Extra-zonal beech forests in Tuscany: structure, diversity and synecologic features
Forest@ - Journal of Silviculture and Forest Ecology, Volume 8, Pages 88-102 (2011)
doi: https://doi.org/10.3832/efor0659-008
Published: Jul 19, 2011 - Copyright © 2011 SISEF
Research Articles
Abstract
The present paper focuses on the structural, synecological and floristic diversity features of beech-dominated forest communities in four major areas of the Antiapenninic Tyrrhenian system in Tuscany: Metalliferous hills, mountains to the south of Mt. Amiata, volcanic area of the upper Lente valley and Mt. Cetona. These are relict woodlands of Holo-Pleistocene origin with a special ecological and conservation value due to their extrazonal location in lowland submediterranean areas. Results show substantial among-area differences in structure, synecology and plant species composition, but in general a potential for coppices to reach the tall forest stage, as demonstrated by the old-growth stands of Pietraporciana and Sassoforte. Compared with montane Apenninic beechwoods, the relatively rich flora of the studied communities include thermophilous species with a southern Apennine-Balkan distribution, making their syntaxonomical position unclear. Closer affinities are found with the calcicolous Beech Forests of the association and with the silicicolous ones of the . Based on the Natura 2000 system, all the examined communities belong to the priority Habitat “Apennine beech forests with and ” (code: 9210*). Due their relict nature, these biotopes appear vulnerable to climate changes and to a production-oriented forest management. Criteria of naturalistic silviculture should instead promote the dynamic development of these communities towards tall forests and their natural regeneration.
Keywords
Beech forest, Extrazonal forests, Phytosociology, Plant diversity, Southern Tuscany, Forest structure, Synecology
Introduzione
Col termine di popolamento extrazonale o eterotopico si designano quei nuclei di vegetazione zonale di origine naturale posti al di fuori del contesto macroclimatico tipico, e quindi della regione biogeografica di normale appartenenza. Generalmente tale fenomeno è ascrivibile all’azione combinata di cause storiche ed ecologiche. È infatti spesso legato a vicende paleoclimatiche e paleogeografiche l’insediamento di determinati popolamenti forestali in un dato territorio, mentre la loro conservazione in alcune parti di esso al mutare delle condizioni generali è da attribuirsi alle caratteristiche microclimatiche ed edafiche particolari rispetto alle zone circostanti ([41], [2], [3], [44], [48]). Ne risultano degli ecosistemi boschivi spesso rari e di limitata estensione, che tuttavia contribuiscono notevolmente alla diversità bioecologica della regione in cui sono insediati a livello di specie ed habitat.
In Italia, ed in Toscana in particolare, esistono diversi esempi di popolamenti forestali extrazonali, quali, ad es., i nuclei relitti di Picea abies della Riserva naturale di Campolino nell’Appennino tosco-emiliano ([36]), o quelli di Betula pendula e Taxus baccata della Val di Farma in provincia di Siena e Grosseto ([21], [40]).
Senza dubbio, una delle specie forestali maggiormente presenti sul territorio con popolamenti di natura eterotopica è il faggio (Fagus sylvatica L.). Specialmente nei vasti sistemi collinari e basso-montani dell’Antiappennino tirrenico esistono lembi di faggeta posti al di sotto dei 1000 m s.l.m., limite inferiore dell’ottimo ecologico per la specie lungo la catena Appenninica principale ([8], [33], [5]). Esiste una sostanziale concordanza sul fatto che l’origine di questi popolamenti risalga ad epoche recenti dell’Olocene, quando l’elevata oceanicità del clima del periodo Subatlantico può avere spinto la specie ad avvicinarsi alla costa tirrenica tosco-laziale scendendo notevolmente di altitudine ([41], [42], [39], [2], [4], [19], [47]). Date le mutate condizioni climatiche ed anche il secolare sfruttamento da parte dell’uomo, questi popolamenti hanno oggi carattere relitto. L’impatto antropico ed il contesto climatico submediterraneo in cui si trovano pongono chiari limiti alla loro capacità di tolleranza verso stress idrici ed alte temperature, rendendoli vulnerabili ad un dinamismo che ne potrebbe determinare la sostituzione con formazioni più termoxerofile o con nuclei di specie alloctone invasive.
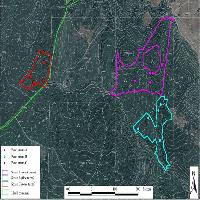
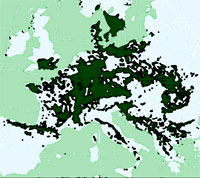
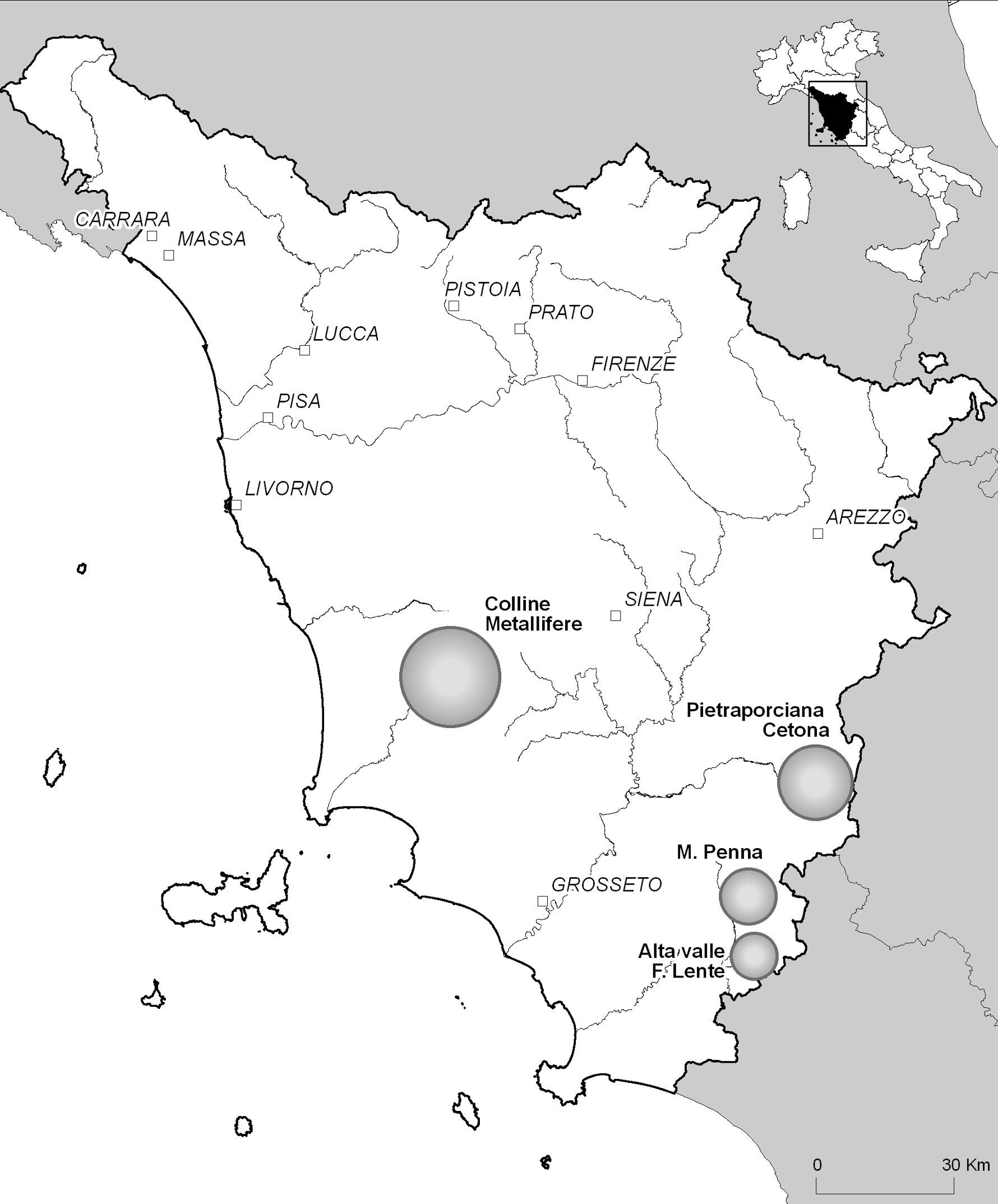
Le faggete extrazonali si possono inquadrare in due tipologie differenziate, le abissali e le cacuminali. Le prime sono accantonate solitamente al di sotto dei 500 m s.l.m., in valloni in prossimità di corsi d’acqua con marcati fenomeni di inversione termica ([2], [3], [48], [51]), mentre le seconde occupano dorsali di gruppi montuosi isolati generalmente a quote inferiori ai 1000 m s.l.m. Entrambe queste tipologie sono presenti nell’Antiappennino tirrenico, più specificamente nei comprensori delle Colline Metallifere, dei rilievi a sud del Monte Amiata, dei tufi volsini dell’Alta Valle del Fiora e del Monte Cetona (Fig. 1).
Fig. 1 - Localizzazione dei comprensori analizzati. Le sfere coprono la superficie del comprensorio al cui interno sono localizzati i rilevamenti effettuati.
L’interesse per tali biotopi è testimoniato da diversi contributi che descrivono popolamenti locali da vari punti di vista; oltre a quelli già ricordati si citano Cavalli & Drosera ([16]), Frassineti & Bottacci ([28]), Scoppola & Caporali ([48]), Viciani et al. ([51]), Angiolini et al. ([1]).
Tuttavia, molte situazioni risultano ancora poco conosciute, e soprattutto manca una visione d’insieme delle loro caratteristiche sinecologiche e della loro possibile posizione sintassonomica in rapporto alle unità attualmente riconosciute per le faggete italiane. Tali aspetti assumono un certo interesse anche in considerazione della natura di transizione fitogeografica della Toscana meridionale, territorio al limite tra la zona europea e quella mediterranea ([4], [44]). Di conseguenza questi boschi potrebbero presentare affinità sia con le faggete zonali di tipo centroeuropeo sia con quelle a carattere mediterraneo-montano che, secondo alcuni autori, risalgono frammentariamente a questa latitudine sul versante tirrenico ([44]). Anche nel recente “Manuale di interpretazione degli habitat italiani” ([10]) i riferimenti sintassonomici rimangono poco definiti, anche a causa della mancanza di dati e della loro limitata estensione.
Il presente studio intende contribuire ad una maggiore conoscenza di questi particolari ambienti forestali, analizzandone le principali caratteristiche strutturali, fitosociologiche e di diversità floristica, e offrendo spunti utili per una loro gestione sostanzialmente conservativa.
Materiali e metodi
Localizzazione delle aree di studio e descrizione del territorio
Sulla base dei dati bibliografici a disposizione ([18], [42], [28], [6], [51], [1]), e di indagini sul campo preliminari, sono stati individuati i popolamenti extrazonali di faggio più cospicui della Toscana meridionale. Le aree campionate sono ripartite tra le seguenti quattro aree principali: Colline Metallifere, rilievi a Sud del M. Amiata (M. Penna), alto bacino del Fiora (Valle del Lente) e comprensorio del M. Cetona (Fig. 1).
Dal punto di vista geologico il territorio è caratterizzato dalla cosiddetta serie ridotta, in cui la Falda toscana non è presente in maniera continua; da ciò deriva una certa varietà nella pedologia ([35], [22], [23], [31], [15]). Le faggete analizzate si trovano su substrati derivati da tre diversi tipi litologici fondamentali: calcarei, arenacei (verrucano e arenaria), vulcanici (tufi e rioliti - vedi Appendice 1), con importanti influenze sulla vegetazione presente. Per quanto riguarda la pedologia, la presenza delle faggete ha contribuito in tutti i casi alla formazione di terre brune, più o meno profonde ed evolute a seconda della litologia di base. Si segnalano in particolare le faggete della Valle del Lente, dove le vulcaniti presenti hanno permesso la formazione di andosuoli, terre brune molto profonde e fertili, caratterizzate dalla presenza di materiali vetrosi amorfi (allofane).
All’interno delle aree di studio non esistono stazioni termo-pluviometriche, quindi i principali parametri climatici sono stati dedotti in base ai dati raccolti nelle stazioni più vicine ([9]), e riportati sinteticamente in Tab. 1. In generale tutte le stazioni sono caratterizzate da precipitazioni piuttosto elevate (attorno a 1000 mm) e ben distribuite durante l’anno. Le temperature medie invernali sono relativamente miti grazie alla ridotta distanza dal mare, mentre la presenza della principale catena appenninica a Nord ripara il territorio dai venti freddi settentrionali. I dati rivelano quindi nel complesso condizioni climatiche di tipo suboceanico ([12], [7]); tale oceanicità risulta probabilmente ancora più marcata nelle aree di studio, essendo queste collocate ad altitudini superiori. In Tab. 1 sono riportati sia le formule climatiche secondo Thornthwaite & Mather ([49]), che i principali parametri di interesse bioclimatico, ricavati dall’elaborazione dei dati climatici secondo l’approccio di Rivas-Martinez. L’analisi conferma che il principale bioclima di riferimento è il Temperato Oceanico, e la fascia bioclimatica quella Supratemperata Umida. Il microclima delle stazioni dominate dal faggio è caratterizzato da condizioni locali di elevata umidità atmosferica ed edafica, con frequenti nebbie mattutine, dovute a fenomeni di inversione termica, e all’effetto dell’esposizione o dell’altitudine. Tutta l’area è favorevolmente esposta ai venti umidi sud-occidentali, carichi di umidità, che grazie alla presenza dei rilievi contribuiscono a determinare l’effetto STAU, cioè la risalita di venti umidi a causa del “blocco” esercitato dall’orografia (convezione forzata), e il conseguente innesco di fenomeni precipitativi per condensazione del vapore acqueo trasportato, che può essere notevole data la relativa vicinanza al mare ([7]).
Tab. 1 - Formule climatiche delle stazioni termo-pluviometriche più prossime alle aree di studio, secondo Thornthwaite & Mather ([49]), calcolate con i dati di Bigi & Rustici ([9]). Con gli stessi dati sono indicate le caratteristiche bioclimatiche secondo l’approccio di Rivas-Martinez (⇒ http://www.globalbioclimatics.org). (T): Temperatura; (P): precipitazioni.
| Stazione termo-pluviometrica (area di riferimento) | Quota stazione (m s.l.m.) | Quote aree di riferimento (m s.l.m.) | T media annua (°C) | P medie annue (mm) | Formula climatica | Bioclima | Fascia bioclimatica |
|---|---|---|---|---|---|---|---|
| Chiusdino(Colline Metallifere) | 564 | 450-1000 | 13.7 | 956 | B1 B’2 s b’4 | Mediterranean Pluvioseasonal-Oceanic | Upper Mesomediterranean Upper Subhumid |
| San Casciano dei Bagni (M. Cetona) | 582 | 800-1100 | 12.4 | 1021 | B2 B’2 r b’4 | Temperate Oceanic (Submediterranean) | Low Supratemperate Low Humid |
| Piancastagnaio(rilievi a Sud del M. Amiata) | 772 | 770-880 | 11.4 | 1270 | B4 B’1 r b’4 | Temperate Oceanic (Submediterranean) | Low Supratemperate Upper Humid |
| Pitigliano(Alto Bacino del Fiora) | 313 | 420-450 | 13.9 | 918 | B1 B’2 s b’4 | Temperate Oceanic (Submediterranean) | Low Supratemperate Upper Subhumid |
Analisi della struttura
Nelle aree indagate sono stati eseguiti 24 rilevamenti strutturali in fitocenosi omogenee su aree di saggio rettangolari di 20 x 10 m (200 m2), scelte in maniera soggettiva ma randomizzata all’interno delle aree dominate dal faggio: dopo un sopralluogo iniziale di ogni popolamento di interesse si sono stabilite tali aree in zone rappresentative delle varie condizioni del popolamento stesso, aumentandone il numero laddove maggiori fossero l’eterogeneità e/o l’estensione. Per consentire lo svolgimento anche del rilevamento fitosociologico nella stessa area, la dimensione è stata scelta secondo i principi del metodo Braun Blanquet ([13], [14]). I dati stazionali e di localizzazione geografica, rilevati con GPS satellitare, sono riportati nell’Appendice 2. In ciascuna area di saggio sono state rilevate la densità e la struttura orizzontale degli alberi di faggio, fortemente dipendenti dal tipo di gestione del bosco attuale e pregresso. Servendosi di cavalletto dendrometrico e rotella metrica, si è eseguito un cavallettamento totale al centimetro coperto dei fusti di diametro a petto d’uomo superiore a 5 cm, durante il quale le matricine, le ceppaie e i singoli polloni sono stati numerati e contati, prestando attenzione ad attribuire ogni pollone alla ceppaia da cui si è originato. Una volta riuniti i diametri misurati in classi ampie 5 centimetri, è stato possibile il calcolo dei seguenti parametri: area basimetrica, numero di ceppaie e di fusti (sia di origine gamica che agamica) ad ettaro e diametro medio.
Analisi fitosociologia e sinecologica
Nelle stesse fitocenosi analizzate dal punto di vista strutturale sono stati eseguiti 24 rilevamenti fitosociologici col metodo di Braun Blanquet ([13], [14]). Dopo le necessarie verifiche floristiche di laboratorio, è stato compilato un elenco floristico basato su Pignatti ([43]), per nomenclatura, forma biologica e corotipo, rendendo possibile l’elaborazione degli spettri biologico e corologico. I rilievi sono stati riuniti in una tabella riassuntiva, in seguito sottoposta ad analisi esplorative tramite l’impiego di pacchetti statistici. È stata effettuata una Cluster Analysis (CA) su una matrice specie x rilievi, con i valori di abbondanza-dominanza modificati in accordo con Van Der Maarel ([50]), usando come indice di similarità la distanza della corda e come algoritmo il legame medio (UPGMA), con i quali si è ottenuto il raggruppamento migliore e il più elevato coefficiente di correlazione cofenetica (> 0.8). La diversità floristica di ogni rilevamento è stata stimata attraverso l’indice di Shannon (H’) sulla base del grado di copertura percentuale di ogni singola specie rispetto al totale ([34]). Un’analisi ecologica di base è stata effettuata utilizzando gli indici di Ellenberg rielaborati da Pignatti ([45]), ponderati per ogni rilevamento in base alla copertura della specie; la risultante matrice di varianza-covarianza dei valori medi di ogni singolo fattore per ciascun rilevamento è stata sottoposta ad analisi multivariata tramite Principal Component Analysis (PCA) con impiego del software PAST ([32]).
In linea con un recente studio sulle formazioni forestali mesofile del Casentino ([46]), un’ulteriore analisi è stata condotta per esaminare il rapporto fra densità del faggio, in termini di area basimetrica, e diversità floristica, per mezzo del coefficiente di correlazione per ranghi di Spearman.
Risultati
Struttura dei popolamenti
La Tab. 2 riassume i principali parametri dendrometrici rilevati in ogni area, mediati per i vari comprensori, mentre la distribuzione delle frequenze diametriche del faggio è mostrata in Fig. 2, ottenuta sommando i dati derivanti da tutte le aree di saggio effettuate in ogni comprensorio. L’area basimetrica media del faggio è risultata di 48.7 m2/ha. Esistono tuttavia sostanziali variazioni in termini di densità e copertura sia all’interno dei singoli comprensori sia fra gli stessi. Nella maggior parte dei casi si tratta di cedui a sterzo invecchiati, in cui le utilizzazioni sono sospese da tempi più o meno lunghi (soprassuoli transitori in conversione all’alto fusto per abbandono delle pratiche selvicolturali); in due casi invece i boschi hanno mostrato una struttura simile a quella tipica della fustaia.
Tab. 2 - Valori medi dei principali parametri dendrometrici in ogni comprensorio. (G/ha): area basimetrica ad ettaro.
| Valori medi Comprensorio | Diametro medio [cm] (CV%) | Ceppaie/ha (CV%) | Piante/ha (CV%) | Polloni/ceppaia (CV%) | G/ha [m2/ha] (CV%) |
|---|---|---|---|---|---|
| Monte Penna | 23.3 (19.2%) | 738.0 (50%) | 1438.0 (64.4%) | 1.8 (32.4%) | 50.5 (39.2%) |
| Colline Metallifere | 23.1 (30.6%) | 715.8 (48.9%) | 1560.1 (67.7%) | 2.2 (39.4%) | 46.9 (39.2%) |
| Valle Lente | 15.3 (27.3%) | 1017.3 (41.1%) | 1728.7 (41.8%) | 1.7 (17.6%) | 38.2 (8.3%) |
| Cetona | 19.7 (56.7%) | 970.7 (31.0%) | 2639.8 (88.0%) | 2.4 (64.6%) | 56.7 (57.6%) |
| Totale | 21.5 (34.5%) | 812.1 (43.5%) | 1775.6 (76.0%) | 2.1 (45.2%) | 48.7 (42.7%) |
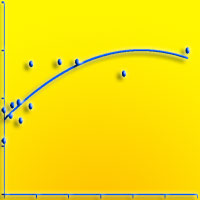
Fig. 2 - Distribuzione delle frequenze diametriche rilevate nelle faggete dei quattro comprensori. L’andamento della curva fornisce indicazioni circa la struttura dei popolamenti analizzati. (A): Monte Penna; (B): Colline Metallifere; (C): Valle del Lente; (D): Cetona.
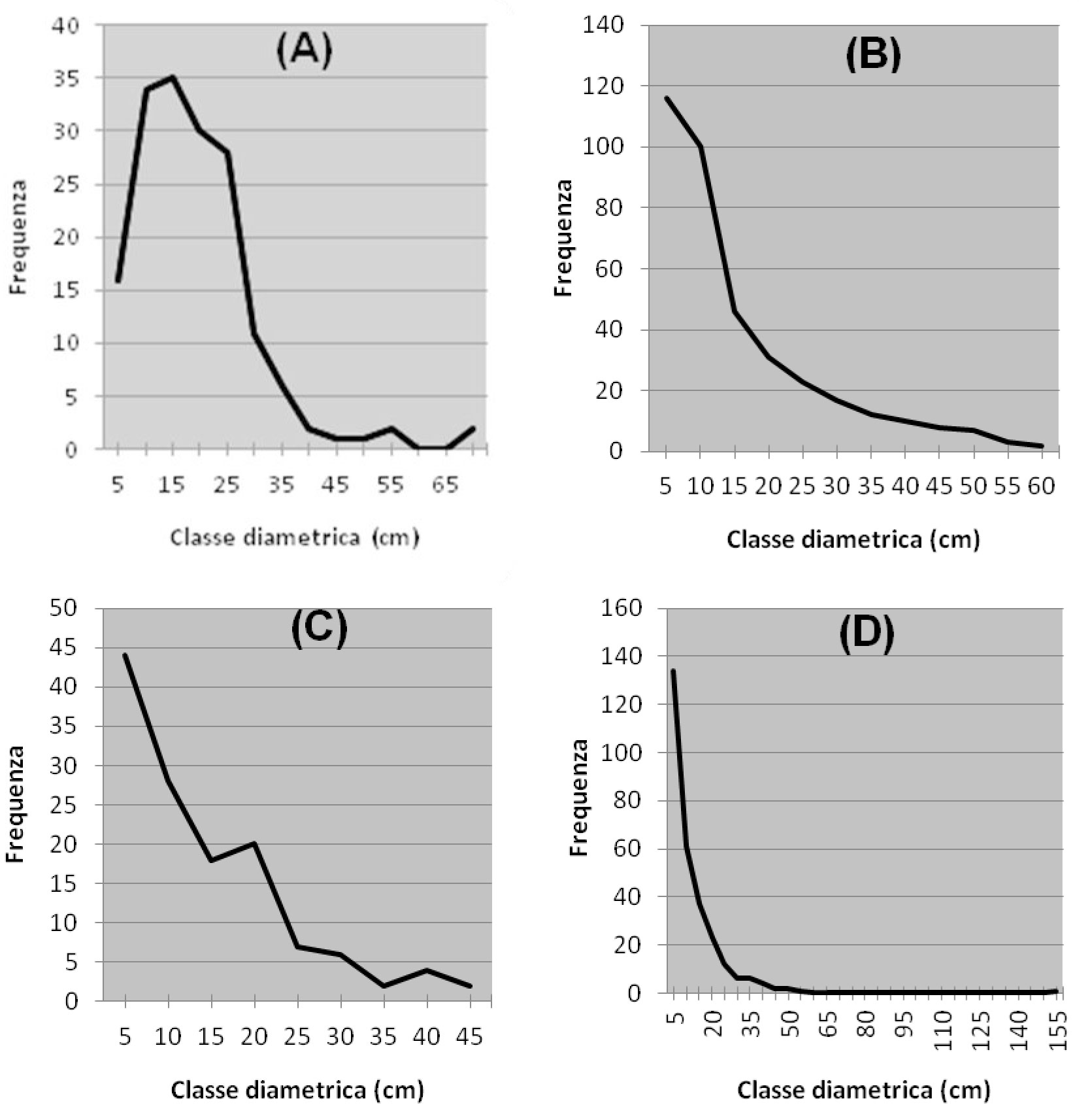
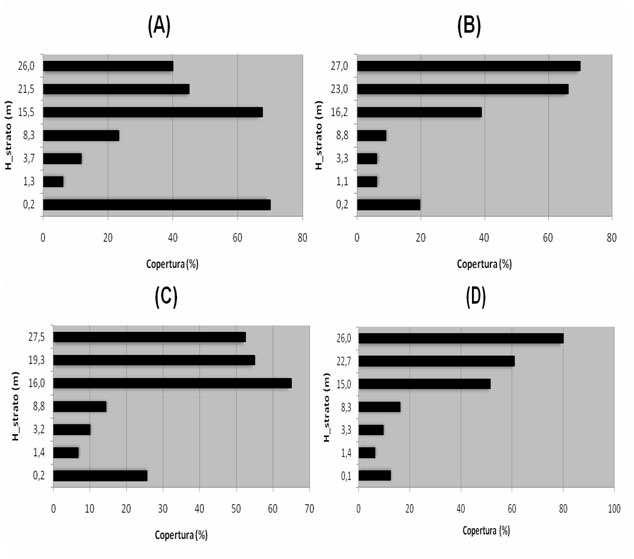
Nella zona del Monte Penna si trovano cedui abbandonati, con densità mediamente elevate (> 50 m2/ha) e dimensioni delle piante abbastanza sostenute (c. 23 cm). Esiste una certa variabilità all’interno del comprensorio ed in alcuni tratti la struttura si avvicina a quella della fustaia a causa del lungo tempo di invecchiamento. La curva delle frequenze diametriche evidenzia la presenza di soggetti di notevoli dimensioni, e illustra come la competizione tra i polloni del vecchio ceduo stia favorendo le classi intermedie (15-25 cm). La struttura orizzontale è caratterizzata da una copertura elevata sia degli strati dominanti e codominanti (> 50%), che dello strato erbaceo. Ridotta è invece la percentuale di copertura degli strati intermedi e arbustivi.
Nell’area più settentrionale delle Colline Metallifere sono state rilevate diverse faggete non in continuità una con l’altra e su diversi substrati geomineralogici, determinando quindi un’ulteriore causa di variabilità dimensionale dei soggetti arborei. Mediamente si ha una densità leggermente inferiore al caso precedente. Si tratta ancora di cedui invecchiati, ma con strutture meno evolute verso l’alto fusto. La distribuzione delle frequenze dei diametri mostra in questo caso il netto predominio delle classi inferiori sulle altre. Un’importante eccezione è data dalla faggeta relitta sul ripido versante settentrionale del M. Sassoforte, che presenta la struttura tipica di un bosco di alto fusto di notevole sviluppo verticale (> 20 m) ed individui di diametro medio di 32 cm. Questo, unitamente alla presenza di discrete quantità di necromassa legnosa in decomposizione, conferisce al popolamento l’aspetto di bosco vetusto. Sul Fosso del Bardellone e nel Poggio di Montieri si ha una situazione di elevata densità, ma l’evoluzione del ceduo verso la fustaia è meno avanzata, soprattutto nel primo caso. In una singola località, Fontalcinaldo, è presente una condizione anomala, in cui il faggio compare solo con pochi esemplari (10% in numero) di grosse dimensioni, i soli rilasciati durante recenti utilizzazioni molto intense.
Nella zona della valle del Lente, la più meridionale tra quelle analizzate, le faggete presentano una densità minore (38.2 m2/ha), e dimensioni medie degli alberi inferiori. La struttura è quella di cedui invecchiati in uno stadio avanzato di evoluzione verso la fustaia. La variabilitàè generalmente inferiore agli altri casi. L’andamento delle frequenze diametriche è simile al caso precedente. La struttura orizzontale è piuttosto diversa dagli altri casi: la copertura maggiore per le specie arboree è rappresentata dallo strato codominante, mentre sotto il 50% risultano i due strati superiori, che comunque raggiungono altezze ragguardevoli; se gli strati arbustivi sono comunque piuttosto contenuti, la situazione avvantaggia nettamente le specie erbacee, presenti con la copertura più elevata in assoluto (70%).
L’area del Cetona presenta complessivamente una situazione intermedia per quanto riguarda le dimensioni diametriche, mentre molto elevata risulta la densità, con i massimi valori di area basimetrica riscontrati (56.7 m2/ha). I coefficienti di variazione sono spesso molto elevati, poiché tale comprensorio riunisce due situazioni estreme: la faggeta secolare di Pietraporciana e la faggeta del Monte Cetona, in cui le utilizzazioni del vecchio ceduo sembrano sospese da un minor numero di anni, e pertanto la struttura risulta meno evoluta (molti polloni per ceppaia e a ettaro). Nel complesso la distribuzione delle frequenze mostra ancora una volta un elevato numero di soggetti di diametro inferiore ai 20 cm. La struttura orizzontale di queste faggete presenta valori di copertura mediamente elevati per gli strati dominanti e codominanti, bassi per quelli dominati e arbustivi, intermedi per lo strato erbaceo.
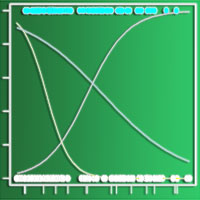
La stratificazione verticale mediata dei popolamenti dei quattro comprensori, derivata dal rilevamento fitosociologico condotto per strati separati, è rappresentata in Fig. 3.
Fig. 3 - Struttura verticale ottenuta considerando la percentuale media di copertura dei vari strati della vegetazione, indicati tramite la loro altezza media nell’asse delle ordinate. (A): Monte Penna; (B): Colline Metallifere; (C): Valle del Lente; (D): Cetona.
Rilevamento fitosociologico
I 24 rilievi fitosociologici mostrano un grado di eterotonia piuttosto elevato (indice di Klement = 6.15), a causa del considerevole numero di specie con frequenza < 40%. Le più diffuse, oltre al faggio, sono risultate Hedera helix, Melica uniflora, Anemone nemorosa, Mercurialis perennis, Ostrya carpinifolia e Ilex aquifolium. I rilevamenti sono stati ordinati nell’Appendice 3 secondo la sequenza del dendrogramma restituito dalla cluster analysis (Fig. 4). Tale grafico mostra la posizione isolata del rilevamento eseguito in località Fontalcinaldo (area Colline Metallifere), a causa di un contingente floristico fortemente alterato da recenti utilizzazioni forestali che hanno provocato la trasformazione del precedente soprassuolo transitorio di faggio in un ceduo misto di specie fortemente pollonifere, quali Fraxinus ornus e Ostrya carpinifolia.
Fig. 4 - Dendrogramma da Cluster Analysis basata sull’abbondanza delle specie nei rilevamenti (metodo fitosociologico). I raggruppamenti sono indicati in alto, e si ottengono dall’analisi delle diramazioni del dendrogramma ai vari livelli di dissimilarità.
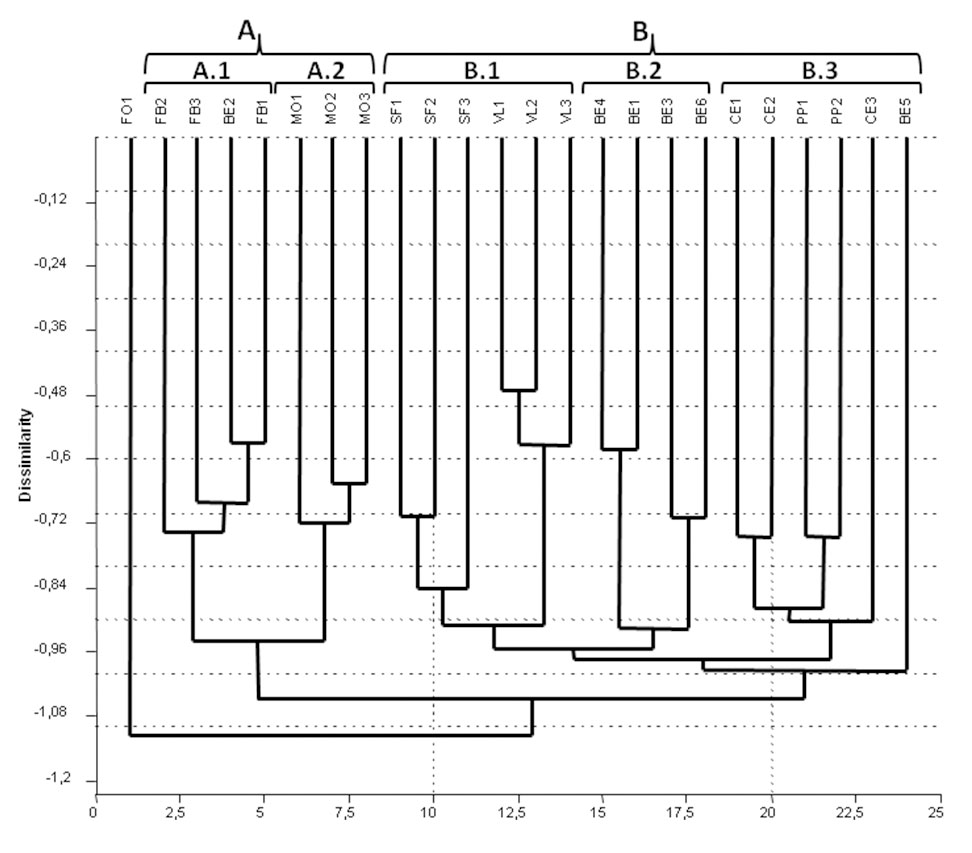
Nei restanti rilievi si individuano due gruppi principali, A e B, suddivisi rispettivamente in due e tre sottogruppi, illustrati qui di seguito.
Gruppo A
Contiene faggete su substrato siliceo, in particolare Verrucano e Arenaria. Il gruppo è caratterizzato dalla presenza importante di Ilex aquifolium e Castanea sativa, ma anche di Rubus hirtus e Carpinus betulus.
- Sottogruppo A.1: raggruppa le faggete del Bardellone con un rilievo del Monte Penna. Tale vicinanza è spiegabile per la presenza di castagno nello strato codominante (molto raro negli altri rilevamenti del Monte Penna), e in misura minore anche per la presenza di Ruscus aculeatus. I rilievi in questione presentano un ridotto numero di specie, il che rafforza il contributo delle somiglianze viste.
- Sottogruppo A.2: comprende le faggete di Montieri, caratterizzate dalla presenza di Acer pseudoplatanus, Mercurialis perennis, Moehringia trinervia, Mycelis muralis, Stellaria media.
Gruppo B
Comprende situazioni piuttosto varie, accomunate dalla presenza abbondante di Ostrya carpinifolia, e in misura minore di Melica uniflora, Acer obtusatum e Quercus cerris.
- Sottogruppo B.1: contiene faggete che crescono su substrato vulcanico (Sassoforte, Valle del Lente). Ostrya carpinifolia, Festuca heterophylla e Polypodium vulgare sono in cinque situazioni su sei, le prime due con abbondanza minore. Anche Fraxinus ornus e Melica uniflora sono molto ben rappresentate, soprattutto nella Valle del Lente.
- Sottogruppo B.2: comprende quattro fitocenosi del Monte Penna, dove le specie più comuni sono Lathyrus venetus, Daphne laureola, Viola reichenbachiana e Anemone nemorosa. Anche Luzula forsteri e Melica uniflora sono piuttosto diffuse, ma con scarsa abbondanza. Si tratta in molti casi di specie a gravitazione appenninico-balcanica. È da segnalare l’assenza di Mercurialis perennis.
- Sottogruppo B.3: comprende le faggete del Monte Cetona e di Pietraporciana, entrambe nello stesso comprensorio e su litologia calcarea, insieme a un rilievo del Monte Penna: in questo caso i motivi sono probabilmente da ricercare nella presenza di Ostrya carpinifolia e Mercurialis perennis. Il carpino nero è anche la specie che accomuna tutte e sei le faggete del gruppo, insieme a Acer obtusatum, e anche a Quercus cerris e Viola alba, sebbene con minore abbondanza.
Sinecologia
L’ordinamento dei rilevamenti sulla base degli indici di Ellenberg secondo Pignatti ([45]), fornisce ulteriori spunti per l’interpretazione sinecologica delle comunità esaminate, anche se non si evince una chiara relazione con i gruppi floristici sopra riconosciuti.
Le due componenti principali estratte dall’analisi spiegano, rispettivamente, il 52.4% ed il 40.6% della variazione totale nella matrice di varianza-covarianza di partenza. Pur essendovi solo limitate variazioni nei principali fattori ecologici, fatto legato alla relativa omogeneità ecologica delle faggete, il diagramma a dispersione (Fig. 5) mostra una ripartizione dei rilevamenti sostanzialmente in funzione della reazione del substrato (R). Questo fattore, infatti, presenta la più elevata correlazione con la componente 1 (r = 0.89). I campioni fitocenotici caratterizzati da una flora meno acidofila sono raggruppati sul quadrante destro. Si nota che, pur prevalendo i rilevamenti effettuati sulle matrici litologiche calcaree delle aree meridionali di Monte Cetona, Pietraporciana e Monte Penna, questo spazio ecologico racchiude anche tre campioni su substrato siliceo delle più settentrionali Colline Metallifere (SF, FB), probabilmente a causa del suolo molto profondo, umificato e ricco in basi. Tuttavia, la maggioranza dei rilevamenti di quest’ultima area, in particolare quelli di Montieri, così come anche quelli della Valle del Lente, risultano maggiormente caratterizzati in senso acidofilo, analogamente ad alcuni rilevamenti al contatto arenaria-calcare del Monte Penna dove erano infatti presenti Castanea sativa ed Avenella flexuosa. Questo quadro indica che il grado di acidofilia della flora fagetale eterotopica risente di fattori molto locali, solo in parte legati alla matrice geolitologica sottostante; fra questi possono esservi le condizioni microclimatiche, la micromorfologia stazionale e tipo ed intensità delle attività antropiche pregresse ed attuali.
Fig. 5 - Diagramma a dispersione della PCA basata sugli indici di Ellenberg delle specie nei rilevamenti. I cerchi neri indicano i singoli rilevamenti accompagnati dalla rispettiva sigla, mentre le frecce derivano dal biplot ed indicano la dipendenza della distribuzione dei rilievi dai parametri ecologici considerati. (N): nutrienti; (U): umidità; (C): continentalità; (L): luce; (T): temperatura; (R): reazione del suolo.
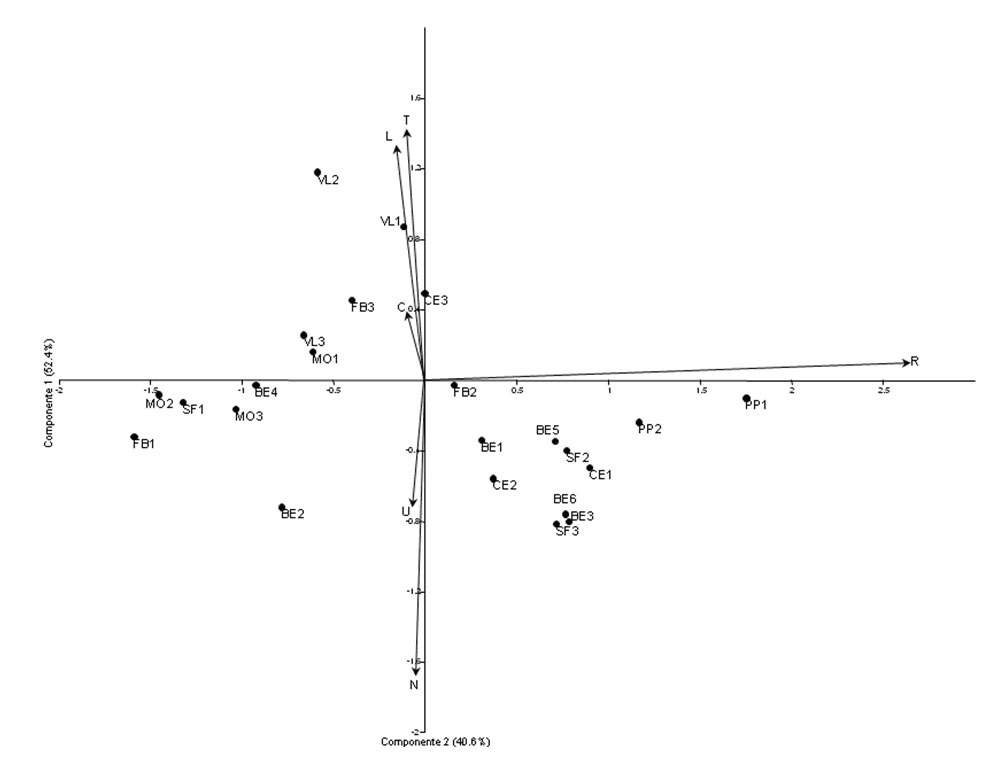
La variazione opposta di umidità e nutrienti da una parte e quella di luce e temperatura dall’altra determinano un’ulteriore ripartizione dei rilevamenti in funzione della seconda componente. Quelli con flora più spiccatamente mesosciafila ed esigente di nutrienti sono raggruppati nella parte bassa, e sono rappresentativi di faggete cacuminali o di dorsale, poste generalmente sopra i 700 m s.l.m., quindi relativamente fresche e con suolo eutrofico. Al contrario, i rilevamenti di faggeta abissale del Fosso Bardellone e della Valle del Lente, a quote inferiori a 500 m, sono caratterizzati da flora relativamente più termofila, che li dispone in maggioranza sui valori positivi delle ordinate (principalmente gruppo B).
Diversità floristica
La flora rilevata nelle faggete relitte conta 126 specie di piante vascolari, con un numero medio di 19-20 taxa per rilevamento (superficie di 200 m2 ). La variabilitàè tuttavia ampia, andando da 6 a 39 specie (CV = 39 %). Prevalgono le forme erbacee perenni, sia emicriptofite (H = 41 %), che geofite (G = 35 %), cui seguono le piante legnose (fanerofite, P = 30 %). Queste percentuali, che danno indici H/T = 3.7 e H/G = 1.1, sottolineano il carattere nemorale del popolamento vegetale delle fitocenosi forestali, in cui sono praticamente assenti sia le forme terofitiche che quelle camefitiche. Dal punto di vista corologico, il quadro è confermato dalla prevalenza di specie europee e boreali sensu lato, mentre la scarsa rappresentanza di specie mediterranee evidenzia come l’ecosistema faggeta sia in grado di mantenere al suo interno condizioni microclimatiche a carattere mesotermico anche a bassa quota e, in diversi casi, in siti non distanti dal mare. Le euri-mediterranee infine confermano la natura di transizione del territorio in esame tra la zona mediterranea e quella centro-europea.
Le variazioni che gli spettri corologico e biologico subiscono passando da un’area all’altra sono molto limitate e non sembrano meritevoli di commento.
Da evidenziare il fatto che tre entità sono considerate “di pregio” nel Manuale Italiano di Interpretazione degli Habitat della Direttiva Europea 92/43 “Habitat” ([10]): Epipactis microphylla (Cetona e Pietraporciana), Neottia nidus-avis (Sassoforte) e Aquilegia vulgaris (Fontalcinaldo). A questo si aggiunge l’evidenza che, nel territorio di studio, alcune specie sono confinate ai nuclei di faggeta, quali ad esempio le Poaceae nemorali Festuca altissima e Milium effusum e la Brassicacea mesoigrofila Cardamine kitaibelii.
Il valore medio dell’indice di Shannon per l’intero campione di rilevamenti è H’=1.48, con valori oscillanti fra 0.43 e 2.86 (Tab. 3). Complessivamente la diversità floristica sembra lievemente maggiore nelle faggete perlopiù su substrato calcareo del gruppo B (H’ = 1.54), che in quelle prevalentemente su silice del gruppo A (H’ = 1.15).
Tab. 3 - Rilevamenti e diversità. (Gruppo): ricavato da analisi multivariata; (H’): Indice di Shannon.
| Comprensorio | Rilievo n. | Gruppo | Num. dispecie | H’ |
|---|---|---|---|---|
| Monte Penna | 1 | B2 | 39 | 1.8 |
| 2 | A1 | 14 | 0.56 | |
| 3 | B2 | 21 | 0.8 | |
| 4 | B2 | 28 | 2.34 | |
| 5 | B3 | 29 | 2.02 | |
| 6 | B2 | 14 | 0.43 | |
| Colline Metallifere | 7 | B1 | 20 | 1.41 |
| 8 | B1 | 28 | 1.71 | |
| 9 | B1 | 20 | 1.28 | |
| 10 | A1 | 11 | 0.78 | |
| 11 | A1 | 11 | 1.27 | |
| 12 | A1 | 14 | 0.94 | |
| 13 | A2 | 23 | 2 | |
| 14 | A2 | 17 | 1.31 | |
| 15 | A2 | 16 | 1.18 | |
| 16 | - | 33 | 2.86 | |
| Valle del Lente | 17 | B1 | 29 | 2.19 |
| 18 | B1 | 21 | 2.68 | |
| 19 | B1 | 20 | 1.66 | |
| Cetona-Pietraporciana | 20 | B3 | 8 | 0.57 |
| 21 | B3 | 6 | 0.73 | |
| 22 | B3 | 15 | 1.63 | |
| 23 | B3 | 21 | 1.98 | |
| 24 | B3 | 21 | 1.44 | |
| Medie | Gruppo A | 15.14 | 1.15 | |
| Gruppo B | 21.25 | 1.54 | ||
| Totale | 19.96 | 1.48 | ||
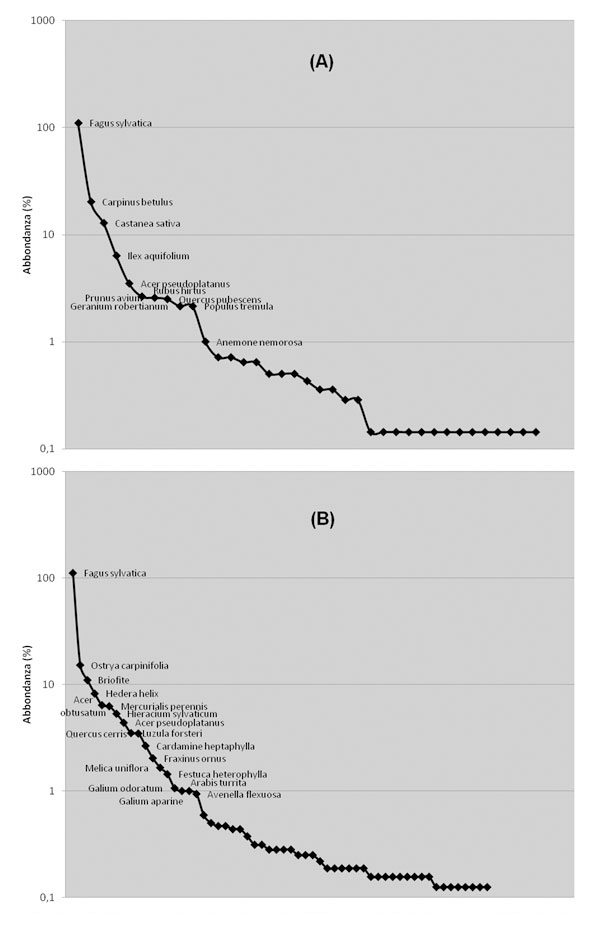
Nelle curve dominance-diversity (Fig. 6) si conferma la costante dominanza del faggio e la presenza di un discreto numero di specie sugli stessi livelli di abbondanza che porta ad appiattimenti in determinati tratti. Le altre specie che contribuiscono alla copertura sono quelle del piano arboreo dominato, in particolare Ostrya carpinifolia, Carpinus betulus, Acer obtusatum e Acer pseudoplatanus. Castanea sativa e Ilex aquifolium risultano importanti per le fitocenosi del gruppo A, dove localmente sono molto abbondanti.
Fig. 6 - Dominance-diversity curves dei gruppi (A) e (B). Le specie con la maggiore dominanza sono riportate con il nome latino. La scala dell’abbondanza è logaritmica.
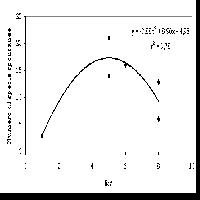
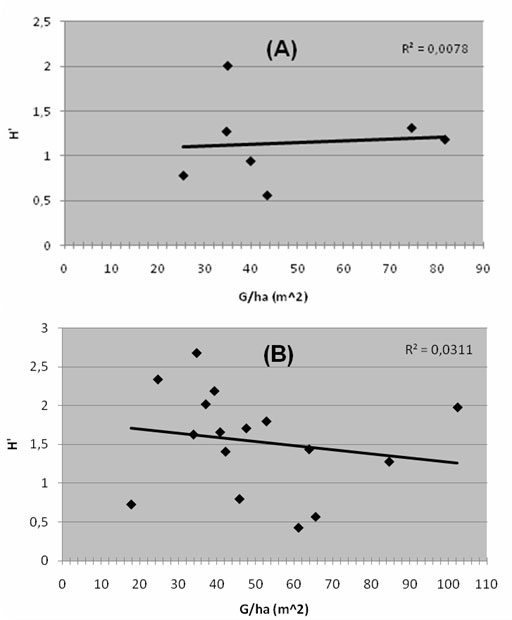
Nessuna correlazione tra indice di diversità H’ ed area basimetrica del faggio è emersa per i rilevamenti del gruppo A, mentre essa è risultata tendenzialmente negativa per quelli del gruppo B (Fig. 7), seppur non significativa (r s = -0.28, P > 0.05). Anche considerando il semplice numero di specie non si è riscontrata alcuna correlazione.
Fig. 7 - Correlazione diversità /struttura dei gruppi (A) e (B). La distribuzione dei rilevamenti nei due gruppi secondo il dato di area basimetrica ad ettaro in funzione della diversità non riporta in nessun caso un valore significativo dell’indice di correlazione. (H’): indice di Shannon; (G): area basimetrica. (R2): indice di correlazione.
Discussione e conclusioni
Il presente contributo allarga le basi conoscitive su un particolare ambiente forestale di natura relitta dell’Antiappennino tirrenico, portando alla luce nuove stazioni di faggio di tipo sia abissale (ad es., Fosso del Bardellone) che cacuminale (ad es., Sassoforte). Al pari di quelle già note, come ad esempio Pietraporciana e l’Alta Valle del Lente ([28], [51]), esse meritano un riconoscimento come biotopi di rilevante interesse conservazionistico.
Dalle analisi dendrometriche è emersa una certa disformità strutturale dovuta soprattutto agli interventi pregressi e al tempo di abbandono dei soprassuoli. In generale però si evidenzia un’elevata densità delle piante e alti valori di area basimetrica all’ettaro che, assieme alla presenza di rinnovazione, sottolineano le buone potenzialità di sviluppo intrinseche ai popolamenti e la loro capacità di recupero, indipendentemente dalle differenze nella sinecologia e nella diversità floristica. A dimostrazione di ciò stanno le faggete di Pietraporciana e Sassoforte, che grazie ad un lungo periodo di abbandono, oggi mostrano caratteristiche strutturali e biocenotiche di “bosco vetusto” o potenzialmente tale su scale temporali brevi.
Sintassonomia e sinecologia
Dal punto di vista fitosociologico e sinecologico, emerge innanzitutto che le faggete extrazonali non rientrano nelle conformazioni tipiche di quelle appenniniche, e che sono caratterizzate da contaminazioni evidenti tra la flora fagetale tipica (sensu [44]) e quella termofila delle formazioni submediterranee circostanti ([2], [48], [51]). Come già accennato, la loro estensione relativamente ridotta ha inoltre impedito di analizzare un elevato numero di aree di saggio. Ciò premesso si è potuto osservare:
- la presenza sui substrati arenacei e sul verrucano di un limitato contingente di specie acidocline (Castanea sativa, Pteridium aquilinum. Cytisus scoparius, Avenella flexuosa);
- la contaminazione, più o meno importante, da parte di specie relativamente termofile e/o dei Quercetalia pubescenti-petraeae;
- la presenza in molti gruppi di specie a gravitazione meridionale (ad es., Ilex aquifolium, Ruscus aculeatus, Cyclamen hederifolium, Digitalis micrantha, Euphorbia amygdaloides, Allium pendulinum, Lathyrus venetus, Daphne laureola, Melica uniflora, Potentilla micrantha). Non è stato mai rilevato Taxus baccata, ma è noto che la specie è presente sia nella zona del Monte Penna ([29]) che in quella di Bardellone - Monte Alto, Val di Farma ([20], [17], [38]).
Dal punto di vista strettamente sintassonomico, la sola presenza di Ilex aquifolium, anche se localmente abbondante, non permette di riferirsi alla nota associazione Aquifolio-Fagetum, la cui presenza è ipotizzata anche nella vicina Val di Farma da Pignatti ([44]), e riportata per l’Alto Lazio ([48]). Infatti Aquifolio-Fagetumsensu Gentile ([30]) è nome illegittimo, corrispondente ad Anemono apenninae-Fagetum ([30]) Brullo 1983 ([24], [26]). Il problema non è solo di natura nomenclaturale, poiché Anemono apenninae-Fagetum è strettamente legato all’area appenninica meridionale e contiene specie che non compaiono né nei nostri rilievi né nell’Italia centrale, quali Geranium versicolor, Doronicum orientale, Luzula sicula, Festuca exaltata, ecc. ([30], [11], [24]). La presenza sul M. Penna di Aquifolio-Fagetum era già stata esclusa anche da Angiolini et al. ([1]), che avevano effettuato tre rilievi in faggeta e ne avevano assegnati due a Aceri platanoidis-Fagetum e uno, dubitativamente, all’alleanza Cephalanthero-Fagion. Riguardo alle faggete appenniniche, Biondi et al. ([11]) considerano due alleanze, Aremonio-Fagion, settentrionale e Geranio versicoloris-Fagion (=Geranio striati-Fagion), meridionale. Nella prima stabiliscono una suballeanza, Cardamino kitaibelii-Fagenion, corrispondente a Geranio nodosi-Fagion, che comprende le faggete appenniniche su calcare fino all’Abruzzo. Istituiscono anche una nuova associazione centro-nord-appenninica, Lathyro veneti-Fagetum, che insieme ad un’altra associazione esistente, Staphyleo-Fagetum, assegnano all’alleanza meridionale Geranio versicoloris-Fagion. Nell’Appennino centro-settentrionale si troverebbero quindi faggete di quote più elevate, di pertinenza di Cardamino kitaibelii-Fagenion, e faggete di quote più basse, di pertinenza di Geranio versicoloris-Fagion. Dal punto di vista floristico generale, i rilievi da noi effettuati risultano piuttosto impoveriti. Quelli relativi ai substrati calcarei, evidenziati a destra nella tabella in Appendice 3, mostrano alcune affinità sia con l’associazione Aceri platanoidis-Fagetum che con l’alleanza Cephalanthero-Fagion, già riportate per il M. Penna da Angiolini et al. ([1]), ma la corrispondenza floristica è piuttosto debole; relativamente maggiore è la somiglianza con la già ricordata associazione Lathyro veneti-Fagetum, alla quale possono essere attribuiti, almeno in via provvisoria, più per le differenziali di alleanza ed il contesto cenologico che non per la frequenza delle differenziali di associazione. I rilievi dei substrati arenacei e silicei, a sinistra nella tabella in Appendice 3, sono molto simili dal punto di vista floristico-ecologico all’associazione Fraxino orni-Fagetum, descritta in passato ([2]) e riconfermata recentemente ([26]) per i Monti della Tolfa nel Lazio; gli autori recenti attribuiscono Fraxino orni-Fagetum, seppur in via provvisoria, all’alleanza meridionale Geranio versicoloris-Fagion. Anche in questo caso la somiglianza coi nostri rilievi è dovuta più al contesto cenologico generale (presenza costante di Castanea sativa, Hedera helix, Ruscus aculeatus, Melica uniflora, ecc.) che non alle differenziali di associazione, tra cui comunque figura Ilex aquifolium. Evidenziando le specie differenziali delle due alleanze/suballeanze (Appendice 3), si vede che, almeno sui substrati calcarei, le specie dell’alleanza meridionale sono più diffuse. Come già accennato, in generale i nostri popolamenti sono poco caratterizzati dal punto di vista floristico, mancando sia delle specie più tipiche di un’alleanza che dell’altra (Geranium nodosum, G. versicolor) e contenendo numerosi elementi a gravitazione sia centro-europea che appenninico-balcanica. Sebbene la presenza di alcune specie (in particolare Ruscus aculeatus) sia stata senz’altro favorita dal disturbo portato in tempi relativamente recenti tramite il governo a ceduo, in altri casi (Geranium nodosum, Cephalantera damasonium, Anemone nemorosa, Ilex aquifolium) questa indeterminatezza conferma che, come supposto, nel territorio della Toscana meridionale avviene il contatto tra i boschi di faggio provenienti dai rifugi meridionali e le faggete centro europee.
Riepilogando si può quindi affermare che, nonostante le difficoltà dette (limitate estensioni, contaminazioni floristiche, influenza del disturbo antropico, relativa povertà del corteggio floristico), la frequenza di specie differenziali di alleanza, insieme al contesto cenologico permettono di attribuire le faggete calcaree rilevate all’associazione Lathyro veneti-Fagetum, e quelle a pedologia arenacea e silicea a Fraxino orni-Fagetum (Box 1). In entrambi i casi si tratta di attribuzioni provvisorie, basate su corrispondenze floristiche non troppo forti, ma che comunque sono state chiaramente evidenziate dalle analisi effettuate, e sono dotate di significatività sintassonomica.
Box 1 - Schema sintassonomico
Il seguente schema sintassonomico è stato individuato per gli ambienti oggetto di studio:
- Querco-Fagetea Br.-Bl. et Vlieger in Vlieger 1937
- Fagetalia sylvaticae Pawlowski. in Pawlowski, Sokolowski et Wallisch 1928
- cfr. Geranio versicoloris-Fagion sylvaticae Gentile 1969
- Lathyro veneti-Fagetum sylvaticae Biondi, Casavecchia, Pinzi, Allegrezza et Baldoni 2002
- Fraxino orni-Fagetum sylvaticae Ubaldi 1995
Riguardo all’habitat di appartenenza però, nonostante le faggete appenniniche siano di difficile interpretazione ([25]), per le cenosi in studio non vi sono molti dubbi: se si prende a riferimento la Direttiva europea 92/43 (“Direttiva Habitat”), si deve affermare che tutti i popolamenti analizzati rientrano tra i “Faggeti degli Appennini con Taxus e Ilex” (codice Natura 2000: 9210*), che raggruppano infatti le faggete meridionali insieme a quelle neutrofile dell’Appennino settentrionale e delle Alpi marittime. L’ingressione di elementi termofili, nonché la presenza di substrati di varia natura, sono entrambi contemplati in questo tipo di habitat dalla Direttiva (All. 1) e dal relativo Manuale di Interpretazione europeo ([27]). Anche il recente Manuale Italiano di Interpretazione degli Habitat della Direttiva ([10]) include nell’habitat 9210 sia le faggete centro-settentrionali riferibili alla sub-alleanza Cardamino kitaibelii-Fagenion (= Geranio nodosi-Fagion), che le faggete meridionali dell’alleanza Geranio versicoloris-Fagion (=Geranio striati-Fagion) e riporta inoltre che le faggete del Cephalanthero-Fagion sono presenti solo sulle Alpi ma non in Appennino.
Diversità floristica
Le faggete analizzate ospitano un discreto numero di specie, nonostante la loro limitata estensione. Sotto il profilo qualitativo, è importante osservare che alcune di esse sono legate, nell’area di studio, a questo tipo di habitat che quindi rappresenta per esse un vero e proprio rifugio. È questo il caso di entità mesosciafile come Cardamine kitaibelii, Festuca altissima, Milium effusum ed altre, la cui presenza dimostra condizioni di suolo localmente fertile e ben umificato in aree dove spesso intorno vi sono marcati fenomeni di impoverimento ed erosione oppure terreni agricoli. Ciò pone in evidenza il ruolo della faggeta come sistema forestale in grado di contrastare la perdita di suolo e di fertilità in aree con un certo grado di siccità estiva.
I dati rilevati non supportano una chiara relazione con la densità del faggio in termini di area basimetrica, né considerando l’indice H’ né come semplice numero di specie. Questo risultato è in linea con quanto recentemente rilevato per i boschi del Tilio-Acerion nelle foreste casentinesi ([46]). Nel caso delle faggete qui esaminate, questo può dipendere dal fatto che altre specie arboree contribuiscono in modo sostanziale alla copertura complessiva del suolo, esercitando quindi una forte selezione sulla flora erbacea al pari del faggio stesso. Sono quindi necessarie analisi di densità di tutte le componenti fanerofitiche per valutare la loro influenza sulla ricchezza floristica del piano erbaceo.
Implicazioni per la gestione e la conservazione
Dal punta di vista conservazionistico, la natura relitta delle faggete extrazonali ne aumenta la vulnerabilità verso i possibili fenomeni di riscaldamento climatico globale e la documentata diminuzione delle precipitazioni in atto nella Toscana meridionale ([37]), rendendo cruciale il tipo di gestione forestale che in esse verrà praticato. È ipotizzabile che le utilizzazioni a ceduo e le conseguenti alterazioni microclimatiche in senso continentale e perdita di fertilità avranno probabilmente un impatto maggiore che non sulle faggete zonali. Il caso della faggeta di Fontalcinaldo, sottoposta negli anni scorsi a taglio ceduo tradizionale, mostra che queste utilizzazioni favoriscono specie termofile come carpino nero ed orniello, compromettendo la rinnovazione del faggio e quindi la sua persistenza nella dinamica evolutiva del bosco.
La strategia gestionale principale dovrebbe quindi basarsi su tecniche proprie di una selvicoltura prossima alla natura, al fine di favorire la conservazione di tali popolamenti, escludendo interventi di ogni genere nelle fustaie di Pietraporciana e Sassoforte, così come nelle faggete abissali della valle del Lente e del Fosso Bardellone. Negli altri casi, interventi di diradamento dal basso volti alla conversione all’alto fusto potrebbero accelerare un’evoluzione naturalmente già innescata ma ancora in fase arretrata, dove ci siano buone potenzialità per il faggio e assenza di particolari rischi di erosione del suolo e perdita di fertilità (ad es., Monte Cetona).
Sarebbe infine importante analizzare e monitorare, la quantità e la frequenza della produzione di frutto da parte del faggio, per poter programmare gli interventi sulla base della capacità ed i cicli di rinnovazione da seme del bosco.
Ringraziamenti
Si ringraziano sentitamente per l’insostituibile aiuto fornito nei rilevamenti in bosco, nonché per il supporto e l’incoraggiamento morale, i seguenti colleghi del corso in Gestione dei Sistemi Forestali dell’Università degli studi di Firenze: Eunice Assako, Federico Guasconi, Elisa Locandro.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Supplementary Material
Principali caratteristiche stazionali.
Specie sporadiche presenti nei rilevamenti con valori di abbondanza/dominanza di “+” o “r”.
Gruppi A1, A2 e B1: cfr. Fraxino orni-Fagetum sylvaticae (Ubaldi 1995). Gruppi B2 e B3: cfr. Lathyro veneti-Fagetum sylvaticae (Biondi et al. 2002). ril. 16: faggeta degradata a bosco misto, potenzialmente cfr. Lathyro veneti-Fagetum sylvaticae (Biondi et al. 2002).