What role for afforestation in Italian strategies towards the Kyoto Protocol? Hints from a Kyoto forest in the Po Valley (Northern Italy)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 2, Pages 333-344 (2005)
doi: https://doi.org/10.3832/efor0320-0020333
Published: Dec 19, 2005 - Copyright © 2005 SISEF
Research Articles
Abstract
The carbon balance of an afforested area in the Emilia-Romagna floodplain has been monitored for a 4-year period. Detailed measurements covering both above- and below-ground components of biomass production and accumulation were complemented by eddy-covariance measurements at the ecosystem level. Experimental results have been extrapolated in time by means of a process-based ecosystem model, so as to better assess the potential role of afforestation towards the carbon-reduction goals of the Kyoto Protocol.
Keywords
Introduzione
Le foreste dei climi temperati presentano nel loro complesso un bilancio del carbonio positivo, accumulando attraverso i processi fotosintetici più CO2 di quanta ne venga rilasciata dalla respirazione delle piante e dei suoli ([9]). Tale capacità di fissazione è dovuta in primo luogo alla progressiva espansione naturale della superficie coperta da foreste, a causa della riduzione delle aree dedicate all’agricoltura, ed alla riduzione delle utilizzazioni forestali, attraverso l’allungamento dei turni forestali e la sospensione di pratiche quali la raccolta di lettiera e di legno morto. A questa espansione naturale vanno sommati peraltro gli effetti degli interventi di afforestazione realizzati nel corso degli anni, spesso con finanziamenti pubblici. Questi sistemi, partendo da una situazione di scarso accumulo di carbonio, stanno ovviamente immagazzinando attivamente carbonio in tutte le loro componenti ([19]). Benché una gran parte degli zuccheri resi disponibili dalla fotosintesi vengano rilasciati dalla respirazione, in sistemi giovani quali gli impianti di arboricoltura il bilancio netto (NEP, net ecosystem production: quantità di carbonio complessivamente sottratta all’atmosfera dall’ecosistema) risulta normalmente positivo.
In considerazione di questo importante contributo, il Protocollo di Kyoto, con cui l’Italia si è impegnata ad una riduzione entro il periodo 2008-2012 del 6.5 % delle emissioni di carbonio rispetto al valore registrato nel 1990, prevede all’Art.3.3 la possibilità per i Paesi firmatari di controbilanciare le emissioni antropiche con attività di riforestazione (nel caso di ricostituzione della foresta in aree recentemente deforestate, ma comunque prima del 1990) o di afforestazione (nel caso di interventi su terreni non precedentemente forestati da almeno 50 anni) che risultino nella immobilizzazione della CO2 nell’ecosistema forestale, sotto forma di biomassa vegetale o di sostanza organica del suolo. Verranno a tal fine presi in considerazione solo interventi di forestazione avviati dopo la data di riferimento del 1990. A questi boschi di neoformazione (categoria che comprende anche gli impianti di arboricoltura specializzata da legno) è stato pertanto attribuito il nome di Kyoto forests.
È stato stimato che gli interventi di riforestazione realizzati nel decennio 1990-2000 usufruendo di finanziamenti pubblici (primi fra tutti quelli previsti dal Reg. CEE 2080/92) abbiano interessato oltre 148000 ha sull’intero territorio nazionale ([1]). Di questi, gli impianti di arboricoltura da legno con latifoglie di pregio coprirebbero da soli oltre 87000 ha, contro un totale di circa 61000 ha fra pioppeti, eucalitteti ed impianti di conifere.
Il ruolo che gli interventi di afforestazione potrebbero svolgere nella riduzione delle emissioni totali non è da trascurare. A livello europeo, si stima che fra il 1990 ed il 2000 l’estensione delle aree rimboschite sia stata pari a circa 170000 ha / anno; se mantenuta, questa tendenza potrebbe portare in combinazione con l’espansione naturale del bosco ad una fissazione di 14 Mt CO2 anno-1 nel periodo di riferimento 2008-2012 ([7]).
Una stima ancora più alta si evince dalle intenzioni espresse dagli Stati membri, secondo cui gli interventi di riforestazione fra il 1990 ed il 2008-2012 dovrebbero contribuire con la rimozione di 23 milioni di tonnellate di CO2 all’anno ([8]), equivalenti ad oltre l’8 % della riduzione dovuta complessivamente dall’Europa a 15.
Per quanto riguarda l’Italia, il Ministero dell’Ambiente prevede un contributo positivo di 10.2 Mt CO2 all’anno dalle attività legate a cambiamenti di uso del suolo e gestione forestale ([14]). Di questi, si assume che 1 Mt CO2 all’anno possa costituire il contributo degli interventi di afforestazione e riforestazione già realizzati a partire dal 1990, mentre altri 2 Mt CO2 anno-1 dovrebbero venire da impianti di nuova realizzazione, equamente distribuiti fra interventi di arboricoltura ed interventi di riforestazione su aree soggette a dissesto idrogeologico, previsti dalla Legge 183/89. La naturale espansione del bosco resa possibile dalla contrazione dell’agricoltura montana contribuirebbe poi per 3 Mt CO2 anno-1, mentre altri 4.1 Mt CO2 anno-1 deriverebbero dalla gestione forestale conservativa realizzata in gran parte dei boschi italiani, grazie principalmente all’allungamento dei turni di utilizzazione ed all’avviamento all’alto fusto di parte dei cedui. Il contributo di terre agricole e pascoli ammonterebbe a non più di 0.1 Mt CO2 anno-1.
In uno studio commissionato dall’APAT, d’altra parte, Anderle et al. ([1]) hanno stimato in sole 0.63 Mt CO2 anno-1 il contributo di tutte le piantagioni forestali realizzate nel periodo 1990-2000, ben al di sotto quindi delle proiezioni formulate dal Ministero. Gli Autori, tuttavia, hanno incluso nei loro calcoli solo il carbonio contenuto nei fusti da lavoro delle piante forestali, che costituisce solo una delle componenti del carbonio fissato nel complesso dall’ecosistema. Questa scelta, motivata in parte dalla necessità di un approccio prudenziale e precisamente quantificabile, era anche motivata dalla mancanza di dati precisi sul bilancio del carbonio di simili Kyoto forests su suoli ex-agricoli negli ambienti italiani.
La presente ricerca si è proposta di colmare queste lacune, per giungere ad una più affidabile valutazione del contributo degli impianti di arboricoltura da legno in Italia rispetto agli obiettivi prefissi dal Protocollo di Kyoto.
A tal fine il bilancio del carbonio di un rimboschimento realizzato con finanziamenti pubblici nella pianura emiliana è stato monitorato per un periodo di 5 anni. Le misure sperimentali di dettaglio hanno riguardato non solo la componente aerea, ma anche le varie componenti ipogee e sono state complementate da misure integrate a livello di intero popolamento. I dati sperimentali sono stati infine estrapolati nel tempo con l’aiuto di un modello matematico a base funzionale, così da meglio stimare quale possa essere il contributo dell’arboricoltura da legno nel suo complesso al raggiungimento degli obiettivi del Protocollo di Kyoto.
Materiali e metodi
Descrizione del sito
La ricerca è stata condotta in un rimboschimento misto realizzato con finalità naturalistiche nell’autunno del 1992 nel Comune di Nonantola (MO), su terreni appartenenti alla Partecipanza Agraria di Nonantola. I terreni interessati dal rimboschimento, soggetti ad allagamenti periodici, erano stati storicamente occupati dal "bosco di Nonantola", documentato fin dal XVI secolo, prima di essere convertiti all’agricoltura alla fine dell’800. Da allora e fino all’anno precedente all’impianto, i terreni erano stati utilizzati per la coltura di frumento, erba medica e barbabietola.
L’impianto costituisce quindi un esempio di Kyoto forest, rispondendo ai criteri imposti dall’Art. 3.3 del Protocollo di Kyoto per l’ammissibilità al conteggio del bilancio del carbonio, ed è stato per questo incluso fra i siti sperimentali del progetto europeo Carbo-Euroflux, il quale si propone di quantificare il bilancio del carbonio della vegetazione europea.
Il bosco è stato piantato su suoli molto profondi, tipici di aree morfologicamente depresse della pianura alluvionale, classificati dalla cartografia tematica regionale come serie regionale o sotto-unità "Risaia del Duca" RSD1 ([20]) e rientranti secondo la Soil Taxonomy della USDA nella classe dei Fine, Mixed, Mesic Entic Chromusterts ([26]). In particolare, il sito oggetto di studio è caratterizzato da un tenore in argilla molto alto, con valori intorno al 60 % ([16]).
Il clima è caratterizzato da forti escursioni termiche, precipitazioni primaverili ed autunnali ed una marcata aridità estiva. Nel periodo autunno-primaverile frequenti sono le nebbie, associate a condizioni anticicloniche. Le precipitazioni medie annue sono di 700 mm; la temperatura media annua è di 13.7 °C.
L’impianto misto di Nonantola è stato realizzato nell’autunno 1992, usufruendo di contributi comunitari (Reg. CEE 797/85 - Set aside). L’impianto ha previsto la messa a dimora di circa 40000 semenzali a radice nuda di 1-2 anni di età su una superficie complessiva di 38.6 ha, con un sesto di impianto di 3 x 3 m, disposizione a quadrato e distribuzione casuale delle diverse specie. La messa a dimora dei semenzali di 1-2 anni di etàè stata preceduta da una rippatura alla profondità di 100 cm, seguita da un affinamento superficiale del terreno. Viste le finalità naturalistiche dell’impianto, non è stato realizzato nessun intervento di fertilizzazione, irrigazione o diserbo.
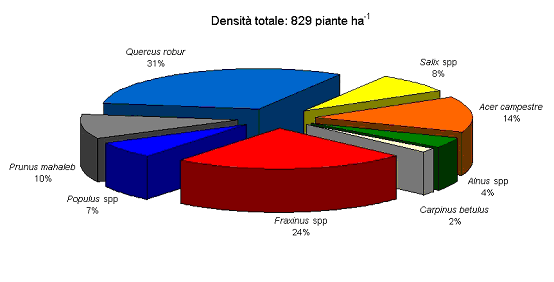
A causa di diverse fallanze e nonostante l’assenza di una reale competizione per la luce, la densità si era ridotta a 829 piante ad ettaro al termine del 1999. La distribuzione fra le diverse specie è riportata in Fig. 1. Prevalgono la farnia (Quercus robur L.) ed i frassini (frassino meridionale, Fraxinus angustifolia L., e frassino maggiore, F. excelsior L.), con una forte compresenza di acero campestre (Acer campestre L.), salici (in particolare salice bianco, Salix alba L.), pioppi (pioppo bianco, Populus alba L. e pioppo nero, P. nigra L.) e magaleppo (Prunus mahaleb L.) e presenze minori di carpino bianco (Carpinus betulus L.) e ontano nero (Alnus glutinosa L.).
Fig. 1 - Contributo delle diverse specie alla densità complessiva dell’impianto misto di arboricoltura di Nonantola, come risultante dai rilievi condotti al termine del 1999. La densità iniziale di impianto stimata era di 1111 piante ad ettaro.
Stima del bilancio di carbonio dell’ecosistema
La capacità del rimboschimento di Nonantola di sottrarre carbonio dall’atmosfera è stata quantificata con due tecniche indipendenti, aumentando così il grado di confidenza nei risultati: dalle variazioni nel carbonio immagazzinato nelle diverse componenti del sistema (metodo inventariale) e da misure dirette di eddy-covariance (metodo micro-meteorologico).
Lo scambio netto dell’ecosistema è stato misurato con la tecnica dell’eddy-covariance ([30], [3]) a partire dal febbraio 2001 utilizzando un analizzatore a risposta veloce Li-6262 (LiCor, Lincoln, NE, USA) ed un anemometro ultrasonico tridimensionale R2 (Gill Instr., Limington, Hampshire, UK) collocato ad un’altezza di 13 m; il dato è stato quindi corretto per l’accumulo di CO2 all’interno della copertura. Tutti i principali parametri micro-ambientali (radiazione, temperatura, umidità dell’aria e del suolo) sono stati anche misurati in continuo in corrispondenza della stazione di misura dei flussi.
A partire dal, 1999 lo sviluppo nel tempo della biomassa epigea è stato determinato attraverso misure invernali di diametro ed altezza su un’ampia area di saggio di 0.6 ha nell’intorno della torre di misura, così da rappresentare tanto la diversità specifica quanto la densità disomogenea del popolamento. Il volume dendrometrico degli individui di ogni specie è stato stimato utilizzando tavole di cubatura locali specifiche per le specie oggetto di studio ([21]). Il volume legnoso complessivo è stato infine stimato utilizzando fattori di conversione (BEF, biomass expansion factors) specifici: per i rami ci si è basati su dati di letteratura relativi a piante di dimensioni similari ([4]), mentre per le radici grossolane si è fatto riferimento a misure dirette su un ampio campione di specie (Grace J., com. pers.). La densità del legno e la sua variazione con l’età cambiale sono state rilevate direttamente su carote legnose estratte a 1.30 m da un campione di 24 piante rappresentative delle quattro principali specie.
La produzione annuale di foglie è stata misurata in queste specie decidue dalle perdite autunnali di lettiera, catturata con 9 trappole di 1 m2 di superficie negli anni 2000 e 2001; negli anni seguenti, la produzione annua di biomassa fogliare è stata stimata attraverso misure periodiche combinate di indice di area fogliare (LAI, leaf area index) ottenute con metodo ottico (LAI-2000, LiCor, Lincoln, NE, USA) e di area fogliare specifica, opportunamente calibrate contro le misure dirette nel corso dei primi due anni.
La biomassa delle radici fini è stata misura direttamente nel corso degli anni 2001-2002 attraverso il prelievo periodico di carote di suolo fino ad una profondità di 40 cm ([17]). Nascita e mortalità delle radici fini stesse sono state determinate nel corso dello stesso periodo con la tecnica del mini-rizotrone ([17]); la calibrazione di queste misure lineari contro le misure distruttive di biomassa radicale sopra descritte ha infine permesso di stimare produttività ipogea e produzione di lettiera radicale nel corso della stagione.
La quantità di carbonio contenuta nella sostanza organica del suolo (SOM, soil organic matter) fino ad una profondità di 30 cm è stata determinata con un analizzatore elementare (EA 1110 Carlo Erba, Italia) su campioni estratti nel corso del 2003. L’accumulo di sostanza organica nel suolo è generalmente lento e molto variabile nello spazio, rendendo difficile una sua quantificazione da misure ripetute a distanza di pochi anni. Si è pertanto assunto che l’accumulo di carbonio nel suolo sia stato uniforme nel corso della vita del popolamento, e che la concentrazione di carbonio al momento dell’impianto fosse pari a quella oggi misurata negli orizzonti più profondi del suolo forestale. Il rapporto fra contenuto di carbonio e peso della sostanza secca è stato determinato direttamente in tutte le componenti del sistema.
A partire da questi dati è stato quindi possibile stimare non solo la distribuzione del carbonio all’interno del sistema, ma anche la produzione netta dell’ecosistema (NEP) e le sue diverse componenti, consistenti nell’accumulo annuo netto di carbonio nei singoli compartimenti dell’ecosistema (foglie, biomassa legnosa, radici fini, suolo).
Dinamiche dell’accumulo di C: applicazione del modello 3PG-3
I modelli di processo sono rappresentazioni semplificate della struttura e delle dinamiche di sistemi complessi quali quelli ecologici. Partendo da conoscenze di dettaglio relative ai singoli processi ed alle mutue interrelazioni fra i componenti dell’ecosistema, essi cercano di spiegarne le proprietà emergenti, permettendo di estrapolare nel tempo le dinamiche osservate. L’applicazione di simili strumenti risulta essenziale per la previsione delle dinamiche future di accumulo di carbonio degli impianti di arboricoltura, non disponendo di impianti maturi su cui sia possibile effettuare misure dirette.
Il modello 3PG-3 è stato sviluppato combinando insieme i modelli 3PG ([13]) ed ICBM (Andrén & Kätterer 1997) al fine di esplorare le potenzialità di fissazione di carbonio in boschi ed impianti di arboricoltura da legno e gli effetti di lungo termine della gestione selvicolturale. Il modello rappresenta l’immobilizzazione di carbonio nel sistema forestale nel suo complesso, includendo cioè non solo la componente aerea ma anche il ruolo del sottobosco e la capacità di immobilizzazione dei suoli forestali. La struttura generale del modello è illustrata in Fig. 2. La rappresentazione della produttività primaria netta del soprassuolo principale e del sottobosco è basata sul concetto di efficienza di uso della luce intercettata, così come adottato nel modello 3PG ([13]). Come in 3PG, tanto la fotosintesi quanto la allocazione alle radici sono funzione dell’età, contribuendo così entrambe a spiegare il decremento di produttività con l’età generalmente osservato nei sistemi forestali. La funzionalità del sottobosco è rappresentata con lo stesso approccio, assumendo però che il carbonio assorbito da questa componente venga prontamente rilasciato al suolo senza alcun accumulo netto. La assunzione appare appropriata per le specie erbacee presenti nel sottobosco di Nonantola.
Fig. 2 - Diagramma schematico del modello 3PG-3, con l’indicazione dei principali flussi e serbatoi di carbonio nell’ecosistema. Il modulo relativo ai prodotti legnosi è stato escluso dalla presente applicazione
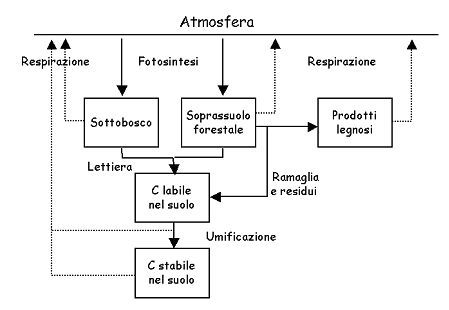
Una frazione costante del carbonio fissato viene rilasciata nell’atmosfera dalla respirazione delle piante stesse ([32]), mentre il rimanente viene accumulato nella biomassa forestale o rilasciato al suolo sotto forma di lettiera (fogliare e radicale) e di essudati. La lettiera e gli essudati radicali vengono decomposti dai microrganismi del suolo e rilasciati come CO2 o trasformati, attraverso i processi di umificazione, in una frazione più stabile, che pure si decompone anche se con dinamiche molto più lente. Le dinamiche della sostanza organica del suolo sono rappresentate invece come nel modello ICBM ([2]).
Al termine del turno forestale, una frazione della biomassa accumulata nelle piante viene rilasciata sotto forma di ramaglia e residui (scarti, apparati radicali, ecc.), mentre gran parte viene trasformata in prodotti legnosi. Questi, a loro volta, saranno caratterizzati da un determinato ciclo di vita al termine del quale sono destinati a decomporsi, rilasciando nell’atmosfera il carbonio in essi contenuto. Quest’ultima componente è stata esclusa nell’ambito della presente applicazione.
Il modello è stato calibrato sulla totalità dei dati disponibili, tanto di flusso quanto di stock di carbonio nelle diverse componenti dell’ecosistema (n = 23), in modo da aumentare al massimo la confidenza nelle stime. Grazie alla sua relativa semplicità, il modello richiede un numero estremamente ridotto di parametri (n = 7), limitando i problemi di covarianza e rendendo quindi possibile la sua calibrazione sui dati con un grado soddisfacente di confidenza.
Risultati
Le ampie dimensioni del campione analizzato permettono di avere una quantificazione precisa degli incrementi delle singole specie all’interno del popolamento, oltre che dell’impianto nel suo complesso.
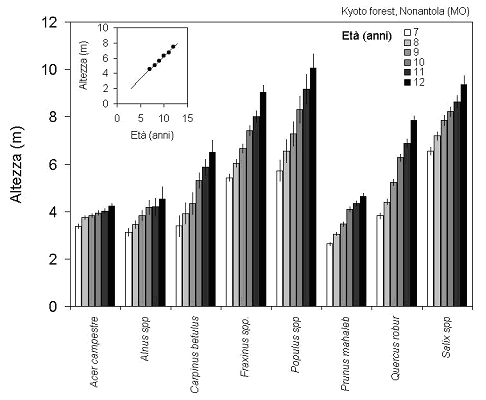
L’altezza media delle piante superava i 7 m all’età di 12 anni (Fig. 3), con un incremento medio piuttosto sostenuto di 60 cm anno-1. Il confronto fra le diverse specie mostra peraltro importanti differenze, con altezze medie comprese fra i 9-10 m di pioppi e salici ed i 4-5 m di aceri (particolarmente soggetti a rottura del cimale e scosciamenti), ontani e magaleppi. Le due specie più rappresentate nel popolamento mostrano pure altezze piuttosto diverse fra loro: più di 9 m per il frassino, meno di 8 m per la farnia. Il diametro medio per l’intero popolamento risultava di 5.5 cm nel 1999 e di 9.8 cm nel 2004.
Fig. 3 - Sviluppo dell’altezza delle diverse specie e del popolamento nel suo complesso nell’impianto misto di Nonantola negli anni 1999-2004. Media ± errore standard.
Nonostante le maggiori dimensioni di pioppi e salici, a livello di intero popolamento sono frassini e querce a dominare le dinamiche di accumulo di carbonio, a causa della loro maggiore incidenza numerica (Fig. 4).
Fig. 4 - Sviluppo dell’incremento corrente di volume totale delle diverse specie e del popolamento nel suo complesso nell’impianto misto di Nonantola negli anni 1999-2004. Si notino gli effetti della siccità del 2003 sul soprassuolo di 11 anni di età.
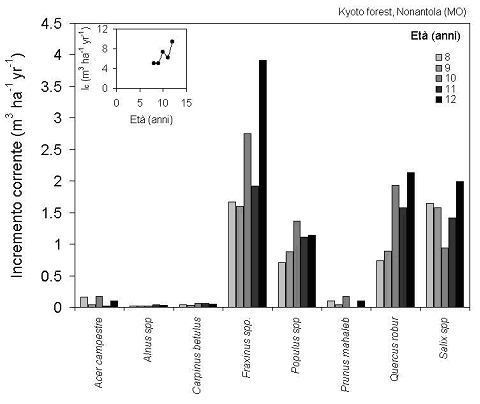
Con oltre 7 m3 ha-1 anno-1 di incremento medio di massa dendrometrica (Fig. 4), l’impianto di Nonantola è caratterizzato da una produttività piuttosto bassa ma ancora in aumento, con un aumento esponenziale della provvigione (vedi inserto in Fig. 5) legato in primo luogo alla chiusura delle chiome ed al conseguente aumento della frazione di luce intercettata. Si è osservato infatti nel periodo coperto dal presente studio un aumento lineare dell’indice di area fogliare (LAI), con valori compresi fra 1.9 ad 8 anni e 2.8 a 12 anni di età. Si tratta comunque di valori abbastanza bassi, tipici di specie eliofile quali frassino e querce, cui corrispondono valori di irradianza relativa al suolo intorno al 38 e al 24%, rispettivamente, che permettono lo sviluppo di uno strato erbaceo pressoché continuo.
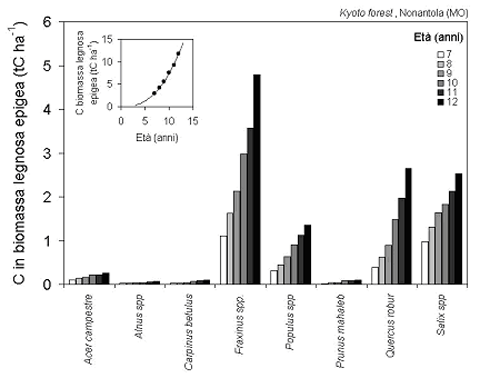
Fig. 5 - Sviluppo del contenuto totale di carbonio nella biomassa legnosa epigea delle diverse specie e del popolamento nel suo complesso nell’impianto misto di Nonantola negli anni 1999-2004.
Di particolare interesse è anche notare gli effetti sugli incrementi della variabilità climatica interannuale, con crescite fortemente ridotte nel 2001 e soprattutto nel 2003 (corrispondenti ad un’età del popolamento di 9 e 11 anni) a causa della marcata aridità estiva. Vale anche la pena di sottolineare come gli incrementi di pioppi e salici, specie tipicamente pioniere, mostrino già una culminazione (Fig. 4).
Il contributo dominante di frassini e querce nelle dinamiche del popolamento risulta ancora più evidente quando queste vengano espresse non in termini di volume ma di carbonio immagazzinato in fusti e rami (Fig. 5), a causa della maggiore densità del legno di queste due specie (dati non presentati). Nel complesso, il bosco di Nonantola racchiude nella sola biomassa legnosa epigea quasi 12 tC ha-1; il tasso di accumulo annuo in questa sola componente ha sfiorato nel 2004 le 2.6 tC ha-1 anno-1, con un valore medio di 1.8 tC ha-1 anno-1 nel periodo coperto dalle misure.
Nello stesso periodo, lo scambio netto dell’ecosistema misurato con la tecnica dell’eddy-covariance è ammontato mediamente a 3.4 tC ha-1 anno-1 (Fig. 6), con un valore massimo di 3.9 tC ha-1 anno-1 nel 2002. Il valore di di 3.7 tC ha-1 anno-1 rilevato nell’anno 2001 è in ottimo accordo con la NEP calcolata per lo stesso anno con metodo inventariale, pari a 3.9 tC ha-1 anno-1. Il confronto con i dati di accumulo nella biomassa legnosa epigea sopra riportati dimostra come mediamente quest’ultima spieghi solo il 57% dell’accumulo netto nell’ecosistema. Tale valore sale al 59% quando si consideri anche la biomassa fogliare.
Fig. 6 - Confronto della produttività netta dell’ecosistema a Nonantola con i dati rilevati in tutta Europa nell’ambito del progetto europeo Carbo-Euroflux ([28]). In nero sono riportati i valori relativi a foreste, in grigio quelli relativi ad impianti artificiali intensivi. In rosso il sito di Nonantola.
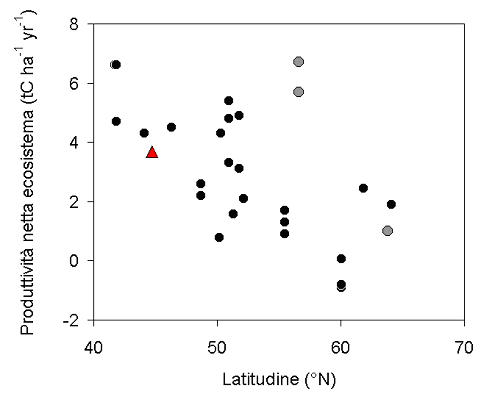
Ancora maggiore è il peso della componente radicale e della sostanza organica del suolo quando si consideri la distribuzione degli stock di C fra le diverse componenti. La sostanza organica nei primi 30 cm di suolo da sola comprende infatti oltre l’85% del carbonio totale presente nell’ecosistema (54.8 tC ha-1). Considerando la sola biomassa, la componente ipogea (radici fini e grossolane) costituisce il 38% del carbonio presente nelle piante, contro un 61% per la biomassa legnosa epigea. Il contributo della biomassa legnosa sul carbonio totale presente nel sistema, d’altra parte, aumenta ovviamente con il maturare delle piante e l’accumulo di provvigione e si stima che all’età di 12 anni si aggirasse intorno al 15%, contro un contributo del 7% all’età di 8 anni.
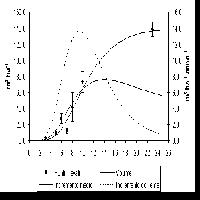
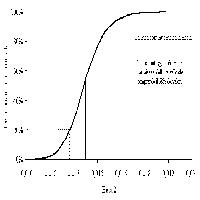
L’accumulo medio di carbonio nell’ecosistema nei primi 9 anni di vita dell’impianto, derivato dalle stime inventariali, è risultato pari a 1.7 t C ha-1 anno-1. Questo valore piuttosto basso è dovuto in primo luogo alla scarsa produttività primaria nella piantagione appena costituita, che nei primi anni è stata anzi presumibilmente caratterizzata da un rilascio netto di carbonio verso l’atmosfera (Fig. 7).
Fig. 7 - Dinamiche attese dello scambio netto dell’ecosistema (NEE) e delle sue componenti epigea ed ipogea nella Kyoto forest di Nonantola, sulla base dell’applicazione del modello 3PG-3.
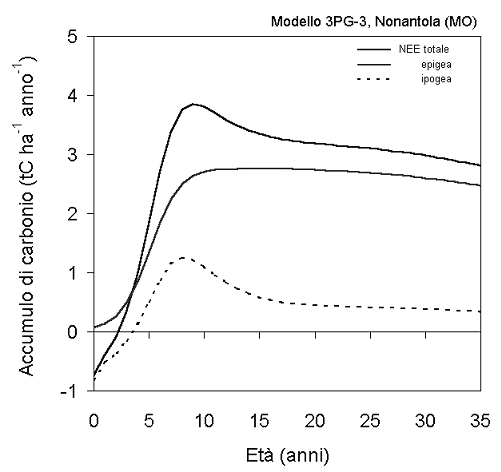
Il forte effetto dell’età sulla entità dell’accumulo di carbonio dimostra l’importanza di integrare ed estrapolare le misure attraverso la applicazione di un modello di ecosistema opportunamente calibrato sui dati. La applicazione del modello 3PG-3 ha permesso di prevedere le dinamiche future di accumulo del carbonio al sito di Nonantola (Fig. 7). L’analisi ha evidenziato come i valori di scambio netto dell’ecosistema rilevati all’età di 9-11 anni corrispondano al picco massimo atteso, a cui seguirebbe negli anni successivi un rapido decremento di circa il 20%. Questo andamento sarebbe da attribuirsi alla rapida immobilizzazione di carbonio nelle radici e nel suolo nella fase giovanile di sviluppo dell’ecosistema, destinata a decrescere col passare del tempo. A questa fase di rapido assestamento farebbe poi seguito un più lento declino dello scambio netto dell’ecosistema, legato alla riduzione con l’età delle potenzialità fotosintetiche ed associato quindi ad una riduzione anche dell’accumulo di carbonio nei fusti. In accordo con le curve di crescita, tale decremento risulta più tardivo e graduale.
Discussione
La ricerca condotta a partire dal 1999 nel rimboschimento di Nonantola costituisce uno dei pochi studi esistenti in Italia sul bilancio del carbonio di un giovane rimboschimento, rispondente alle specifiche previste dall’Art. 3.3 del Protocollo di Kyoto e classificabile quindi come una "Kyoto forest ", atta a controbilanciare le emissioni antropogeniche di gas-serra nell’atmosfera. I numerosi studi di dettaglio condotti nel corso degli ultimi anni nel sito di Nonantola ([5], [16], [22], [17], [11], [12], [10]) hanno permesso di espandere l’analisi dalla dimensione puramente produttiva e forestale ad una più ecologica ed ambientale, quantificando non solo la crescita epigea, ma anche la capacità di immobilizzazione del carbonio atmosferico in tutte le componenti dell’ecosistema, oltre che la sua risposta alla variabilità ambientale interannuale.
Stante la recente entrata in vigore del Protocollo di Kyoto ed i pesanti impegni di riduzione del bilancio totale del carbonio che esso prevede per l’Italia entro il periodo di riferimento 2008-2012, è stato proposto che interventi di rimboschimento di questo tipo possano svolgere nel prossimo futuro un ruolo di notevole importanza.
Come già menzionato, si prevede che in Italia i soli impianti di arboricoltura da legno realizzati con finanziamenti 2080/92 nel decennio 1990-2000 possano assorbire nel periodo di riferimento 2008-2012 all’incirca 1 Mt CO2 all’anno ([14]), mentre una uguale quantità dovrebbe essere assorbita da 40000 ha di nuovi impianti da realizzarsi nel periodo 2003-2008. Questo costituisce una frazione sostanziale del contributo complessivo delle foreste al raggiungimento degli obiettivi del Protocollo, pari a 10.2 Mt CO2 eq. anno-1.
Una stima molto più conservativa del potenziale contributo dell’arboricoltura da legno è stata proposta da Anderle et al. ([1]) nella loro analisi dell’assorbimento del carbonio nelle foreste italiane, che indica per il complesso dei rimboschimenti realizzati fra il 1990 ed il 2000 una capacità di fissazione di sole 0.63 Mt CO2 anno-1 per il periodo di riferimento 2008-2012. Va peraltro sottolineato come vengano conteggiati in questa stima solo gli incrementi legnosi cormometrici, che nel caso dell’impianto di Nonantola ammontavano ad esempio al solo 46% della fissazione complessiva nell’ecosistema.
Nel caso dell’impianto di Nonantola, la biomassa legnosa del fusto conteneva come già ricordato all’incirca il 15 % del carbonio presente nell’intero ecosistema (comprensivo della biomassa epigea ed ipogea e della necromassa), e spiegava il 57 % dello scambio netto annuo di carbonio con l’atmosfera, la variabile di reale interesse ai fini del Protocollo di Kyoto. Lo studio condotto dimostra come la quantificazione della capacità degli ecosistemi forestali di sottrarre carbonio all’atmosfera non possa derivare semplicemente dalla misura degli incrementi legnosi, tipico degli inventari forestali, ma debba prendere in considerazione anche l’accumulo di carbonio in radici e suolo, pari nel caso di Nonantola al 41% della capacità complessiva di fissazione del carbonio nell’ecosistema. Valori similari del 35-38 sono stati riportati in rimboschimenti su campi abbandonati in Nord-Europa da Poulton et al. ([19]). Una ripartizione dell’accumulo di carbonio nelle sue diverse componenti (Magnani et al., in preparazione) dimostra come nel caso di Nonantola gran parte dell’accumulo ipogeo sia inoltre da attribuire all’incremento di radici fini e grossolane, mentre l’accumulo di sostanza organica nel suolo ammonterebbe a non più di 2.02 t CO2 ha-1 anno-1. Tale valore non si discosta da quelli generalmente rilevati in terreni agricoli in via di ricolonizzazione da parte della vegetazione naturale ([18]).
Vale peraltro la pena di sottolineare le potenzialità di lungo termine di accumulo di carbonio nei suoli ex-agricoli. In un recente studio su una cronosequenza di afforestazione nella pianura ferrarese, Tonon et al. ([27]) hanno riportato valori di contenuto di carbonio del suolo significativamente superiori in un bosco di 270 anni di età rispetto ad un adiacente impianto di afforestazione di 19 anni. Valori ancora inferiori sono stati rilevati nei terreni agricoli adiacenti (dati non pubblicati). È poi verosimile che tale capacità di accumulo sia particolarmente marcata in suoli come quello di Nonantola a elevato tenore di argilla, per il forte potere stabilizzante di quest’ultima nei confronti del carbonio del suolo ([25]).
L’apporto della componente ipogea fa sì che la capacità di immobilizzazione del carbonio dell’impianto di Nonantola sia più che doppia di quella media riportata da Anderle et al. ([1]), nonostante il fatto che gli incrementi medi massimi stimati nel presente studio non si discostino da quelli assunti nel lavoro citato. Correggendo il dato per questa sottovalutazione, l’analisi di Anderle et al. ([1]) fornisce per i rimboschimenti realizzati nel periodo 1990-2000 un valore di 1.37 Mt CO2 anno-1, ancora superiore a quanto previsto dal Ministero.
Resta da definire quanto i risultati del bosco di Nonantola possano essere considerati rappresentativi di impianti realizzati con specie consimili. I valori di incremento corrente rilevati sono infatti ben inferiori agli incrementi massimi di circa 12 m3 ha-1 anno-1 riportati per frassino maggiore, farnia e rovere in Germania in condizioni di ottima fertilità ([24], [23]), per non parlare degli oltre 42 m3 ha-1 anno-1 riportati per il pioppo ibrido in Pianura Padana ([4]). Al tempo stesso, un confronto con i risultati dell’analisi degli aspetti produttivi dell’arboricoltura, recentemente realizzata dal Gruppo di Selvicoltura dell’Università di Bologna per la Regione Emilia-Romagna, dimostra come gli incrementi osservati non si discostino dalla media di 24 impianti di arboricoltura da legno nella pianura emiliana (dati non presentati).
Vale anche la pena di mettere in relazione lo scambio netto dell’ecosistema misurato a Nonantola con i valori rilevati in un gran numero di boschi europei nell’ambito del progetto EU Carbo-Euroflux ([3]). Il confronto con i valori di assorbimento rilevati su un ampio numero di soprassuoli a scala europea ([28], [29]) dimostra come i tassi misurati a Nonantola, per quanto superiori ai valori rilevati in gran parte d’Europa, siano in effetti inferiori a quelli riscontrabili in diverse formazioni forestali italiane; valori ancora più elevati sono stati invece riscontrati in piantagioni a rapida crescita del Regno Unito, in condizioni ambientali particolarmente favorevoli (Fig. 6). Se invece si limita il confronto ai soli impianti di arboricoltura con specie autoctone su suoli ex-agricoli, i valori medi riportati in letteratura sono compresi fra 2.5 e 3.5 tC ha-1 anno-1 ([19], [31]). Appare dunque realistico ritenere i risultati riportati nel presente studio sufficientemente rappresentativi del contributo dell’arboricoltura nel suo complesso.
Al fine di ottenere una prima stima approssimata del possibile contributo delle Kyoto forests al bilancio nazionale del carbonio, i risultati del presente studio sono stati estrapolati a livello nazionale, moltiplicando la superficie degli impianti realizzati negli singoli anni fra il 1990 ed il 2000 per la capacità di fissazione unitaria di carbonio corrispondente alla loro età al 2010 (vedi Fig. 7). Sulla base di questa stima preliminare, si arriverebbe ad ascrivere agli impianti realizzati nel periodo 1990-2000 una capacità di fissazione pari a 1.51 Mt CO2 anno-1. Come già osservato da Anderle et al. ([1]), peraltro, queste stime non prendono in considerazione il carattere temporaneo di gran parte delle Kyoto forests finora realizzate in Italia, che richiederebbe di interrogarsi anche sul loro destino alla conclusione del turno. Se relativamente al primo periodo di implementazione del Protocollo di Kyoto (2008-2012) il problema riguarda solo gli impianti a pioppo ed eucalitto, non bisogna dimenticare come anche per gli altri impianti produttivi di arboricoltura da legno di qualità il turno prescritto per accedere ai finanziamenti non superi i 20 anni.
La stessa metodologia sopra applicata porterebbe a concludere che i nuovi impianti originariamente programmati per il periodo 2003-2008 a causa della giovane età non giungerebbero a contribuire più di 0.4 Mt CO2 anno-1, pari al 40% della stima proposta ([14]). In questo caso l’obiettivo di arrivare ad assorbire annualmente 1 Mt CO2 eq. attraverso la realizzazione entro il 2008 di nuovi rimboschimenti ([14]) potrebbe essere raggiunto o facendo ricorso a specie più produttive ed in condizioni ambientali non limitanti o aumentando considerevolmente la superficie interessata dagli interventi di afforestazione. Occorre peraltro notare che, sebbene le superfici agricole destinabili all’arboricoltura da legno ammontassero nel 1987 a ben 225000 ha ([15]), tale stima rischi di risultare sovradimensionata, essendo l’arboricoltura proponibile da un punto di vista produttivo ed economico solo in condizioni biologiche e sociali favorevoli ([6]). Resta da definire quanto la crisi diffusa dell’agricoltura italiana, con la marginalizzazione e l’abbandono di terreni di buona fertilità in collina e in pianura, abbia già espanso e possa ulteriormente espandere in futuro la superficie potenzialmente idonea.
In conclusione, i risultati qui presentati confermano l’importanza dell’apporto che l’arboricoltura da legno può dare al miglioramento del bilancio nazionale del carbonio, soprattutto quando si considerino tutte le componenti dell’ecosistema e superando quindi una prospettiva puramente produttivistica. D’altra parte, l’integrazione di dati sperimentali e modelli di simulazione ha sottolineato l’importanza di considerare la dinamica temporale dell’accumulo di carbonio da parte dell’ecosistema, che soprattutto nella fase iniziale di affermazione su terreni ex-agricoli è in larga misura svincolata dall’accumulo di biomassa legnosa. Tali dinamiche dovranno essere prese in considerazione nella pianificazione dei futuri interventi di afforestazione, così da poter prevedere il loro contributo agli obiettivi nazionali di riduzione delle emissioni di carbonio.
Ringraziamenti
La ricerca è stata supportata dai progetto EU-Carboeuroflux, MURST-COFIN 2000 "Bilancio e stock di carbonio negli ecosistemi forestali: determinanti fisiologici, effetti legati all’età, limitazioni ambientali" e COFIN 2003 "Aridità e foreste mediterranee: meccanismi di resistenza e vulnerabilità" e da un finanziamento della Regione Emilia-Romagna, Servizio Parchi e Risorse Forestali e Servizio Aiuti alle Imprese.
References
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar