Large-scale indicators for monitoring forest diversity of the main forest types in Calabria (Italy)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 14, Pages 84-93 (2017)
doi: https://doi.org/10.3832/efor2293-014
Published: Feb 27, 2017 - Copyright © 2017 SISEF
Research Articles
Guest Editors: ALForLab Project (PON03PE_00024_1 - PON R&C- PAC 2007-2013)
« Public-private lab for the application of innovative technologies with high environmental sustainability in the forest-wood-energy chain »
Collection/Special Issue: Alessandro Paletto (CREA, Trento), Giorgio Matteucci & Mauro Maesano (CNR-ISAFOM)
Abstract
Recently, the Society’s perception of forest resources has gone through significant changes. Forest ecosystems play a multifunctional role and host an important portion of the whole biodiversity, particularly in the Mediterranean area. Remote sensing technologies provide a unique way to obtain spatially extensive information on forest ecosystems, but relatively few studies used such information to evaluate forest habitat and biotic diversity. In this paper we evaluate the effectiveness of remote sensing to predict forest diversity by linking remotely sensed information with diversity metrics obtained from ground measurements of butterfly diversity. The field work was carried out in Calabria in four different forest types (beech, chestnut, black pine and silver fir forests). The sampling of Lepidoptera was carried out by LED light traps. We positioned 9 traps per forest type, for a total of 36 sites chosen to sample the different stages of forest succession in each forest type. Samples were carried out once a month from May to November 2015. Data from in situ butterfly measurements were compared with above ground forest biomass estimated from airborne LiDAR with NDVI estimated from Landsat 8. Results indicated that the Geometridae/Noctuideae ratio of lepidopteran communities was significantly correlated with the tree biomass, its distribution among tree size classes and the NDVI. The Geometridae/Noctuidae ratio, therefore, represents an index easy to calculate, which can be employed to integrate data acquired from remote sensing in order to obtain continuous spatial estimates of forest naturalness.
Keywords
LiDAR, Moths, Biodiversity, Bioindicators Taxa, Landsat, NDVI
Introduzione
Negli ultimi decenni la percezione della società nei confronti del ruolo delle risorse forestali ha subito profondi mutamenti. La gestione forestale, in passato prevalentemente orientata alla massimizzazione della produzione legnosa, si è andata configurando sulla base di nuove richieste relative alla promozione e alla salvaguardia del ruolo ecologico-ambientale, turistico-ricreativo e paesaggistico del bosco ([3]). Accanto a queste funzionalità e servizi richiesti al bosco si pone il problema della perdita di biodiversità a livello globale ([14]). Gli ecosistemi forestali ospitano una elevata porzione di biodiversità, in termini di specie, genotipi e processi ecologici. Questi aspetti assumono un significato forse ancora più importante in un ambiente come quello mediterraneo dove la notevole varietà di condizioni ambientali (climatiche, geologiche, morfologiche) e l’utilizzo secolare delle risorse territoriali hanno creato un mosaico di paesaggi culturali e naturali estremamente ricchi e diversificati. D’altro canto, la forte frammentazione e omogeneizzazione rappresentano una minaccia alla diversità biologica e alla valenza multifunzionale delle componenti naturali del paesaggio e delle risorse forestali ([15]).
L’impiego delle informazioni derivate dal telerilevamento rappresenta un importante supporto per l’osservazione e il monitoraggio delle risorse naturali e per la programmazione di azioni coordinate utili alla prevenzione degli incendi boschivi ([11]). Il settore forestale eredita una lunga tradizione di impiego di informazioni derivanti dai sistemi di osservazione della terra (OT) per la stima di attributi forestali e per la loro inventariazione: esempi delle variabili stimate includono composizione specifica, volume legnoso e suo incremento. Per maggiori dettagli, si veda Corona ([10]).
L’impiego del telerilevamento ai fini della stima e del monitoraggio della biodiversità ha ricevuto solamente in tempi recenti adeguata considerazione. Ad esempio, alcuni studi hanno utilizzato informazioni derivanti da telerilevamento da satellite per valutare la struttura e la composizione di ecosistemi forestali in ambienti tropicali/subtropicali ([22], [24]). Pochi studi invece hanno utilizzato le informazioni derivanti dai sistemi OT per valutare la diversità non-arborea di ecosistemi vegetali. Tale diversità può essere quantificata a terra selezionando comunità biotiche ritenute indicatori di particolari caratteristiche degli habitat forestali e della biodiversità. Esempi di integrazione di stime di diversità da telerilevamento mediante l’impiego di bioindicatori includono l’utilizzo di specie ornitiche ([9]), licheni ([13]), funghi ([26]). Pochissimi studi si sono focalizzati sulla valutazione degli habitat e della loro diversificazione utilizzando i Lepidotteri adulti come indicatori ecologici ([20]). Per esempio, Hess et al. ([16]) hanno utilizzato informazioni derivanti da LiDAR e da fotogrammetria aerea per valutare l’idoneità ambientale a scala di paesaggio per quattro specie diverse di farfalle. Il loro studio ha prodotto interessanti risultati, dimostrando che l’informazione tridimensionale derivabile da un dato LiDAR fornisce più informazioni rispetto ai dati ottici per caratterizzare un habitat forestale. Tuttavia, lo studio ha valutato un limitato numero di specie di Lepidotteri diurni che rappresentano una frazione minima di questo Ordine di insetti e, al contrario di quelli notturni, forniscono scarse informazioni per gli habitat forestali perché legati generalmente ad ambienti aperti e soleggiati.
I Lepidotteri Eteroceri (sottogruppo che comprende le famiglie più numerose, con abitudini generalmente notturne) hanno molte caratteristiche tipiche dei bioindicatori. Questo gruppo, infatti, è composto da specie con diversificate esigenze ecologiche, per le quali si ha una buona conoscenza della tassonomia e dell’autoecologia. Inoltre, le loro comunità reagiscono ai cambiamenti ambientali a differente scala sia spaziale che temporale. Infine, le larve, e spesso anche gli adulti, sono legati troficamente alla vegetazione, per cui le comunità sono direttamente influenzate dalla componente floristica degli ecosistemi.
La ricerca intende testare l’ipotesi secondo la quale i boschi che hanno una struttura eterogenea hanno un maggiore valore naturalistico utilizzando per la verifica i Lepidotteri notturni quali bioindicatori ([31], [12], [29], [6], [19]).
In questa prospettiva, scopo della presente sperimentazione è valutare l’impiego dei Lepidotteri a volo notturno come indicatori della biodiversità in ecosistemi forestali naturali. All’uopo, lo studio ha utilizzato uno dei principali prodotti del progetto ALForLAB (Laboratorio Pubblico-privato per l’applicazione di tecnologie innovative ad elevata sostenibilità ambientale nella filiera foresta-legno-energia - [PON03PE_00024_1 - PON R&C- PAC2007-2013]), relativo alla mappatura della biomassa di soprassuoli forestali derivata da dato LiDAR aereo (ALS), per valutare la correlazione tra stime di biomassa prodotte da telerilevamento, indici di diversità strutturale delle foreste e indici relativi alle comunità dei Lepidotteri notturni.
Materiali e metodi
Area di studio
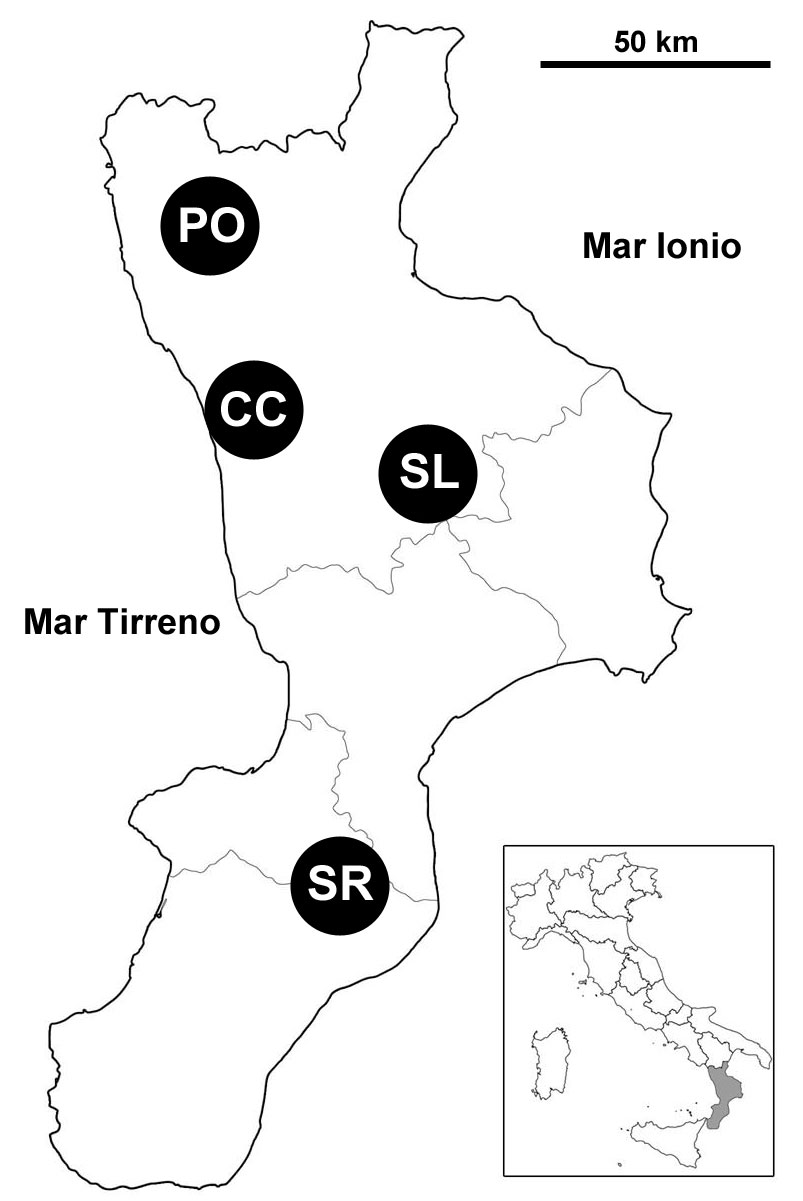
Le attività sono state condotte in formazioni boschive rappresentative dei principali tipi forestali (European Forest Types - [2]) della Regione Calabria, per la precisione: faggete montane a Fagus sylvatica L. (Forest Type 7.3), castagneti a Castanea sativa Mill. (F.T. 8.7), pinete a dominanza di pino nero di Calabria (Pinus nigra calabrica [Loud.] Cesca & Peruzzi - F.T. 10.2) e boschi a dominanza di abete bianco (Abies alba Mill. - F.T. 10.6). I quattro tipi forestali sono stati campionati in altrettanti comprensori montuosi differenti (Fig. 1).
Fig. 1 - Inquadramento geografico delle aree di studio. CC è il codice identificativo dei castagneti della Catena Costiera, PO quello delle faggete del Pollino-Orsomarso, SL quello delle pinete della Sila e SR quello delle abetine delle Serre Vibonesi.
I castagneti sono stati individuati lungo la Catena Costiera, fra 550 e 750 m di quota. Per la maggior parte, i popolamenti in questione per quest’area sono sottoposti a ceduazione con un turno tra 12 e 20 anni e un’estensione del taglio variabile, ma che in alcuni casi può arrivare a 10 ha. Piccole porzioni vengono utilizzate per la produzione di castagne.
Le faggete, campionate nell’area Pollino-Orsomarso, ricadono quasi interamente nella Zona A (protezione integrale) del territorio del Parco Nazionale del Pollino, fra 990 e 1475 m di quota e non sono sottoposti a trattamenti selvicolturali ordinari, per cui si rinvengono per lo più fustaie e cedui avviati a fustaia.
Le pinete di pino nero di Calabria sono molto estese sull’altopiano della Sila. Le aree scelte ricadono quasi tutte nel territorio del Parco Nazionale della Sila, fra 1300 e 1450 m di quota. Anche se sono foreste storicamente sfruttate per la produzione di materiale legnoso e resine, negli ultimi decenni le attività forestali si sono focalizzate prevalentemente alla produzione di biomasse, quindi con tagli a raso su piccole strisce di dimensioni variabili in base alle caratteristiche topografiche. Nel periodo a cavallo degli anni ’ÂÂ70-’80, queste aree sono state interessate da consistenti interventi di rimboschimento a cui, però, spesso non sono seguite le necessarie operazioni selvicolturali.
Le abetine sono i boschi maggiormente rappresentati sui monti delle Serre Vibonesi, sia come boschi monofitici, sia nella classica associazione con il faggio. Le aree di studio rientrano nei territori del Parco Regionale delle Serre, fra 830 e 1090 metri di altitudine. L’industria forestale rappresenta, insieme al turismo, il settore economico più importante del comprensorio ([27]): la pratica selvicolturale tradizionalmente più diffusa è il taglio saltuario, con periodi di curazione piuttosto lunghi.
Campionamento dei Lepidotteri
Le comunità dei Lepidotteri notturni sono state campionate tramite l’utilizzo di trappole luminose a LED con picco di emissione a 395 nm (15 W - [18]).
In ogni complesso forestale sono state individuate 9 stazioni, per un totale di 36 siti scelti in modo da includere vari stadi successionali del bosco. In dettaglio, sono stati inclusi 3 boschi con età superiore a 80 anni, 3 boschi giovani (età compresa tra 20 e 40 anni) e 3 stadi iniziali della successione ecologica (aree in rinnovazione, prati o zone sottoposte a tagli recenti). In ognuno di questi siti è stata collocata una trappola luminosa. Le trappole ricadenti nella stessa area sono state attivate simultaneamente, mentre ogni ciclo di campionamento, comprendente tutte le aree indagate, è stato ultimato nel giro di 10-15 giorni. I trappolamenti hanno avuto cadenza mensile e sono stati effettuati tra maggio e novembre 2015.
Per ogni stazione di campionamento vengono riportati i nomi delle località, i codici alfanumerici delle stazioni, le coordinate UTM, la quota e lo stadio di evoluzione del bosco (Tab. 1).
Tab. 1 - Descrizione sintetica dei siti di campionamento: vengono riportati i codici alfanumerici assegnati a ogni stazione, i nomi delle località, le coordinate UTM, la quota e lo stadio di evoluzione del bosco.
| Cod. Stazione | Località | UTM E | UTM N | Quota (m s.l.m.) |
Stadio Evolutivo |
|---|---|---|---|---|---|
| PO_A1 | Serrapaolo | 593380.28 | 4408629.75 | 990 | Bosco maturo |
| PO_A2 | Serrapaolo | 593145.98 | 4408625.88 | 1010 | Bosco giovane |
| PO_A3 | Serra Ambruna | 592154.60 | 4408713.41 | 1035 | Bosco in rinnovazione |
| PO_B1 | Bocca di Novacco | 589227.26 | 4407585.74 | 1315 | Bosco maturo |
| PO_B2 | Bruscata | 589605.06 | 4407229.84 | 1370 | Bosco giovane |
| PO_B3 | Bocca di Novacco | 589457.29 | 4407641.73 | 1339 | Bosco in rinnovazione |
| PO_C1 | Timpone della Magara | 590076.78 | 4405412.04 | 1465 | Bosco maturo |
| PO_C2 | Timpone della Magara | 589926.23 | 4405142.61 | 1460 | Bosco giovane |
| PO_C3 | Timpone della Magara | 590112.84 | 4404524.50 | 1475 | Bosco in rinnovazione |
| CC_A1 | Fiego di San Fili | 597305.26 | 4354806.35 | 723 | Bosco maturo |
| CC_A2 | Fiego di San Fili | 597338.10 | 4354031.43 | 749 | Bosco giovane |
| CC_A3 | Fiego di San Fili | 597286.88 | 4353884.90 | 734 | Bosco in rinnovazione |
| CC_B1 | Bosco dei Gesuiti | 597450.20 | 4357511.91 | 618 | Bosco maturo |
| CC_B2 | Bosco dei Gesuiti | 597428.66 | 4357562.12 | 626 | Bosco giovane |
| CC_B3 | Mandarino | 597407.18 | 4358601.68 | 583 | Bosco in rinnovazione |
| CC_C1 | Glicarello | 596907.77 | 4362569.92 | 550 | Bosco maturo |
| CC_C2 | Vallone Argentino | 596647.81 | 4362750.45 | 565 | Bosco giovane |
| CC_C3 | Vallone Argentino | 596863.17 | 4362822.96 | 545 | Bosco in rinnovazione |
| SR_A1 | Santa Maria | 614221.47 | 4268211.51 | 847 | Bosco maturo |
| SR_A2 | Il Palmento | 614499.70 | 4269051.60 | 831 | Bosco giovane |
| SR_A3 | Il Palmento | 614593.16 | 4269034.60 | 827 | Bosco in rinnovazione |
| SR_B1 | Cattarinella | 614985.16 | 4267580.80 | 943 | Bosco maturo |
| SR_B2 | Cattarinella | 615080.79 | 4267287.71 | 963 | Bosco giovane |
| SR_B3 | Cattarinella | 615261.05 | 4266275.33 | 1045 | Bosco in rinnovazione |
| SR_C1 | Sietto dello Caricatore | 617070.03 | 4267910.15 | 1086 | Bosco maturo |
| SR_C2 | Pietra del Signore | 616059.05 | 4266558.50 | 1090 | Bosco maturo |
| SR_C3 | Sietto dello Caricatore | 617022.42 | 4267763.47 | 1091 | Bosco maturo |
| SL_A1 | Montagna Grande | 638895.15 | 4348709.71 | 1352 | Bosco maturo |
| SL_A2 | Montagna Grande | 639189.07 | 4349312.45 | 1325 | Bosco giovane |
| SL_A3 | Montagna Grande | 638566.08 | 4348166.12 | 1344 | Bosco in rinnovazione |
| SL_B1 | Torre Scarda | 630869.06 | 4344438.23 | 1324 | Bosco maturo |
| SL_B2 | Quaresima | 625633.51 | 4341424.98 | 1306 | Bosco giovane |
| SL_B3 | Quaresima | 630222.35 | 4343968.61 | 1325 | Bosco in rinnovazione |
| SL_C1 | Colle Macchie | 631744.21 | 4346638.83 | 1428 | Bosco maturo |
| SL_C2 | Vallivone | 632054.06 | 4346721.55 | 1446 | Bosco giovane |
| SL_C3 | Colle Macchie | 631599.43 | 4346653.07 | 1433 | Bosco in rinnovazione |
Dati derivati da telerilevamento
Lo studio ha impiegato dati LiDAR acquisiti nel periodo 2009-2011 con una copertura pressoché continua (wall-to-wall) della superficie regionale della Calabria acquisiti da piattaforma aerea (ALS). Il dato ALS disponibile consiste di un impulso LiDAR a ritorno discreto, con una densità approssimativa di 1.5 impulsi al metro quadrato. I dati sono stati pre-processati per ottenere i rispettivi modelli della superficie (DSM) e del terreno (DTM), che hanno consentito, per sottrazione, di ricavare un modello digitale delle chiome (CHM) esportato in formato raster-matrix a una risoluzione di 1 metro. Tramite l’integrazione tra dati ottenuti dai punti di campionamento a terra rilevati in terza fase dell’INFC e aree realizzate nell’ambito del progetto ALForLAB è stato possibile modellizzare la biomassa epigea in funzione delle metriche LiDAR estratte sulle aree a terra (per maggiori dettagli sulla metodologia, si veda [30]).
Si è poi proceduto a modellizzare la ripartizione della biomassa arborea epigea in tre classi dimensionali: (i) alberi piccoli: diametro a 1.30 m (DBH) < 15 cm, (ii) alberi medi: 15 cm < DBH < 28 cm, (iii) alberi grandi: DBH > 28cm, utilizzando modelli multiregressivi basati sulle seguenti metriche LiDAR del CHM: somma, massimo, media, mediana, deviazione standard. I modelli sono risultati particolarmente accurati per predire la biomassa appartenente alla classe dimensionale degli alberi grandi (Tab. 1). Tramite questo approccio è stato possibile calcolare un indice di equitabilità (distribuzione relativa della biomassa arborea nelle tre classi dimensionali - Tab. 2) utilizzando l’indice di Shannon.
Tab. 2 - Modello di stima della ripartizione della biomassa arborea epigea in classi dimensionali (piccole: diametro a 1.30 m [DBH] < 15 cm; medie: 15 cm < DBH < 28 cm; grandi: DBH > 28 cm).
| Biomassa epigea Classe dimensionale |
metriche del CHM | R2 | ||||
|---|---|---|---|---|---|---|
| somma | massimo | media | mediana | dev. standard | ||
| Piccole | -0.0004 (0.0001) |
0.0897 (0.0410) |
-0.0796 (0.125) |
0.2418 (0.0889) |
-0.1349 (0.0870) |
0.39 |
| Medie | -0.0002 (0.0003) |
0.0205 (0.1316) |
0.1091 (0.4011) |
0.2434 (0.2852) |
0.0482 (0.2781) |
0.56 |
| Grandi | 0.00152 (0.0005) |
-0.8645 (0.2042) |
2.2736 (0.6223) |
-1.5065 (0.4425) |
1.6476 (0.4315) |
0.72 |
Un ulteriore strato informativo derivato da OT è costituito dal dato Landsat 8 (L8) OLI, che è stato utilizzato per il calcolo dell’indice NDVI (Normalized Difference Vegetation Index - [23]).
Gli strati informativi ottenuti da ALS e L8 sono stati confrontati con i dati ottenuti dal campionamento dei Lepidotteri adulti (vedi sotto).
Analisi dei dati
Per caratterizzare le comunità dei Lepidotteri sono stati utilizzati i più comuni indici di ricchezza e diversità specifica (numero di Specie, Shannon, Simpson, Fisher α), calcolati tramite il software EstimateS 9.1.0 ([8]). Inoltre, è stata calcolata la proporzione (rapporto G/N) fra specie e fra individui appartenenti alle famiglie dei Geometridi, la cui diversità aumenta in ecosistemi forestali, e dei Noctuidae, la cui diversità aumenta in ecosistemi dominati da formazioni erbacee. Si assume che più è elevato questo rapporto, ovvero più alta risulta la frazione di Geometridae, maggiore è la naturalità del bosco ([1], [5]).
Gli indici di ricchezza e diversità specifica delle comunità dei Lepidotteri notturni sono stati confrontati con le informazioni derivanti da OT, specificatamente: metriche CHM, biomassa arborea epigea, equitabilità della distribuzione della biomassa da ALS, NDVI da L8. Nota la coordinata delle trappole luminose installate a terra, le metriche OT sono state ottenute estraendole da una area circolare di raggio pari a 20 m centrata sulle trappole. I dati sulle comunità dei Lepidotteri notturni e quelli telerilevati sono stati correlati utilizzando come indice statistico del loro grado di associazione il coefficiente di Pearson (r test).
Risultati
Lepidotteri
In totale sono stati campionati 50.234 individui (N) di Lepidotteri appartenenti a 546 specie (S) così suddivisi tra i diversi tipi forestali: castagneti (N = 8.164; S = 328), faggete (N = 26.558; S = 352), pinete (N = 10.487; S = 306), abetine (N = 5.025; S = 254). Nella Tab. 3 sono riassunti i dati di cattura, gli indici di diversità e il rapporto G/N.
Tab. 3 - Dati di cattura dei Lepidotteri: per ogni stazione sono riportati il numero di specie e di individui catturati, i valori degli indici di diversità più comunemente utilizzati e il rapporto Geometridae/Noctuidae, calcolato sia in termini quantitativi (sul totale degli individui campionati) che in termini qualitativi (sul totale delle specie campionate).
| Stazione | N individui |
N specie |
Fisher α | Shannon | Simpson | G/N Individui |
G/N Specie |
|---|---|---|---|---|---|---|---|
| CC_A1 | 1029 | 150 | 48.42 | 3.86 | 15.13 | 2.56 | 1.08 |
| CC_A2 | 987 | 145 | 46.87 | 3.9 | 19.05 | 2.00 | 1.51 |
| CC_A3 | 507 | 121 | 50.32 | 3.82 | 17.77 | 1.29 | 1.57 |
| CC_B1 | 651 | 143 | 56.62 | 4.17 | 34.22 | 1.73 | 1.07 |
| CC_B2 | 1649 | 193 | 56.7 | 4.37 | 44.24 | 1.41 | 0.87 |
| CC_B3 | 452 | 120 | 53.39 | 3.97 | 26.88 | 1.86 | 1.49 |
| CC_C1 | 665 | 137 | 52.33 | 4.14 | 36.45 | 1.30 | 1.16 |
| CC_C2 | 1601 | 172 | 48.87 | 4.04 | 27.62 | 3.33 | 1.30 |
| CC_C3 | 623 | 118 | 43.1 | 3.76 | 18.45 | 2.31 | 1.30 |
| PO_A1 | 6361 | 204 | 40.24 | 2.72 | 3.64 | 0.94 | 1.01 |
| PO_A2 | 8295 | 226 | 42.88 | 2.34 | 2.64 | 1.31 | 1.04 |
| PO_A3 | 1966 | 166 | 43.25 | 3.16 | 5.96 | 1.48 | 1.08 |
| PO_B1 | 918 | 110 | 32.62 | 3.44 | 15.24 | 2.68 | 0.79 |
| PO_B2 | 2399 | 128 | 28.88 | 3.3 | 15.2 | 2.03 | 0.87 |
| PO_B3 | 602 | 90 | 29.32 | 3.22 | 10.78 | 1.73 | 1.00 |
| PO_C1 | 3491 | 97 | 18.49 | 2.08 | 3.24 | 8.87 | 0.93 |
| PO_C2 | 1842 | 119 | 28.42 | 3.22 | 11.68 | 3.91 | 1.00 |
| PO_C3 | 684 | 121 | 42.69 | 3.91 | 25.77 | 1.09 | 0.96 |
| SL_A1 | 1114 | 147 | 45.36 | 4.03 | 28.25 | 3.21 | 1.34 |
| SL_A2 | 2530 | 157 | 37.04 | 3.33 | 12.88 | 7.49 | 0.93 |
| SL_A3 | 1046 | 141 | 43.9 | 4.01 | 31.52 | 2.38 | 0.89 |
| SL_B1 | 1367 | 139 | 38.69 | 3.72 | 13.29 | 3.78 | 1.51 |
| SL_B2 | 869 | 135 | 44.76 | 4.11 | 36.18 | 1.52 | 0.93 |
| SL_B3 | 247 | 75 | 36.65 | 3.81 | 32.53 | 0.66 | 0.78 |
| SL_C1 | 1119 | 142 | 43.1 | 3.8 | 22.74 | 2.13 | 1.12 |
| SL_C2 | 1790 | 146 | 37.59 | 3.71 | 19.76 | 3.49 | 1.19 |
| SL_C3 | 405 | 83 | 31.61 | 3.55 | 17.93 | 1.15 | 0.81 |
| SR_A1 | 672 | 119 | 42 | 3.71 | 13.78 | 3.31 | 1.69 |
| SR_A2 | 751 | 136 | 48.55 | 4.2 | 40.49 | 3.94 | 1.48 |
| SR_A3 | 155 | 73 | 53.86 | 3.95 | 47.93 | 2.41 | 2.33 |
| SR_B1 | 487 | 98 | 36.96 | 3.94 | 35.59 | 3.61 | 1.60 |
| SR_B2 | 637 | 103 | 34.79 | 3.82 | 25.66 | 3.76 | 1.42 |
| SR_B3 | 221 | 76 | 40.96 | 3.82 | 33.72 | 1.30 | 1.00 |
| SR_C1 | 641 | 93 | 29.89 | 3.58 | 21.38 | 2.06 | 0.74 |
| SR_C2 | 1201 | 113 | 30.57 | 3.62 | 19.2 | 3.15 | 0.98 |
| SR_C3 | 260 | 69 | 30.69 | 3.56 | 22.57 | 3.36 | 1.11 |
La maggiore ricchezza specifica e la maggiore abbondanza di individui sono state registrate nelle faggete del Pollino. A questi dati non corrispondono proporzionali valori degli indici di diversità (Shannon, Simpson, Fisher), che, anzi, sono mediamente i più bassi delle quattro aree. Ciò è dovuto principalmente alla marcata dominanza di poche specie su tutte le altre. Gli indici di diversità hanno mostrato i valori più alti per i siti dominati dal castagno, ma sono piuttosto elevati anche quelli di pinete e abetine, nonostante queste ultime presentino di gran lunga la minore quantità di individui e la minore ricchezza specifica, frutto di una marcata equiripartizione delle abbondanze relative. Le diverse comunità entomiche sono ben caratterizzate e molto differenziate tra loro. Considerando le cinque specie più abbondanti di ogni biocenosi (Tab. 4) non ve ne sono in comune fra i vari tipi forestali, ad eccezione di Alcis repandata, specie caratterizzante sia i boschi a dominanza di pino nero sia le formazioni a dominanza di abete bianco.
Tab. 4 - Specie più abbondanti di Lepidotteri per tipo forestale analizzato. (*): Unica specie presente in più tipi forestali.
| Castagneti | Faggete |
|---|---|
| Peribatodes rhomboidaria | Eilema lurideola |
| Clemathada calberlai | Operophtera fagata |
| Paracolax tristalis | Campaea margaritata |
| Dysauxes famula | Epirrita christyi |
| Eilema caniola | Cosmia trapezina |
| Pinete | Abetine |
| Alcis repandata* | Macaria liturata |
| Pennithera firmata | Pungeleria capreolaria |
| Idaea deversaria | Lithosia quadra |
| Thaumetopoea pityocampa | Alcis repandata* |
| Epirrhoe galiata | Dendrolimus pini |
Il rapporto G/N relativo alle specie ha mostrato valori superiori a 1 (maggiore numero di Geometridae rispetto ai Noctuidae) nella maggior parte dei siti di campionamento, raggiungendo generalmente i valori massimi nei boschi di maggiore età.
Dati telerilevati e analisi di correlazione
I dati ottenuti da OT e quelli relativi allo studio delle comunità di Lepidotteri sono stati sottoposti ad analisi di correlazione, i cui risultati sono illustrati in Tab. 5.
Tab. 5 - Risultato dell’analisi di correlazione tra indici di diversità e dati misurati a terra (Pearson r test); (Ps1_median): mediana della biomassa del fusto calcolata nell’area di raggio pari a 20 m centrata sulle trappole dei lepidotteri; (Ps2_median): mediana della biomassa epigea calcolata nell’area di raggio pari a 20 m; (SH_BIOM): indice di equitabilità della distribuzione della biomassa epigea in classi dimensionali; (CHM_x): metriche del CHM estratte nell’area di raggio pari a 20 m centrata sulle trappole dei lepidotteri; (*): p<0.05; (**): p<0.01; (***): p<0.001; (ns): non significativo (p>0.05).
| Parametro | N individui |
N specie |
Fisher a | Shannon | Simpson | G/N Individui |
G/N Specie |
SH_BIOM |
|---|---|---|---|---|---|---|---|---|
| Quota | ns | ns | *** | * | * | ns | ** | ns |
| Ps1_median | ns | ns | *** | ns | ns | * | ns | *** |
| Ps2_median | ns | ns | *** | ns | ns | * | ns | *** |
| NDVI_median_L8 | ** | ** | ns | * | ns | ns | ns | ns |
| CHM_sum | ns | ns | * | ns | ns | ** | ns | *** |
| CHM_mean | ns | ns | * | ns | ns | ** | ns | *** |
| CHM_median | ns | ns | ns | ns | ns | ** | ns | *** |
| CHM_stdev | ns | ns | ns | ns | ns | ns | ns | *** |
| CHM_min | ns | ns | ns | ns | ns | ns | ns | ns |
| CHM_max | ns | ns | * | ns | ns | * | ns | *** |
| SH_BIOM | ns | ns | ns | ns | ns | ** | ns | - |
Il risultato più significativo riguarda il rapporto G/N, positivamente correlato alla biomassa arborea epigea e negativamente correlato alla sua equitabilità: l’aumento di massa arborea favorisce i Geometridae, generalmente più legati a tali ambienti, mentre la dominanza di una sola classe dimensionale arborea sulle altre fa diminuire il rapporto a vantaggio dei Noctuidae. La biomassa è, inoltre, negativamente correlata con il Fisher α, indice di diversità frequentemente utilizzato per i Lepidotteri. Inoltre, l’indice G/N tende a diminuire con la quota e ad aumentare con la copertura (approssimata dall’NDVI). A quest’ultimo indice di copertura è positivamente correlato il numero di Lepidotteri in termini specifici, mentre è inversamente correlato l’indice di Shannon. La quota influisce negativamente su tutti gli indici di diversità relativi ai Lepidotteri.
Discussione
I Lepidotteri possono fornire preziose indicazioni sul grado di diversità degli ambienti forestali, soprattutto in caso di habitat frammentati o in rinnovazione perché strettamente associati alla distribuzione delle loro piante nutrici ([17]). A volte, però, può risultare complicato reperire informazioni dettagliate sulla dieta larvale delle singole specie, mentre maggiori sono le conoscenze sulle preferenze ecologiche. I diversi tipi forestali esaminati in questo studio sono caratterizzati da comunità di Lepidotteri peculiari, il cui studio completo offrirebbe numerose chiavi interpretative delle complesse interazioni ecologiche. Ciononostante, è possibile individuare alcune caratteristiche della struttura nelle biocenosi che possono dare indicazioni abbastanza attendibili sulla naturalità degli ecosistemi e che prescindono dalla specie arborea dominante ([21]). Di fatto, i risultati ottenuti in questo studio mostrano una stretta rispondenza tra la struttura delle comunità di Lepidotteri e le caratteristiche dei popolamenti forestali.
Particolarmente indicativo è il rapporto G/N. Un rapporto G/N a favore dei Geometridae è considerato indice di maggiore naturalità per gli ambienti forestali ([32]) e ciò è confermato dai risultati delle analisi effettuate che mostrano una correlazione con la quantità di biomassa e con la copertura arborea. La correlazione risulta invece negativa nel caso in cui una classe dimensionale sia dominante nel bosco: l’assenza di una struttura forestale complessa, infatti, rappresenta una condizione spesso ascrivibile a una gestione selvicolturale che tende a semplificare le cenosi con la creazione di sistemi comparativamente meno favorevoli alla biodiversità.
In termini di valutazione dello stato di conservazione delle foreste, il rapporto G/N, fornisce indicazioni più attendibili rispetto agli indici di diversità, a volte ritenuti poco affidabili nel caso di ambienti frammentati ([4], [28]). Questo rapporto, infatti, è il risultato di una valutazione, seppure semplificata, della struttura di comunità che prende in considerazione anche le caratteristiche autoecologiche dei taxa.
Anche NDVI si dimostra un indice di un certo interesse: correlazioni tra questo indice e altre variabili ecologiche hanno fornito risultati significativi in passato, anche riguardo alla componente animale e le relazioni tra cambiamenti climatici e dinamica delle popolazioni ([7], [25]). La correlazione positiva di NDVI con il rapporto G/N e con la ricchezza di specie di Lepidotteri conferma l’utilità di questo dato per la mappatura della naturalità forestale.
Nel complesso, i risultati sono da ritenersi particolarmente significativi in quanto riferiti a quattro tipi forestali differenti e ciò sembrerebbe suggerire la possibilità di generalizzare le conclusioni raggiunte, almeno nelle condizioni ambientali (montano-mediterraneee) considerate.
Ringraziamenti
Si ringraziano Carlo Di Marco, Franco Calabrese, Gino Scarpelli, Massimo ed Ettore Salerno per il supporto tecnico. Il lavoro è stato finanziato dal Progetto “ALForLab” (PON03PE_00024_1) cofinanziato dal Programma Operativo Nazionale Ricerca e Competitività (PON R&C) 2007-2013, attraverso il Fondo Europeo di Sviluppo Regionale (FESR) e risorse nazionali (Fondo di Rotazione (FDR) - Piano di Azione e Coesione (PAC) MIUR).
References
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar