura che tengono conto sia del numero che dell’abbondanza relativa di ciascun taxa ([11]): (1) Indice di Shannon-Weaver; (2) Indice di Uniformità o Equiripartizione di Pielou; (3) Indice di Simpson; (4) Indice di Margalef (Tab. 1).
Tab. 1 - Indici di diversità biologica utilizzati nello studio. pi : proporzione della i-esima specie (Σpi = 1); S: numero delle specie presenti; N: numero totale degli individui.
| Indice | Formula | Range |
|---|---|---|
| Shannon-Weaver | H’= - ∑ pi ln pi | 1.5 - 3.5 |
| Equiripartizione di Pielou |
E = H’/lnS | 0 - 1 |
| Indice di diversità Simpson | D = 1/ ∑ (pi )2 | 0 - 1 |
| Indice di Margalef | Dmg = (S-1)/ln N | ≥ 0 |
L’analisi degli effetti degli incendi in bosco ha previsto la costruzione di modelli di regressione lineare al fine di spiegare la variabilità del numero di individui per ciascun famiglia (abbondanza) e del numero di famiglie (ricchezza) di insetti in funzione della distanza dal punto d’innesco dell’incendio (DIST - variabile indipendente). In modo analogo si è proceduto per gli indici di biodiversità.
Risultati
Sono stati identificati 7 ordini e 21 famiglie di insetti, per un totale di 1077 individui (Tab. 2). La maggior parte degli esemplari ritrovati nelle trappole appartiene all’ordine dei Lepidotteri, cui seguono gli ordini dei Ditteri, Coleotteri, Imenotteri, Ortotteri, Emitteri, e un solo esemplare appartenente all’ordine Blattodea. In Tab. 3 si riportano i dati relativi all’abbondanza e la ricchezza di famiglie di insetti in relazione alle classi di distanza.
Tab. 2 - Risultati complessivi delle catture di insetti nel periodo di campionamento (ottobre-dicembre 2009).
| Ordini | Famiglie | Tot individui |
|---|---|---|
| Lepidoptera | Noctuidae | 667 |
| Nimphalidae | 9 | |
| Coleoptera | Carabidae | 79 |
| Staphylinidae | 59 | |
| Curculionidae | 3 | |
| Coccinellidae | 1 | |
| Crhysomelidae | 4 | |
| Anobidae | 2 | |
| Diptera | Muscidae | 76 |
| Tabanidae | 28 | |
| Gasterophilidae | 33 | |
| Cecidomyiidae | 39 | |
| Bibionidae | 3 | |
| Syrphidae | 2 | |
| Orthoptera | Tettigonidae | 6 |
| Gryllidae | 2 | |
| Hymenoptera | Vespidae | 54 |
| Formicidae | 5 | |
| Ichneumonidae | 2 | |
| Hemiptera | Pyrrochoridae | 2 |
| Blattodea | Blattidae | 1 |
| Totale | 1077 | |
Tab. 3 - Numero totale di famiglie e di individui raccolti per classe di distanza dal punto d’innesco dell’incendio.
| Distanza (m) | Totale famiglie | Totale individui |
|---|---|---|
| 100 | 12 | 169 |
| 200 | 11 | 151 |
| 300 | 8 | 180 |
| 400 | 15 | 257 |
| 500 | 14 | 154 |
| 600 | 14 | 166 |
| Totale | 74 | 1077 |
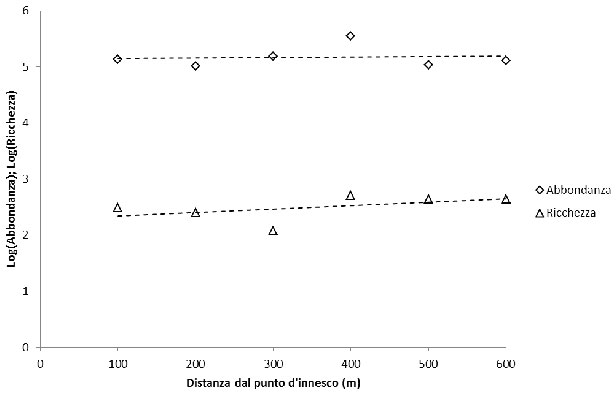
Dall’analisi dei modelli di regressione si evince l’esistenza di una relazione positiva crescente tra distanza dal fuoco (DIST) e ricchezza di famiglie di insetti. All’aumentare della distanza dal punto d’innesco dell’incendio aumenta la ricchezza di famiglie e quindi la diversità biologica a livello di entomofauna. Lo stesso andamento si riscontra considerando l’abbondanza di individui come variabile dipendente (Fig. 2). La capacità esplicativa dei modelli (R2) è, in entrambi i casi, pari a 0.24.
Fig. 2 - Relazione tra ricchezza di famiglie e abbondanza di individui (asse delle ascisse in forma logaritmica) e distanza dal punto d’innesco dell’incendio (m): Log (Ricchezza)= 2.279 0.001 · DIST (R2=0.24; p<0.05) Log (Abbondanza)=4.042 0.001 · DIST (R2=0.24; p<0.05).
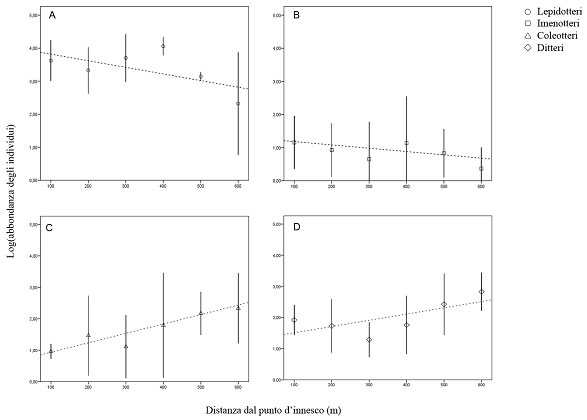
In Fig. 3 sono rappresentate le abbondanze relative ai principali taxa presi in considerazione. Si nota come nei Lepidotteri nottuidi e negli Imenotteri l’abbondanza tende a diminuire con l’aumentare della distanza dal punto d’innesco dell’incendio, al contrario di Ditteri e Coleotteri dove la polarità della relazione tra le due variabili risulta positiva per cui l’abbondanza di individui cresce con il crescere della distanza dal punto d’innesco dell’incendio.
Fig. 3 - Relazione tra abbondanza di individui e distanza dal punto d’innesco dell’incendio (m). (A) Lepidotteri = 0.392 - 0.002 · DIST (R2=0.33; p<0.05); (B) Imenotteri: Log(Abbondanza) = 1.283 - 0.002 · DIST (R2=0.26; p<0.05); (C) Coleotteri: Log(Abbondanza) = 0.981 0.003 · DIST (R2=0.77; p<0.05); (D) Ditteri: Log(Abbondanza) = 1.532 0.002 · DIST (R2=0.51; p<0.05)
Al fine di comprendere la diversa risposta degli insetti nei diversi taxa è stato determinato il numero di individui raccolti in due classi di distanza dal punto d’innesco: classe 1= 0-300 m; classe 2 = 300-600 m.
L’analisi della varianza (ANOVA) mostra differenza significative (p<0.05) nel caso dei Coleotteri la cui media di individui raccolti nelle due classi di distanza sono rispettivamente 4.33 e 12.11. Per gli altri taxa le differenze di abbondanza di individui nelle due classi di distanza non sono significative (p>0.05). Tali risultati evidenziano come i Coleotteri terricoli (Carabidi e Stafilinidi), sono influenzati in modo negativo dall’incendio soprattutto in relazione al mutamento dell’habitat originario.
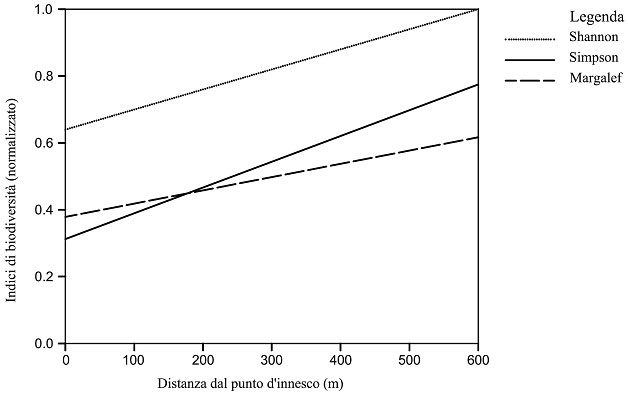
In Tab. 4 si riportano i valori degli indici di biodiversità in relazione alla distanza dal punto d’innesco dell’incendio (DIST). Dalla analisi di questi dati è stato possibile costruire dei modelli di regressione al fine di comprendere la differente risposta di questi indici sintetici in funzione della variabile DIST (Fig. 4). A questo riguardo, l’indice di Simpson ha mostrato un coefficiente angolare maggiore rispetto agli altri due e questo può essere interpretato come una maggiore attitudine di questo indice a monitorare gli effetti nel breve periodo degli incendi sulla biodiversità dell’entomofauna.
Tab. 4 - Valori degli indici di biodiversità presi in considerazione in questo studio in relazione alla distanza dal punto d’innesco dell’incendio.
| Dist. (m) |
Shannon- Weaver |
Uniformità di Pielou |
Simpson 1-D |
Margalef |
|---|---|---|---|---|
| 100 | 1.10 | 0.45 | 0.45 | 2.14 |
| 200 | 1.33 | 0.55 | 0.55 | 1.99 |
| 300 | 0.80 | 0.4 | 0.33 | 1.34 |
| 400 | 1.24 | 0.45 | 0.50 | 2.52 |
| 500 | 1.43 | 0.55 | 0.77 | 2.58 |
| 600 | 1.92 | 0.7 | 0.80 | 2.54 |
Fig. 4 - Relazione tra gli indici di diversità biologica e distanza dal punto d’innesco dell’incendio (m). Shannon = 0.656 0.005 · DIST (R2=0.48; p<0.05); Simpson = 0.317 0.007 · DIST (R2=0.51; p<0.05); Margalef (normalizzato) = 0.386 0.004 · DIST (R2=0.31; p<0.05)
Discussione
La biodiversitàè un tema centrale in ecologia forestale, specie se considerata in termini di influenza sulla stabilità delle popolazioni in risposta ad eventi perturbativi interni e/o esterni al sistema ([4], [9], [26]).
Dall’analisi dei risultati di questo studio emerge una relazione significativa tra distanza dal punto d’innesco dell’incendio e biodiversità, intesa come ricchezza di famiglie di insetti o sotto forma di indice sintetico. Gli incendi possono ridurre notevolmente la complessità strutturale di un bosco: (1) condizionando il libero movimento degli animali all’interno del territorio; (2) alterandone in modo a volte irreversibile i loro cicli fisiologici e riproduttivi; (3) influenzando negativamente l’abbondanza e la ricchezza dell’entomofauna, soprattutto terricola, che avendo bassa mobilità, soffre maggiormente questi fenomeni di disturbo ([9]).
Ciò spiega in parte l’andamento delle regressioni lineari positive riferite alla ricchezza di famiglie e al numero di individui (Fig. 2). Nel caso dell’abbondanza di individui, ad esempio, la retta di regressione cresce lievemente all’aumentare della distanza dall’area incendiata a seguito del diverso comportamento riscontrato per i singoli ordini campionati (ampia varianza: R2 = 0.24). L’andamento delle rette di regressione in Fig. 3, conferma la netta differenza di comportamento tra i taxa osservati: i Coleotteri, così come i Ditteri, aumentano di numero, passando dalla trappola numero 1 alla trappola numero 6, al contrario dei Lepidotteri e degli Imenotteri che tendono a diminuire.
Nel caso dei Coleotteri Carabidi l’andamento del modello è in linea con lo studio effettuato da Saint-Germain et al. ([23]), i quali hanno riscontrato in un’area incendiata di 250 km2 una bassa abbondanza di Coleotteri Carabidi e l’assenza di specie tipiche post-incendio per i successivi due anni. I risultati di questo studio sono in accordo anche con Niemelä et al. ([17]), secondo i quali è necessario molto tempo affinché specie di Carabidi possano stabilire popolazioni numerose e colonizzare spazi aperti come quelli che si vengono a formare dopo un incendio.
In uno studio comparativo di Paquin & Coderre ([18]), si è constatato che la deforestazione riduce l’abbondanza di artropodi terricoli in media del 55%, rispetto al 95.5% registrato in conseguenza di eventi perturbanti come l’incendio.
Alcuni taxa di insetti (ad es., Lepidotteti Nottuidi) si avvalgono di organi sensoriali per individuare fumo e calore. Questi segnali vengono utilizzati per identificare e convergere nei nuovi habitat che si vengono a creare dopo l’incendio. Questo processo aumenta la competitività e l’antagonismo nei confronti delle specie rimaste nell’area percorsa e spiega parzialmente la differenza nel numero di individui nelle popolazioni di Carabidi e Lepidotteri.
Gandhi et al. ([5]) hanno analizzato l’abbondanza e la ricchezza di specie di Carabidi e Stafilinidi in aree residuali post-incendio nelle foreste di conifere in Alberta, Canada. Il risultato dello studio dimostra la relazione tra grosse popolazioni di Carabidi e le suddette aree residuali. In quest’ultime, sono spesso presenti aree caratterizzate da elevata necromassa e biomassa in grado di fornire habitat per Carbidi e Stafilinidi. La conservazione di queste patch all’interno di un ecosistema forestale percorso dal fuoco è essenziale al fine di mantenere intatte le popolazioni di Coleotteri e incrementare la complessità del paesaggio.
Questo studio è coerente con i risultati riportati nella letteratura di settore, poiché il graduale aumento della popolazione avviene parallelamente al cambiamento delle condizioni della vegetazione e del soprassuolo che diventano sempre più idonee ai Coleotteri. Alberi sani e rigogliosi, vegetazione di sottobosco, forniscono l’habitat ideale per il nutrimento, e offrono una naturale difesa contro i predatori. Nel caso dei Lepidotteri l’andamento del modello dimostra che la sostanziale diminuzione del numero di esemplari man mano che ci si allontana dal punto d’innesco dell’incendio, in accordo con numerosi studi effettuati in passato ([20]). I risultati di questo studio sono in linea con il lavoro di Huntzinger ([7]) che ha testato la risposta di Lepidotteri all’uso della pratica degli incendi controllati nel Yosemite National Park e Rogue River National Forest (USA).
Grundel et al. ([6]) hanno riscontrato come l’eterogeneità degli incendi (stagionalità, tipologia, severità ) possa essere necessaria al mantenimento di una complessa struttura del soprassuolo che assicuri ai Lepidotteri accoppiamento, ovideposizione e nutrimento. Questo perché molte specie di Lepidotteri necessitano di habitat differenti per completare il loro ciclo di vita. Inoltre non è da trascurare che l’aumento di Lepidotteri incrementa la disponibilità di prede animali a favore non solo di invertebrati entomofagi, ma anche di uccelli e mammiferi insettivori. Ciò riveste un particolare interesse per quanto riguarda la qualità degli habitat di specie a rischio, individuate nell’ambito delle reti di protezione della natura ([13]).
In questo studio i Lepidotteri non hanno risentito del passaggio del fuoco poiché in grado di spostarsi da una patch boschiva ad una altra in cerca di nutrimento e riparo. I Lepidotteri abbondano in aree aperte dove la bassa vegetazione viene raggiunta con più facilità in assenza di copertura arborea. Inoltre, in questi contesti la radiazione luminosa consente lo sviluppo di una densa vegetazione arbustiva e procura corridoi per il movimento e la dispersione dei Lepidotteri tra patch ([2]).
Dai risultati si evince anche una presenza numerosa di Imenotteri e Ditteri, i quali pur non essendo dei veri e propri indicatori ecologici, svolgono una funzione molto importante nell’ecosistema in qualità di insetti impollinatori. Tra gli Imenotteri trovati, la famiglia più numerosa è stata quella dei Vespidi, che per il nutrimento e il nido dipendono molto dall’habitat originario, caratterizzato da ambienti aperti e assolati contraddistinti dalla presenza di una flora diversificata; inoltre risulta indispensabile l’esistenza di condizioni favorevoli alla nidificazione ([19], [8]). Queste preferenze di habitat sono in accordo con quanto analizzato in questo lavoro e spiegano la relazione negativa del modello “abbondanza-distanza” per gli Imenotteri.
Quest’analisi è in linea anche con il lavoro di Moretti et al. ([14]) i quali hanno riscontrato un effetto positivo del fuoco sugli insetti impollinatori che approfittano della riduzione della copertura arborea e della presenza di una folta vegetazione transitoria nella prima fase post-incendio.
Per quanto riguarda i Ditteri (Fig. 3), il modello di regressione mostra un effetto negativo del fuoco che determina abbassamento della popolazione nelle vicinanze al punto d’innesco dell’incendio. Risulta difficile dare una spiegazione chiara del motivo di tale andamento del modello poiché le famiglie campionate hanno caratteristiche ecologiche e esigenze trofiche piuttosto eterogenee.
Analizzando la famiglia più rappresentativa tra quelle campionate, ossia i Cecidomidi, possiamo dire che la maggior parte delle specie, ascrivibili a tale famiglia, sono prevalentemente fitofaghe e si nutrono a spese di piante superiori. Fra i fitofagi sono comprese anche numerose specie galligene, che producono vere e proprie galle oppure deformazioni degli organi dette “pseudogalle”; altre specie vivono come commensali in galle prodotte da altri insetti, talvolta come coprofaghe, saprofaghe o micetofaghe ([12]). La presenza di un denso bosco all’esterno dell’area incendiata e la presenza di galle sui rami di Roverella potrebbe chiarire l’andamento del modello riferito ai Ditteri.
La poca certezza è data da evidenze scientifiche in contrasto tra loro: Ne’eman et al. ([16]) hanno osservato un effetto negativo di incendi estivi su insetti impollinatori, tra cui anche i Ditteri, mentre Moretti et al. ([15]) hanno verificato un effetto positivo degli incendi invernali sulle Alpi Svizzere nei confronti di Ditteri Sirfidi.
Conclusioni
I risultati di questo studio consentono di affermare che fenomeni perturbanti quali gli incendi in aree boschive mediterranee determinano una generale riduzione della biodiversità entomatica.
Dal punto di vista della abbondanza di individui e ricchezza di famiglie, l’andamento dei modelli di regressione è di tipo lineare positivo ossia la biodiversità entomatica aumenta all’aumentare della distanza dal punto di innesco dell’incendio. Considerando la risposta degli insetti nei diversi taxa, i risultati di questo studio evidenziano come i Coleotteri terricoli (Carabidi e Stafilinidi), sono influenzati in modo negativo dall’incendio soprattutto in relazione al mutamento dell’habitat originario. Tuttavia occorre considerare che nel caso dei Lepidotteri e degli Imenotteri la relazione distanza e abbondanza di individui è di tipo lineare negativo: il fuoco e gli habitat che si vengono a creare dopo l’incendio sono elementi attrattivi per Lepidotteri (Nottuidi) e per gli Imenotteri.
I risultati di questo studio, seppur limitati dal breve periodo di campionamento dell’entomofauna, lasciano intendere una influenza negativa tra incendi in bosco e biodiversità entomatica ad un anno dall’evento. L’azione dell’incendio riduce nel breve periodo il numero e l’abbondanza di famiglie di insetti con conseguenze negative su tutta una serie di processi ecologici legati alla presenza di questi taxa ([4]).
Ringraziamenti
Si ringrazia il Prof. Vittorio Leone per i suggerimenti forniti durante lo studio e per la revisione critica.
References
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar