Evolution of revegetated ski slopes in different environments
Forest@ - Journal of Silviculture and Forest Ecology, Volume 8, Pages 208-215 (2011)
doi: https://doi.org/10.3832/efor0681-008
Published: Dec 19, 2011 - Copyright © 2011 SISEF
Research Articles
Abstract
Revegetation of ski slopes is a useful technique to limit soil erosion, reduce the visual impact of the tracks and lengthen the duration of snow cover. Restoration is often performed with commercial forage mixtures with the aim of creating a fast soil cover, then allowing the natural recolonization of artificial swards in the mid-long term. To investigate on the recolonization dynamics, data were collected from 21 different plots from the Alps and the Apennines (Valtellina, Plan de Corones, Sappada, Cimone). Knowledge of both the original mixtures used for restoration and the timespan since intervention (ranging from 1 to 21 years) allowed to throw light on the naturalization process for the studied plots. Ground cover, floristic richness and relative presence of sown and native species were measured along linear transects established on the analyzed ski tracks. Results showed the effectiveness of plant restoration, in terms of soil coverage and (in some cases) persistence of species of the original mixtures. Recovery of autochthonous species was strongly affected by site elevation and time elapsed since restoration. Moreover, the distance of ski lanes from forest edges seems to influence the dynamics of recolonisation process. Renaturalization was remarkably faster in the lower-altitude Apennine study plot. Application of a regression analysis revealed that elevation and timespan since restoration may be considered useful predictors of the level of naturalization of the restored canopies.
Keywords
Specific contribution, Elevation, Native species, Linear analysis, Mixtures
Introduzione
Gli sport invernali rappresentano uno dei più importanti introiti economici delle località situate in aree montane. Il vivo interesse turistico verso questi sport ha stimolato la crescita e la diversificazione dell’offerta, spingendo verso la creazione di nuove piste da sci per attirare un flusso sempre crescente di turisti ([13]). Se questo da un lato favorisce lo sviluppo economico dei comuni situati nelle aree di montagna, dall’altro mette a serio repentaglio l’ambiente montano, visto che la creazione delle piste da sci e delle altre infrastrutture legate agli sport invernali può avere effetti drammatici sui fragili ecosistemi montani, soprattutto a livello del suolo e della vegetazione ([19], [16]). Il continuo passaggio di sciatori e di mezzi battipista provoca infatti il compattamento dello strato nevoso aumentando le possibilità di un congelamento del suolo, alterandone i cicli bio-chimici e diminuendone la microporosità e la permeabilità ai gas, effetti questi che si ripercuotono sulla vegetazione ([26]). Inoltre, nei periodi dell’anno in cui la superficie di una pista da sci non è ricoperta dalla coltre nevosa, il pericolo di erosione del suolo è altissimo ([18]). E’ stato stimato infatti che è necessaria una copertura vegetale superiore al 70% per contrastare significativamente l’erosione in superfici poste in pendenza ([17]).
Quanto detto sottolinea l’importanza della pratica che prevede l’inerbimento delle piste da sci nelle aree montane ([2]). Questa pratica consente infatti di limitare l’erosione superficiale, di ridurre l’impatto visivo determinato dall’alterazione e dall’eradicazione della copertura vegetale, ove presente, e di mantenere più a lungo la copertura nevosa ([10]). Rispetto ad un terreno nudo, infatti, lo scioglimento della neve sopra il tappeto erboso è molto più lento poiché la copertura vegetale blocca i flussi geotermici tra il suolo e la coltre nevosa, mantenendo costante la temperatura a livello del terreno ([7]).
Uno dei principali obiettivi a breve termine che la pratica dell’inerbimento vuole conseguire è quello di ottenere una rapida ricostituzione della copertura vegetale, ma alcuni studi condotti (ad es., [24], [12], [1], [21]) hanno mostrato la difficoltà e la lentezza di questo processo, specialmente in zone ad altitudine elevata ([9]). Spesso, inoltre, vengono utilizzate misture di sementi economiche e facilmente reperibili sul mercato ([4], [15]) che generalmente non sono quelle più idonee a rispondere alle condizioni stazionali dell’ambiente locale ([20], [25], [14]). Per queste ragioni, e per prevenire un eventuale insuccesso dell’inerbimento, è consigliabile un’analisi preliminare dell’area per individuare il mix di specie più opportune da utilizzare al momento della semina ([3]).
Una volta che la copertura del suolo è stata ottenuta grazie alle specie introdotte con la semina, l’obiettivo a medio e lungo termine è quello di ottenere un elevato livello di ricolonizzazione da parte di specie autoctone dell’ambiente locale, per garantire l’integrazione ecologica delle aree sottoposte ad intervento artificiale ([12], [1]). Secondo Bédécarrats ([8]) è possibile identificare tre stadi di evoluzione della comunità vegetale a seguito di un inerbimento di una pista da sci: una prima fase dominata dalle specie seminate, una seconda durante la quale le specie autoctone cominciano a “contaminare” il prato artificiale con la loro presenza, ed una terza, considerato stadio di maturazione, dove le specie native tendono spontaneamente a diventare dominanti rispetto alle essenze introdotte con l’inerbimento. Il tasso di ricolonizzazione da parte delle specie native è influenzato in maniera notevole dalle condizioni ambientali della stazione e dalla gestione della copertura vegetale. Talamucci & Argenti ([22]), analizzando piste da sci in comprensori sciistici alpini hanno preso in considerazione una serie di parametri con l’obiettivo di verificare sia la persistenza nel tempo delle essenze impiegate nel miscuglio originale che il grado di rinaturalizzazione dei cotici tramite il reingresso della flora autoctona, evidenziando come la rapidità del processo di ricolonizzazione sia legata soprattutto all’altitudine della stazione e alla distanza in anni dalla semina e questo è stato confermato anche in indagini successive ([6]).
Il lavoro qui presentato analizza la situazione di piste da sci inerbite collocate in areali montani diversificati per ottenere informazioni utili sul successo delle operazioni di inerbimento e sull’evoluzione della vegetazione presente.
Materiali e metodi
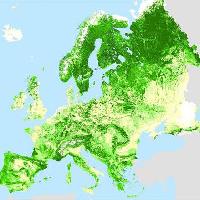
Le zone indagate nell’ambito del presente studio (Fig. 1) hanno riguardato sia comprensori sciistici posti nel settore centro-orientale dell’arco alpino (Valtellina, Plan de Corones e Sappada) che situati nell’Appennino settentrionale (Cimone).
Fig. 1 - Dislocazione delle stazioni sciistiche in cui sono stati effettuati i rilievi.
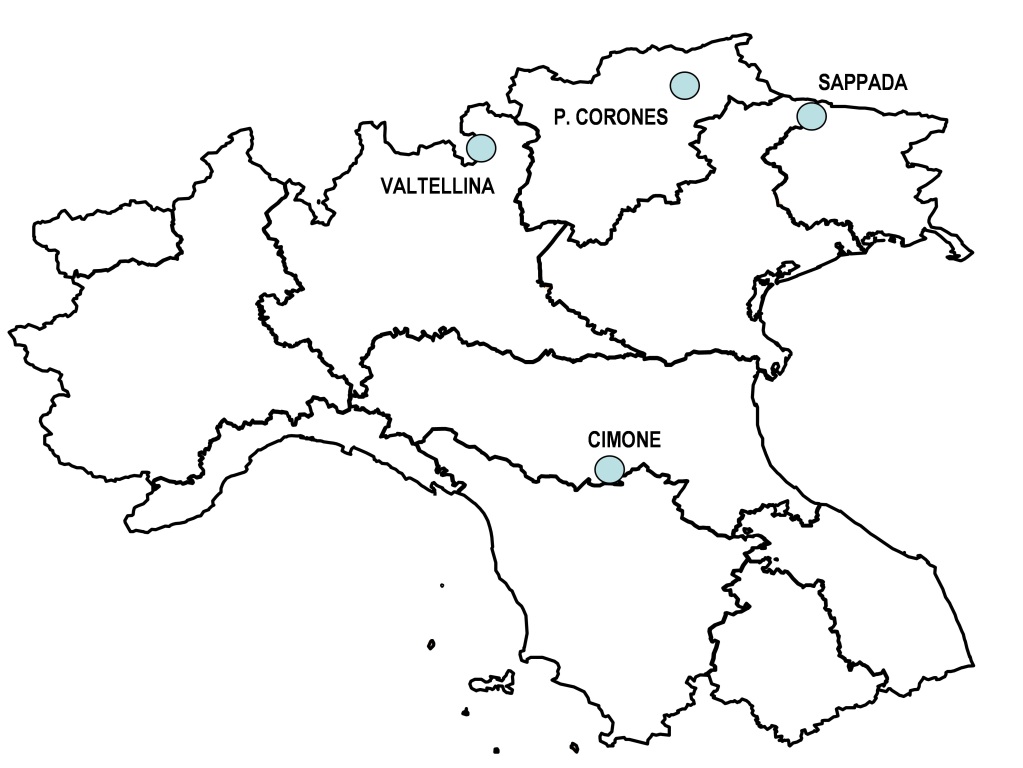
I rilievi sono stati eseguiti in diversi anni nell’ambito delle attività di ricerca sugli inerbimenti restaurativi svolte dal Dipartimento di Scienze delle Produzioni Vegetali, del Suolo e dell’Ambiente Agroforestale dell’Università di Firenze. Le stazioni erano collocate in situazioni ambientali molto diversificate (per latitudine, altitudine e tipologia di suolo) e anche gli interventi di inerbimento erano stati condotti con modalità differenziate. Inoltre, alcuni tratti di pista erano in parte localizzati in zone intraforestali ed in parte a quote poste oltre il limite superiore della vegetazione (Tab. 1). In totale sono state analizzate le coperture vegetali di 21 tratti di piste da sci all’interno dei quattro comprensori studiati, su tratti di pista con altitudine variabile da 1183 m s.l.m. della pista più bassa (Pista Rossa, Cimone) a 2240 m di quella più alta (Pista Masucco, Valtellina). Inoltre, utilizzando le informazioni raccolte, è stato possibile anche determinare l’età dei cotici erbosi al momento delle rilevazioni botaniche in campo (con un intervallo di età variabile da 1 a 21 anni) e il miscuglio utilizzato durante le operazioni di ripristino vegetale. Le formulazioni dei miscugli sono risultate molto diverse da caso a caso, dato che su alcune piste da sci sono stati utilizzati miscugli bifiti a due sole componenti (una graminacea e una leguminosa, quasi sempre costituite da Festuca gr. rubra e Trifolium repens), mentre in altri casi le miscele impiegate erano molto complesse, con formulazioni comprendenti fino ad un massimo di 15 specie.
Tab. 1 - Principali caratteristiche delle piste e degli inerbimenti studiati.
| Area | Nome pista | Presenza del bosco | Altezzamedia (m slm) | Specie nel miscuglio originario | Età |
|---|---|---|---|---|---|
| Valtellina | Masucco | no | 2240 | 2 | 5 |
| Valtellina | Masucco | no | 2240 | 5 | 5 |
| Plan de Corones | Sylvester | no | 1620 | 11 | 21 |
| Plan de Corones | Sylvester | no | 1620 | 10 | 21 |
| Plan de Corones | Pramstall | no | 2030 | 10 | 20 |
| Sappada | Turistica | si | 1425 | 13 | 18 |
| Sappada | Turistica | si | 1550 | 13 | 8 |
| Sappada | Turistica | si | 1550 | 13 | 19 |
| Sappada | Monte Siera | no | 1670 | 5 | 2 |
| Sappada | Monte Siera | no | 1670 | 10 | 2 |
| Sappada | Monte Siera | no | 1670 | 15 | 2 |
| Sappada | Sambl | no | 1880 | 3 | 3 |
| Sappada | Sambl | no | 1880 | 8 | 3 |
| Sappada | Sambl | no | 1880 | 13 | 3 |
| Monte Cimone | Rossa | si | 1183 | 2 | 8 |
| Monte Cimone | Rossa | si | 1183 | 2 | 9 |
| Monte Cimone | Passo Lupo | si | 1570 | 3 | 3 |
| Monte Cimone | Passo Lupo | si | 1570 | 3 | 4 |
| Monte Cimone | Direttissima | no | 1630 | 2 | 10 |
| Monte Cimone | Direttissima | no | 1630 | 2 | 11 |
| Monte Cimone | N. Beccadella | no | 1740 | 2 | 1 |
Su ogni tratto di pista è stato eseguito il rilevamento della composizione botanica presente tramite rilievi lineari ([11]) con i quali è stato possibile ottenere la percentuale di presenza delle specie rilevate, o contributo specifico (CS), ed il numero delle specie presenti all’interno di ogni linea di flora. Inoltre è stato effettuato anche il rilevamento della copertura (mediante stima visiva) del cotico erboso presente sul tracciato inerbito, secondo una metodologia già validamente adottata in contesti simili ([5]).
La conoscenza del miscuglio impiegato nelle operazioni di inerbimento ha permesso, in tutte le elaborazioni successive, di tenere separate le specie seminate, intendendo con questo termine quelle presenti nel miscuglio originario ([23]), da quelle insediatesi successivamente nel corso del processo di riconquista del cotico da parte della flora locale. In questo modo non solo è possibile verificare la persistenza nel tempo delle essenze impiegate nel miscuglio adottato al momento dell’intervento, ma è anche possibile stabilire il grado di rinaturalizzazione del cotico tramite l’analisi del reingresso della flora autoctona.
Risultati e discussione
L’insieme dei dati raccolti ha permesso di individuare degli andamenti generali secondo i quali avviene il processo evolutivo dei cotici introdotti con gli inerbimenti in contesti fragili dal punto di vista ecologico come quelli delle piste da sci.
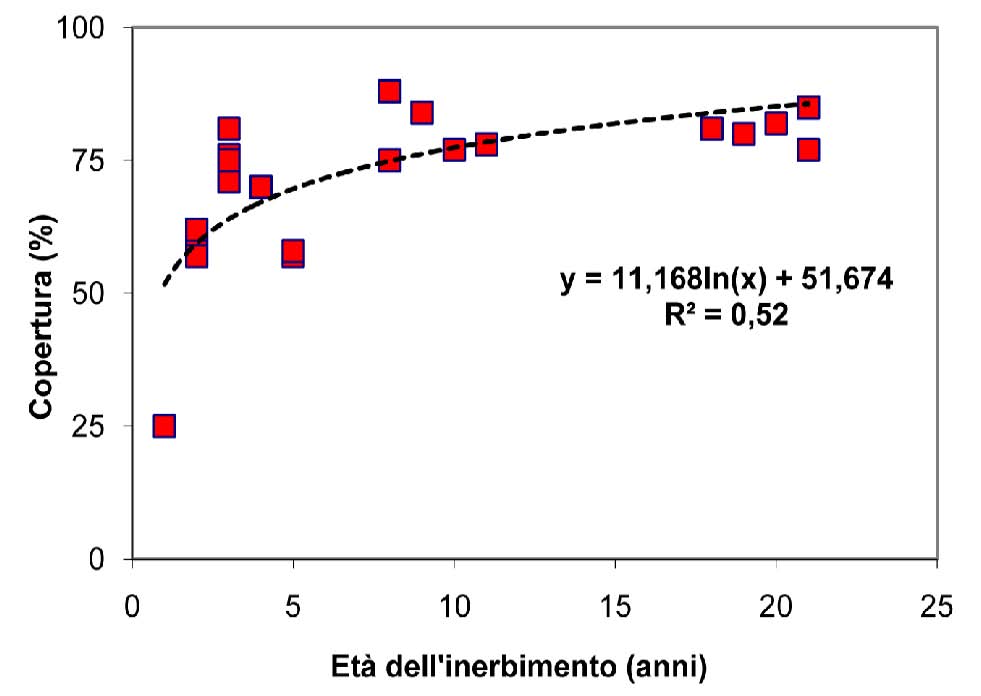
L’inerbimento dei tratti di pista presi in considerazione ha avuto successo e la copertura percentuale del suolo, influenzata in maniera sensibile dagli anni di distanza dalla semina, è risultata variabile dal 25% dopo 1 anno sul Cimone (pista Beccadella) all’88% dopo 8 anni sulla pista Rossa, sempre nello stesso comprensorio sciistico (Tab. 2). È anche vero che valori di copertura molto elevati sono stati registrati in corrispondenza di piste inerbite da un numero di anni mediamente superiore, come nel caso delle piste collocate nel comprensorio di Plan de Corones e alcuni tratti della pista Turistica a Sappada. In entrambi i casi i valori di copertura si aggirano intorno all’80% ma in corrispondenza di età del cotico decisamente superiori (tra 18 e 21 anni) rispetto a quelle caratterizzanti i tratti di pista precedentemente presi in esame. Questa considerazione evidenzia la diversa rapidità di copertura da parte del cotico in situazioni stazionali differenti, soprattutto per quanto riguarda l’altitudine dell’area sottoposta ad intervento. Infatti la pista Rossa risulta essere quella con copertura maggiore, ma anche quella situata alla quota più bassa (1183 m s.l.m.) e questo ha permesso una rapida evoluzione del cotico e un veloce ricoprimento del terreno.
Tab. 2 - Principali parametri rilevati sugli inerbimenti analizzati.
| Area | Nomepista | Copertura (%) | N. specie seminate osservate | N. speciespontanee osservate | CS specie spontanee (%) |
|---|---|---|---|---|---|
| Valtellina | Masucco | 57 | 2 | 8 | 18.2 |
| Valtellina | Masucco | 58 | 4 | 6 | 12.7 |
| Plan de Corones | Sylvester | 85 | 7 | 18 | 56.7 |
| Plan de Corones | Sylvester | 77 | 7 | 14 | 48.2 |
| Plan de Corones | Pramstall | 82 | 5 | 6 | 37.7 |
| Sappada | Turistica | 81 | 6 | 9 | 56.2 |
| Sappada | Turistica | 75 | 5 | 15 | 37.4 |
| Sappada | Turistica | 80 | 4 | 21 | 33.8 |
| Sappada | Monte Siera | 60 | 5 | 1 | 1.1 |
| Sappada | Monte Siera | 57 | 8 | 1 | 3.1 |
| Sappada | Monte Siera | 62 | 7 | 2 | 6.7 |
| Sappada | Sambl | 76 | 3 | 3 | 13.0 |
| Sappada | Sambl | 81 | 8 | 6 | 20.3 |
| Sappada | Sambl | 75 | 6 | 5 | 18.2 |
| Cimone | Rossa | 88 | 2 | 18 | 77.9 |
| Cimone | Rossa | 84 | 2 | 21 | 85.2 |
| Cimone | Passo Lupo | 71 | 2 | 9 | 56.7 |
| Cimone | Passo Lupo | 70 | 2 | 8 | 50.0 |
| Cimone | Direttissima | 77 | 2 | 12 | 63.0 |
| Cimone | Direttissima | 78 | 2 | 9 | 76.1 |
| Cimone | N. Beccadella | 25 | 2 | 1 | 16.7 |
Se confrontiamo il numero di specie del miscuglio originario (Tab. 1) con quella relativa al numero di specie seminate osservate (Tab. 2), è possibile fare alcune considerazioni circa la permanenza nel tempo delle specie introdotte con la semina. Laddove il miscuglio originario era formato da due sole specie o da un numero comunque esiguo, molte di queste si ritrovano anche a distanza di qualche anno dalla semina, mentre in caso di miscugli formati da molte componenti il numero delle specie seminate che poi viene osservato a distanza di tempo si riduce drasticamente, come ad esempio nel caso della pista Turistica nel comprensorio di Sappada, sulla quale solo 4 delle 13 specie componenti del miscuglio originario sono state ritrovate a distanza di 19 anni. È quindi possibile affermare che alcune specie presentano una fortissima capacità di adattamento e di persistenza nelle condizioni saggiate e quindi possono essere rinvenute a distanza di molto tempo dalla semina: fra queste è da considerare senza dubbio Festuca gr. rubra, che è la specie generalmente più adattabile alle condizioni stazionali presenti in queste tipologie di interventi ed infatti è la più diffusa nei miscugli specializzati per le piste da sci. La diversa durata nel tempo delle specie è influenzata dalle caratteristiche delle specie stesse. È probabile che alcune di esse abbiano un comportamento più aggressivo rispetto ad altre e che nel giro di qualche anno tendano a diventare predominanti. È anche vero però che la diminuzione del numero di specie seminate appare più netta nei cotici inerbiti da un numero maggiore di anni, i quali presentano contemporaneamente anche un numero più elevato di specie spontanee osservato. È chiaro quindi che il livello di naturalizzazione dell’inerbimento è un parametro fortemente collegato all’età del cotico. Al tempo stesso è possibile rilevare che alcune specie introdotte con l’inerbimento non sono presenti nei cotici anche a distanza di pochi anni: questo fatto è dovuto sia all’impiego di specie che devono avere un rapido ricoprimento iniziale ma poi sono destinate a sparire prontamente (le cosiddette specie starter) sia alla presenza nei miscugli complessi anche di specie che non riescono ad affermarsi in quanto assolutamente non adatte alle condizioni locali.
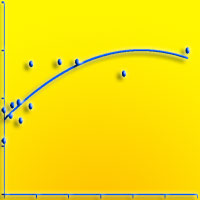
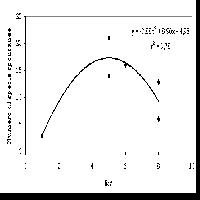
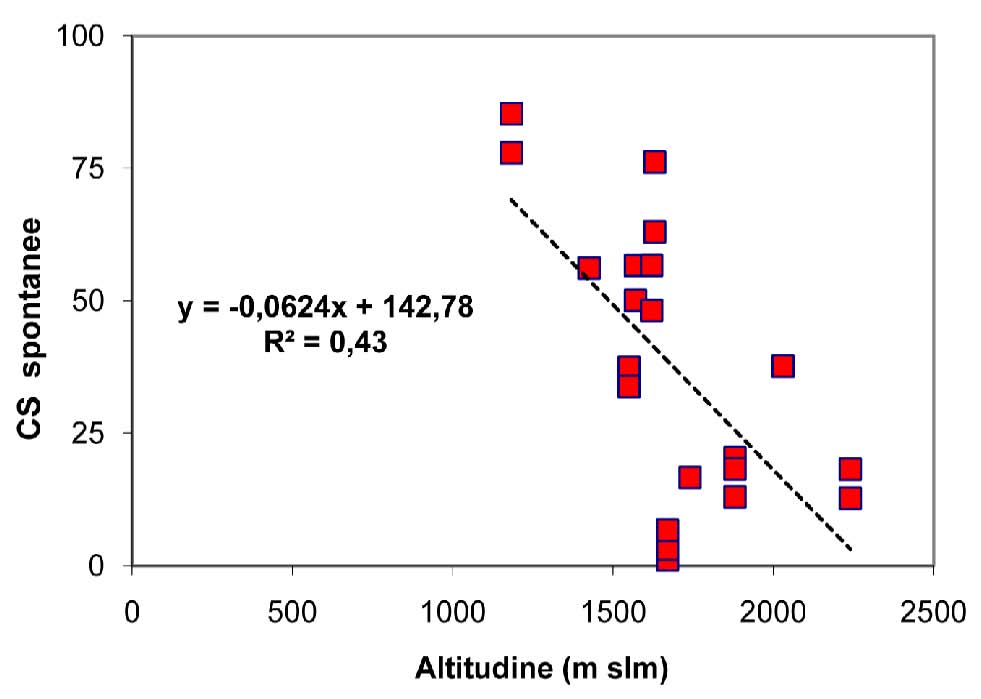
Per quanto riguarda l’altitudine, questa non sembra influenzare solo la rapidità di copertura del cotico ma anche il successivo reingresso delle specie spontanee. Il contributo specifico delle specie autoctone, ovvero la loro presenza percentuale, segue un andamento lineare inversamente proporzionale rispetto all’aumentare dell’altitudine, indipendentemente dall’età dell’inerbimento (Fig. 2). Infatti, all’abbassarsi dell’altezza della stazione, aumenta in maniera netta la presenza di flora autoctona rispetto alle specie seminate, arrivando a percentuali superiori all’80% alle quote più basse, grazie alla presenza di periodi vegetativi più lunghi e al maggior apporto di sostanza organica al terreno da parte della copertura forestale.
Fig. 2 - Contributo specifico delle specie spontanee in funzione dell’altitudine della pista.
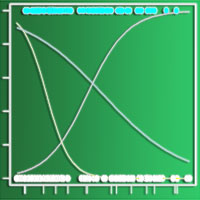
Come già visto, l’altro parametro che influenza notevolmente la persistenza delle specie seminate ed il reingresso di quelle spontanee è l’età del cotico. In Fig. 3 si può notare come la copertura percentuale aumenti all’aumentare dell’età del cotico: il rapido incremento iniziale, evidenziato dalla inclinazione della curva, è dovuto all’insediamento delle specie seminate che ritroviamo abbondantemente in cotici giovani, inerbiti da pochi anni. La curva tende poi ad assumere un andamento crescente ma meno ripido a causa del reinsediamento più graduale delle specie autoctone che concorrono a coprire il terreno. È comunque ragionevole pensare che anche alcune piante appartenenti alle specie presenti nel miscuglio originario e ritrovate a distanza di tempo durante i rilievi, siano frutto della ricolonizzazione della flora spontanea. E’ evidente che quindi il ruolo principale delle specie seminate sia quello di coprire il più rapidamente possibile il terreno, riducendo il rischio di erosione nella fase iniziale dell’inerbimento, lasciando al successivo reingresso della flora autoctona il compito di naturalizzare il cotico così creato.
Fig. 3 - Copertura del terreno in funzione dell’età del cotico.
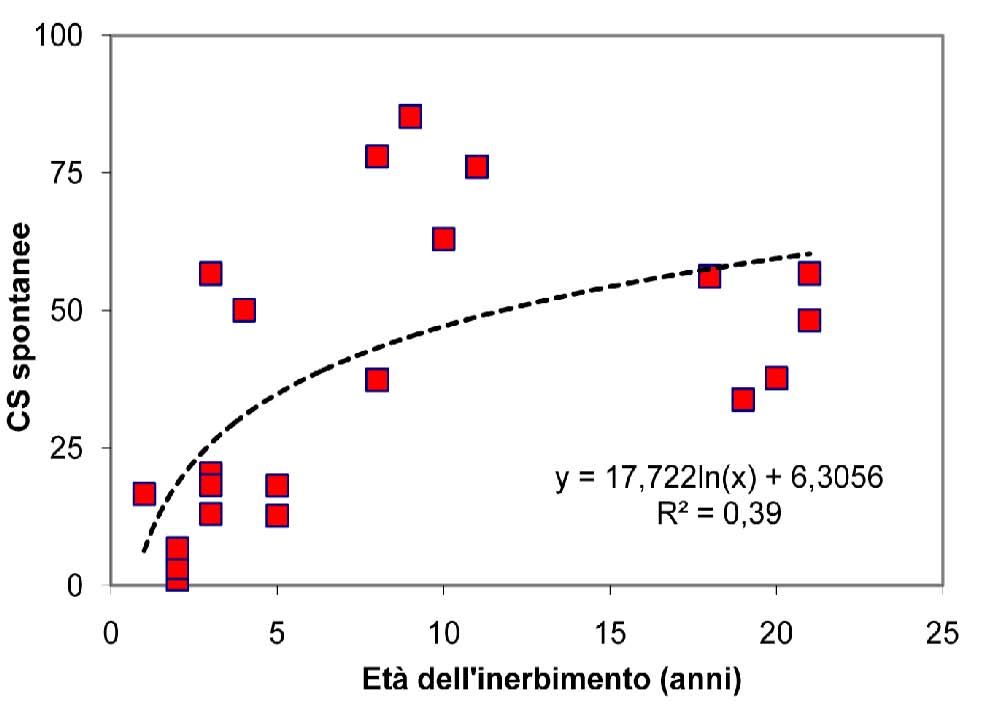
La curva relativa al numero delle specie spontanee in funzione dell’età dell’inerbimento (Fig. 4) presenta, in analogia all’andamento della curva del contributo specifico di tali specie (Fig. 5), un andamento di tipo logaritmico che indica, per le specie native, una più rapida ricolonizzazione nei primi anni ed una stabilizzazione successiva, quando il reingresso della flora autoctona tende a rallentare. I dati rilevati nell’ambito del presente studio indicano che è necessario un intervallo di tempo di circa 10-15 anni per ottenere un cotico con caratteristiche di naturalità, intendendo con questo termine una situazione di equilibrio nella quale il contributo specifico delle specie spontanee è circa la metà di quelle presenti in totale. L’affermarsi delle specie spontanee avviene comunque gradualmente e contemporaneamente alla progressiva scomparsa delle componenti seminate. Il tratto finale delle curve rappresenta la fase ultima della successione e la rinaturalizzazione del cotico che perde la connotazione di artificialità raggiungendo una forma che possiamo considerare quasi definitiva, stabile e ben inserita nell’ambiente.
Fig. 4 - Numero delle specie spontanee in funzione dell’età del cotico.
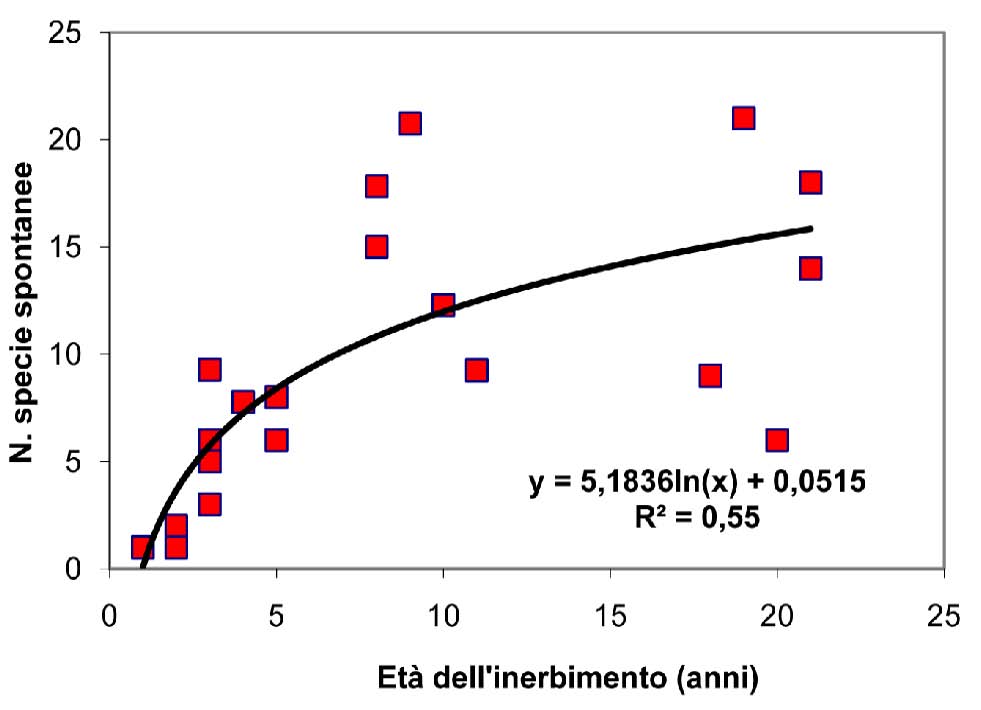
Fig. 5 - Contributo specifico delle specie spontanee in funzione dell’età del cotico.
Per valutare gli effetti della copertura arborea sui parametri analizzati sono state calcolate le medie delle principali variabili relative agli inerbimenti in funzione della presenza del bosco (Tab. 3). Essendo l’età media degli inerbimenti non significativa, è possibile attribuire le eventuali differenze di comportamento all’altro fattore che è risultato molto correlato con l’evoluzione degli inerbimenti, e cioè alla quota, e quindi anche alla collocazione delle piste rispetto al bosco. Oltre all’altitudine, i parametri che descrivono l’evoluzione della flora autoctona sono nettamente più elevati nelle piste infraforestali rispetto a quelle poste oltre il limite del bosco. Infatti non sono state riscontrate differenze significative per quanto riguarda la copertura percentuale e il numero delle specie introdotte con la semina, mentre la presenza del bosco sembra influenzare positivamente il reingresso della flora locale, sia in termini di presenza percentuale che di ricchezza floristica.
Tab. 3 - Effetto della presenza del bosco sui principali parametri osservati. (**): P < 0.01; (*): P < 0.05; (ns): non significativo.
| Presenza del bosco | Si | No | Prob. |
|---|---|---|---|
| Altezza media (m slm) | 1433 | 1814 | ** |
| Età | 9.9 | 7.8 | ns |
| Copertura (%) | 78 | 68 | ns |
| Numero specie seminate osservate | 3.3 | 4.9 | ns |
| Numero specie spontanee osservate | 14.4 | 6.6 | ** |
| Cs specie spontanee (%) | 57 | 28 | * |
Avendo valutato come i parametri più importanti nel determinare l’evoluzione della vegetazione siano l’altitudine e l’età dell’inerbimento è stata calcolata una regressione multipla per mettere in relazione il contributo specifico percentuale delle specie spontanee con i due parametri citati. La regressione ottenuta è la seguente (eqn. 1):
dove Y rappresenta la sommatoria dei CS delle specie spontanee rilevate, x1 l’età in anni del cotico e x2 l’altitudine media del tratto di pista esaminato in m s.l.m. Si può quindi affermare che, nelle situazioni analizzate, all’aumentare di un anno di età del cotico il contributo specifico totale delle specie autoctone aumenta di 1.36%, mentre all’aumentare di 100 m di quota tale contributo diminuisce del 6%. In tal modo mediante un semplice modello è possibile prevedere il tasso di ricolonizzazione di un inerbimento artificiale in funzione dei fattori più importanti nel condizionare il reingresso della flora autoctona.
Conclusioni
L’inerbimento delle piste da sci può essere effettuato utilizzando tecniche diverse e miscugli di sementi differenziati per complessità floristica ma gli scopi principali sono sostanzialmente gli stessi: nel breve termine la finalitàè quella ottenere una rapida copertura vegetale che sia protettiva del suolo e in grado di inserirsi nel paesaggio montano, mentre nel medio e lungo periodo si dovrebbe arrivare ad avere un cotico naturalizzato, stabile, formato da vegetazione in prevalenza autoctona e con ridotto connotato di artificialità.
Nell’ambito del nostro lavoro, nonostante l’eterogeneità delle situazioni saggiate, il processo di reingresso della flora locale sembra essere un fenomeno assai efficace e inarrestabile, anche se può variare localmente nella dinamica e nelle forme. Un ruolo rilevante è quindi giocato dalle specie spontanee che, nel corso degli anni, tendono a sostituire le specie introdotte con la semina, contribuendo così al reinserimento ecologico dell’area sottoposta ad intervento nel contesto circostante. Le specie seminate, infatti, una volta assolta la funzione transitoria di coprire e proteggere il terreno nell’attesa della formazione di un cotico naturalizzato, tendono progressivamente ad essere sostituite, anche se in alcuni casi sono state notate persistenze notevoli anche a distanza di 15-20 anni dalla semina, sebbene in questi casi non sia possibile stabilire se le piante presenti provengano dalla semina originaria o si siano sviluppate a partire dalla flora locale come riportato da Tallarico & Argenti ([23]). E’ quindi possibile individuare un diverso ruolo per le specie seminate rispetto a quello frutto di reingresso della flora locale: le prime svolgono essenzialmente una funzione biotecnica di difesa antierosiva, essendo presenti anche in numero ridotto ma con elevati ricoprimenti specifici, mentre le seconde svolgono un ruolo prevalentemente ecologico, contribuendo all’incremento della biodiversità e della ricchezza floristica anche se ognuna di esse è poco rappresentata come presenza percentuale.
Il processo di ricolonizzazione è influenzato dalle diverse caratteristiche stazionali che possono però essere riassunte sinteticamente nella sola altitudine come indicato da Borreani et al. ([9]). Come evidenziato nel corso del presente studio, a quote inferiori si ha un processo di ricolonizzazione delle specie autoctone più rapido e di conseguenza uno stadio stabile del cotico è raggiunto in un tempo più breve rispetto a quanto riportato da altri autori ([8] [22]). In particolare è evidente il maggior reingresso delle specie spontanee nel comprensorio appenninico, posto ad una quota nettamente più bassa delle aree alpine studiate. Anche la presenza della copertura forestale (anch’essa ovviamente da porsi in relazione all’altitudine della pista) può costituire un elemento caratterizzante notevole di una pista da sci, tanto che è possibile individuare dinamiche diverse per le piste infraforestali e sopraforestali come già trovato in lavori precedenti riferiti solo a piste alpine ([4]).
L’altro parametro fondamentale che influenza fortemente le performance di colonizzazione della vegetazione è la distanza in anni dall’inerbimento. Sia il contributo specifico delle specie native che il loro numero aumentano all’aumentare dell’età del cotico seguendo l’andamento di una curva di tipo esponenziale, evidenziando una più rapida dinamica della ricolonizzazione della flora autoctona nella fase iniziale del processo evolutivo ed un successivo rallentamento. I dati comunque dimostrano che, operando a quote non troppo eccessive, il processo di reingresso avviene inesorabilmente, anche se lentamente, e quindi in molte situazioni è possibile operare anche tramite l’adozione di miscugli molto semplificati lasciando all’evoluzione successiva il compito di rinaturalizzare i cotici creati artificialmente, come suggerito da Argenti & Ferrari ([7]).
Infine, i risultati hanno mostrato che basandosi sui parametri maggiormente connessi con la velocità del processo evolutivo è possibile creare semplici modelli del grado di naturalizzazione di un inerbimento in modo da poter prevedere la percentuale di presenza delle specie spontanee in funzione dell’altitudine della pista e dell’età dell’inerbimento.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar