Observations on natural regeneration in grazed Holm oak stands in the Ogliastra province (Sardinia, Italy)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 8, Pages 103-112 (2011)
doi: https://doi.org/10.3832/efor0665-008
Published: Jul 19, 2011 - Copyright © 2011 SISEF
Research Articles
Abstract
This paper deals with the effects of grazing pressure in two Holm oak (Quercus ilex) stands in Ogliastra (central-eastern Sardinia, Italy), with particular reference to the interactions with the natural regeneration processes. There is a positive interaction (facilitation) between shrubs and seedlings of tree species, as observed in other similar studies carried out by the same authors in other areas of Sardinia. Rubus ulmifolius proved to be the most efficient shrub for the protection and growth of saplings; the other shrub species play a less marked facilitating role. Although the density of shrubs hosting Holm oak seedlings is fairly high, their age and small size confirm that the current grazing pressure is incompatible with any chance of growth of the saplings and success of the natural regeneration. The conservation of these stands must therefore rely on the rationalisation of human activities through the exploitation of a whole range of resources.
Keywords
Facilitation process, Grazing in Forest, Natural regeneration, Holm oaks stands
Premessa
L’attenzione del mondo scientifico e dell’opinione pubblica è da tempo concentrata sui problemi riguardanti la gestione sostenibile delle risorse ambientali, considerato anche che l’azione dell’uomo, negli ultimi cinquanta anni, ha cambiato gli ecosistemi più rapidamente ed in modo più intenso di qualunque altro periodo della storia umana (Valentini in [17]).
La Sardegna non rappresenta un’eccezione e, almeno per quanto riguarda una parte consistente dei territori montani dell’Isola, la mancanza di regolamentazione del pascolo nelle aree boscate, rappresenta una delle maggiori cause di regressione degli ecosistemi forestali ([31], [19]), come del resto descritto anche per altre aree mediterranee.
L’uso del bosco per il pascolo ha in Sardegna e nel resto del mondo radici antiche, e la sua compatibilità con la gestione sostenibile della foresta era resa possibile da alcune consuetudini che alleggerivano la pressione sul bosco da parte degli animali, come la transumanza e il periodico allontanamento del bestiame[1] ([14]). Oggi la gestione dell’attività di pascolo è cambiata, è migliorato il tenore di vita dei pastori, che spesso svolgono anche altre attività, è migliorata la viabilità ed è notevolmente aumentata la diffusione dei mezzi di trasporto per cui la presenza di una guida presso le greggi è quasi sempre saltuaria e quindi anche meno sensibile e attenta alle “necessità” della natura.
Per questi motivi, soprattutto nei boschi di proprietà pubblica gravati da usi civici, al naturale invecchiamento degli alberi si accompagnano una sintomatica carenza di rinnovazione naturale,la compattazione del suolo, l’innesco di fenomeni erosivi, i furti di legname, con fenomeni di degradazione spesso molto intensi, che mettono in serio pericolo la conservazione di questi ecosistemi.
Con il presente lavoro si è inteso integrare una serie di valutazioni condotte su questi argomenti in altre realtà dell’isola ([14], [2], [3]), cercando di caratterizzare l’impatto degli animali domestici al pascolo sulla rinnovazione naturale delle specie quercine, e verificando l’efficacia delle specie arbustive nella protezione dei semenzali.
Il fine ultimo di queste indagini è quello di verificare l’importanza dei fenomeni osservati sulle dinamiche della rinnovazione, in modo da fornire uno strumento utile per definire linee guida di gestione sostenibile e conservazione dei paesaggi tradizionali dell’entroterra sardo.
Materiali e metodi
Le aree di studio
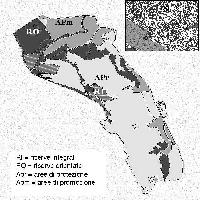
Lo studio è stato condotto in due località della provincia d’Ogliastra: Villagrande Strisaili e Gairo Taquisara.
L’individuazione delle aree da sottoporre a indagine e le successive operazioni di campionamento hanno riguardato terreni ubicati all’interno dei rispettivi Demani Comunali, in zone ben raggiungibili dalla viabilità ordinaria (che facilita il trasporto del bestiame), con copertura boschiva a densità variabile. Sono risultate necessarie due fasi successive d’indagine: (a) caratterizzazione e delimitazione dell’area d’indagine; (b) individuazione delle aree di saggio.
Le aree sono state inizialmente individuate con la collaborazione del personale dell’Ente Foreste della Sardegna e in seguito valutate attraverso l’osservazione delle ortofoto 1:10000 della Provincia Ogliastra. Su tale supporto, sono stati definiti i perimetri delle aree d’indagine facendo riferimento a limiti fisiografici ben riconoscibili sul terreno (crinali, vallecole, corsi d’acqua, strade, ecc.), complessivamente è stata delimitata una superficie di 50 ettari nell’agro di Gairo e di 74 in quello di Villagrande.
Le leccete di Villagrande, situate in località“Donna Concoledda”, sono ubicate su una pendice poco acclive con esposizione sud-est a 900 m s.l.m. La morfologia del terreno varia da tratti quasi pianeggianti con ridotta pietrosità superficiale alternati a modesti rilievi rocciosi. La piovosità media annua (relativa alla stazione di Villagrande) risulta pari a 1012 mm, con massimi autunno-invernali e con minimi estivi (mediamente con periodo di aridità da giugno ad agosto). La temperatura media annua, con riferimento alla vicina stazione di Arzana, è di 15.2 °C.
Per quanto concerne il substrato pedogenetico si tratta prevalentemente di rocce intrusive (graniti, granodioriti, ecc) del Paleozoico e relativi depositi di versante, da cui derivano suoli acidi ascrivibili ai Xerumbrepts, Xerochrepts e Xerorthents, che si caratterizzano per la tessitura in genere franco-sabbiosa con una discreta permeabilità ed erodibilità ([1]).
Secondo la classificazione fitoclimatica del Pavari il territorio ricade nella sottozona media e fredda del Lauretum. Secondo la classificazione di Arrigoni ([4]) la formazione forestale prevalente è caratterizzata dall’orizzonte freddo umido della foresta montana del climax di Quercus ilex e Quercus pubescens; nelle aree più fresche e fredde si vede prevalere Q. pubescens, anche in formazioni pure, che sono state talvolta favorite dall’uomo per la buona integrabilità del pascolo con la modesta copertura della roverella; mentre nei versanti più meridionali ed a quote inferiori domina la foresta pura di Q. ilex. Le specie caratterizzanti la facies erbacea dell’area indagata sono: Centaurea calcitrapa, Euphorbia cupari, Gagea arvensis, Geranium robertianum, Melica uniflora, Pancratium illyricum, Paeonia morisii, Plantago major, Ranunculus bulbosus, Rumex suffocatus, Silene cucubalus, Stachys glutinosa, Stellaria media, Trifolium subterraneum, Viola alba.
Il bosco di leccio campionato a Gairo Taquisara, in località Genna Orruali, è ubicato su un versante esposto a nord-ovest ad una quota di 950 m s.l.m. La pendenza del terreno oscilla fra il 5 e il 15%, con una morfologia più regolare rispetto all’altra area, ma con una pietrosità superficiale molto più diffusa. La piovosità media annua (relativa alla stazione di Gairo) è pari a 958 mm con la stessa distribuzione dell’altra area di studio. Per le temperature medie si fa riferimento anche in questo caso alla sopra citata stazione di Arzana.
Il substrato pedogenetico dominante è rappresentato da metamorfiti (scisti, scisti arenacei, ecc.) del Paleozoico con relativi depositi di versante, i suoli che ne derivano sono essenzialmente Xerorthents, Xerumbrepts e Xerochrepts, a reazione sub-acida, tessitura franco-argillosa mediamente permeabili e di elevata erodibilità ([1]).
La maggior parte del territorio gairese, come nel caso di Villagrande, ricade nella sottozona media e fredda del Lauretum e anche gli aspetti vegetazionali sono paragonabili. Fra le specie caratterizzanti la facies erbacea dell’area di studio ricordiamo: Asphodelus microcarpus, Bellis perennis, Carex flacca, Sedum album, Cetera officinarum, Cyclamen hederifolium, Geranium robertianum, Pancratium illiyicum, Plantago major, Poa bulbosa, Rumex suffocatus, Silene cucubalus, Stachys glutinosa, Stellaria media, Ptilostemon casabonae, Trifolium subterraneum, Viola alba.
Aspetti zootecnici
Non essendo stato possibile rilevare l’effettivo carico di animali presente nelle due aree, nel presente lavoro si fa riferimento a quanto riportato dall’Anagrafe Nazionale Zootecnica (dati aggiornati al 30/09/ 2008) e negli elenchi ufficiali delle ASL di Lanusei.
Nel territorio di Villagrande sono presenti 244 aziende zootecniche di cui il 47 % suinicole, il 32% ovinicole e caprinicole, il 20 % allevano bovini e solo 1% cavalli. Complessivamente sono stati dichiarati 12918 capi di cui il 42% sono caprini, il 41% bovini, il restante suini (9%) e ovini (8%).
Nel territorio di Gairo sono state registrate 127 aziende zootecniche di cui il 52% allevano suini, il 39% ovini e/o caprini il restante bovini. Complessivamente risultano presenti 4625 capi di cui il 77% caprini e ovini, il 21% bovini e solo il 3% suini.
I dati ufficiali rispecchiano da soli un diverso carico di animali, più elevato a Villagrande dove è anche proporzionalmente più elevata la presenza di bovini e suini. Inoltre, come testimoniato dai tecnici locali, in questa zona il bestiame staziona tutto l’anno a differenza dell’area di Gairo dove invece è presente solo nel periodo estivo. I segni del sovraccarico sono comunque evidenti in entrambe le aree: il cotico erboso è povero di specie pabulari ed estremamente discontinuo per la presenza di ampi tratti scoperti caratterizzati da erosione superficiale ed incanalata.
Aspetti sperimentali
Ciascuna area è stata suddivisa, in ambiente GIS, in plot quadrati coincidenti con altrettante maglie del reticolato geografico di 100 metri di lato (rappresentazione Gauss-Boaga), la superficie di ciascun plot è stata ulteriormente suddivisa in 25 subplot quadrati di 20 m di lato (400 m2). Per ciascun plot è stato stimato il grado di copertura arborea attraverso la digitalizzazione delle chiome e sono stati raggruppati in 4 classi di copertura: A < 25 %; 26 < B < 50 %, 51 < C < 75% e D > 76%.
I rilievi al suolo sono stati effettuati in aree di saggio circolari, posizionati al centro dei subplot, a due livelli di dettaglio: (1) 1000 m2 per la valutazione delle caratteristiche componente arborea (Ads1); (2) 400 m2 per la valutazione delle caratteristiche del novellame (Ads4). Per la definizione del campione è stata eseguita la stratificazione dei plot in base alla classe di copertura, procedendo poi all’estrazione casuale dei subplot sulla base della frequenza osservata in ciascuna classe. Complessivamente sono state effettuate: 7 Ads1 e 12 Ads4 a Villagrande, 6 Ads1 e 13 Ads4 a Gairo.
In ciascuna Ads1, per ogni individuo arboreo, sono stati rilevati: specie, diametro a 1.30 m distinguendo fra polloni di ceppaia e individui singoli (matricine), e ne è stata rilevata l’altezza totale (per i polloni è stata rilevata l’altezza di quello dominante).
In ogni Ads4, per ogni arbusto presente, sono state rilevate: specie, l’altezza (hc), la dimensione planimetrica maggiore (L1) e quella ad essa perpendicolare (L2); per ogni semenzale di specie arborea è stata rilevata: la specie, l’altezza (hs), la posizione all’interno dei cespugli (definendola centrale se ricadevano all’interno di una superficie circolare con raggio pari a L2/2, oppure periferiche) e, nel caso in cui fossero presenti gruppi di semenzali, è stato rilevato il numero totale (ns) e l’altezza dell’individuo più alto.
Infine, per avere delle indicazioni circa l’età dei semenzali, sono stati campionati casualmente 30 semenzali collocati all’interno di cespugli per area di studio. Per ciascuno di essi, con l’ausilio di uno stereomicroscopio, è stata eseguita la conta degli anelli sulla sezione trasversale del fusto al livello del colletto.
Elaborazione dei dati
L’area coperta da ciascun cespuglio (S) è stata valutata attraverso la formula dell’ellisse (eqn. 1):
Tutti i dati sono stati elaborati con i comuni test di statistica inferenziale:
- Test del χ2 per saggiare le differenze fra le specie arbustive in termini di presenza o assenza di semenzali di specie arboree al loro interno;
- ANOVA a due vie (schema fattoriale con interazione) per le variabili S, hc, hs (fonti di variazione: specie di cespuglio; presenza o assenza di semenzali);
- test di Duncan in tutti i casi in cui F < 0.05;
- correlazione lineare fra: S, hc e hs.
Risultati
Il soprassuolo arboreo
A Villagrande il soprassuolo campionato è un bosco puro di leccio, probabilmente derivato da un ceduo matricinato, dove tuttora convivono mescolate anche per piede d’albero, piante singole e ceppaie. La densitàè molto irregolare, il grado di copertura medio delle chiome è di 3231 m2 ha-1 (± 245 SE): si alternano tratti a densità colma con zone a fisionomia di pascolo arborato, piante senescenti a chioma rada con individui visivamente più giovani e di grande vigoria.
L’area di Gairo non è strutturalmente dissimile dalla precedente, la copertura media delle chiome è pari a 5836 m2 ha-1 (± 342 SE). Anche qui si alternano piante singole, esclusivamente di leccio e di probabile origine gamica, con ceppaie che danno evidenza delle passate forme di governo. In Tab. 1 sono riportati i parametri dendrometrici medi dei popolamenti.
Tab. 1 - Principali dati dendrometrici medi dei soprassuoli delle due località indagate. (N): numero di individui; (G): area basimetrica; (dg): diametro dell’albero con area basimetrica media; (Hdg): altezza dell’albero con diametro pari a dg.
| Località | Tipo | G | dg | Hdg | |
|---|---|---|---|---|---|
| N ha-1 | m2 ha-1 | cm | m | ||
| Villagrande | Piante singole | 254 | 18.6 | 31 | 11.4 |
| Polloni | 165 | 2.7 | 14 | 9.4 | |
| Totale | 419 | 21.3 | - | - | |
| Gairo | Piante singole | 132 | 13.0 | 35 | 12.9 |
| Polloni | 427 | 7.1 | 15 | 9.9 | |
| Totale | 558 | 20.0 | - | - | |
Il soprassuolo arbustivo
Nell’area di Villagrande sono mediamente presenti 306 ± 33 cespugli ad ettaro, le specie più abbondanti e frequenti sono: l’erica (Erica scoparia, 210 ± 33 cespugli ad ettaro), il rovo (Rubus spp., 138 ± 35 cespugli ad ettaro) e il ginepro comune (Juniperus communis, 63 ± 22 cespugli ad ettaro). Sporadicamente si rilevano anche cespugli di biancospino (Crataegus minogyna, perastro (Pyrus paraster) e prugnolo (Prunus spinosa), complessivamente 23 ± 16 cespugli ad ettaro.
La superficie media dei cespugli è significativamente diversa fra le varie specie campionate (F[3, 143] = 5.17; P < 0.03), la specie che forma i cespugli più grandi è il rovo (3.2 m2) seguita dal ginepro (2.47 m2) ed erica scoparia (1.3 m2). Anche l’altezza media dei cespugli risulta significativamente diversificata fra le varie specie (F[3, 143] = 14.56; P < 0.001): il ginepro raggiunge le altezze maggiori (2.13 m) rispetto a rovo (1.25 m) ed erica (1.22 m) omogenee fra loro. Complessivamente si osserva una correlazione lineare positiva fra superficie ed altezza del cespuglio (R = 0.58; P < 0.001).
A Gairo sono mediamente presenti 421 ± 97 cespugli ad ettaro, le specie più abbondanti e diffuse sono: erica scoparia (129 ± 74 cespugli ad ettaro), ginepro comune (127 ± 39 cespugli ad ettaro) rovo (88 ± 27 cespugli ad ettaro) e rosmarino (Rosmarinus officinalis, 44 ± 25 cespugli ad ettaro). Sporadicamente si rilevano anche cespugli di Rosa canina, biancospino e, più raramente, di timo (Tymus spp.), complessivamente 33 ± 9 cespugli ad ettaro.
La superficie media dei cespugli è significativamente diversa fra le varie specie campionate (F[4, 195] = 3.59; P < 0.01), la specie che forma i cespugli più grandi è il ginepro (3.09 m2) quelli più piccoli sono di erica scoparia e rosmarino (1.7 m2 e 1.8 m2, rispettivamente), mentre quelli di rovo (1.89 m2) e quelli di biancospino e rosa canina assieme (1.9 m2) si collocano in posizione intermedia. Anche l’altezza media dei cespugli risulta significativamente differente fra le varie specie (F[4, 195] = 8.55; P < 0.001), il ginepro raggiunge le altezze maggiori (1.38 m) assieme a biancospino e rosa canina assieme (1.25 m), quelli più bassi sono di erica (0.96 m) e rosmarino (0.76 m). Il rovo presenta altezze intermedie (1.15 m). Anche in quest’area si osserva una correlazione lineare positiva fra superficie ed altezza dei cespugli (R = 0.69; P < 0.001).
La rinnovazione delle specie arboree
Nell’area di Villagrande la maggior parte dei semenzali campionati è di leccio (96 %) i rimanenti di roverella. Mediamente sono presenti 1765 semenzali ad ettaro con una variabilità evidente (CV 54%), ma meno marcata rispetto all’area di Gairo.
Nell’area di Gairo, il numero di semenzali ad ettaro è mediamente pari a 3225 seppur con un’elevata variabilità fra i subplot campionati (CV 108 %), l’unica specie rilevata è il leccio.
Tali differenze dei valori medi non risultano tuttavia statisticamente significative e ciò è da imputare soprattutto all’elevata variabilità esistente nelle aree di indagine.
L’età media dei semenzali è omogenea fra le due aree e pari a circa 10 anni (errore standard 1.7 a Gairo e 1.0 a Villagrande).
L’interazione fra arbusti e semenzali di specie arboree (facilitazione)
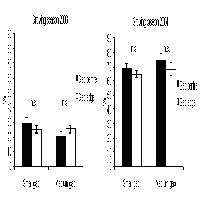
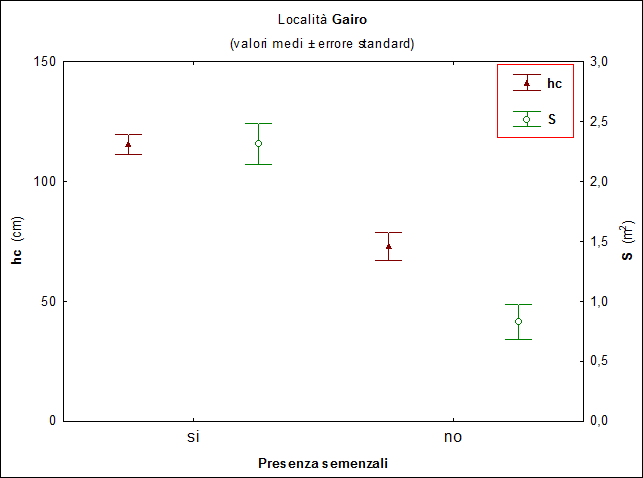
A Gairo, dei 202 cespugli campionati 184 (91%) presentavano al loro interno semenzali, con differenze significative fra le specie costituenti i cespugli (χ2[5] = 12.92; P > 0.05). I cespugli di rosmarino sono quelli che meno frequentemente (71%) presentano semenzali al loro interno mentre quelli di tutte le altre specie, in oltre il 90% dei casi osservati, ospitano semenzali di leccio. La superficie media dei cespugli che presentano semenzali all’interno è significativamente (F[1, 200] = 7.31; P < 0.02) più elevata di quelli senza: 2.31 m2 i primi e 0.83 m2 gli altri. Anche per l’altezza media dei cespugli emergono risultati simili, risultano infatti differenze molto significative (F[1, 200] = 10.26; P < 0.01) con valori pari a 115.6 cm per gli arbusti ospitanti semenzali contro 73.1 cm per gli altri (Fig. 1).
Fig. 1 - Località Gairo: valori medi di altezza (hc, cm) e superficie (S, m2) degli arbusti distinguendo fra quelli con (si) e senza (no) semenzali all’interno. Le barre verticali indicano l’errore standard.
Facendo riferimento ai cespugli con all’interno semenzali, confrontando le diverse specie (ad eccezione della rosa canina per la scarsa rappresentatività del campione), emergono differenze significative rispetto al numero medio di piantine di leccio presenti (F[4, 177] = 3.97; P < 0.01). In particolare, in base al test di Duncan, i cespugli di erica presentano al loro interno un numero medio di semenzali (11) significativamente più alto di quelli di rosmarino (3) e biancospino (5), quelli di rovo (7) e ginepro (9) risultano omogenei fra di loro e si collocano in posizione intermedia. L’altezza media dei semenzali risulta significativamente diversa fra le specie di cespuglio che li ospitano (F[4, 177] = 4.32; P < 0.01). I cespugli di rovo, in base al test di Duncan, presentano i semenzali con l’altezza media più elevata (77.4 cm) assieme a quelli dei cespugli di biancospino (71.0 cm), mentre quelli nei cespugli di rosmarino (38.0 cm) ed erica (45.2 cm) sono quelli più bassi, i semenzali all’interno dei cespugli di ginepro (59.2 cm) si collocano in posizione intermedia. Si osservano differenze molto significative (F[1, 182] =25.01; P < 0.01) in termini di altezza media dei semenzali in funzione della loro collocazione nel cespuglio: quelli collocati nella parte centrale risultano più alti di quelli posti in periferia (74.8 cm e 43.7 cm, rispettivamente).
In generale, come riportato in Tab. 2, si osserva una correlazione lineare positiva fra altezza del cespuglio e altezza del semenzale più alto presente all’interno (R = 0.53; P < 0.01), questa tendenza appare più marcata per i cespugli di biancospino (R = 0.88) e rovo (R = 0.66).
Tab. 2 - Località Gairo Taquisara: correlazione lineare fra hc e hs.
| Specie | Casi | R | P |
|---|---|---|---|
| Rosmarino | 14 | 0.86 | <0.001 |
| Erica scoparia | 60 | 0.52 | <0.001 |
| Ginepro comune | 57 | 0.46 | <0.001 |
| Biancospino | 10 | 0.88 | <0.001 |
| Rovo | 37 | 0.66 | <0.001 |
| Totale | 184 | 0.53 | <0.001 |
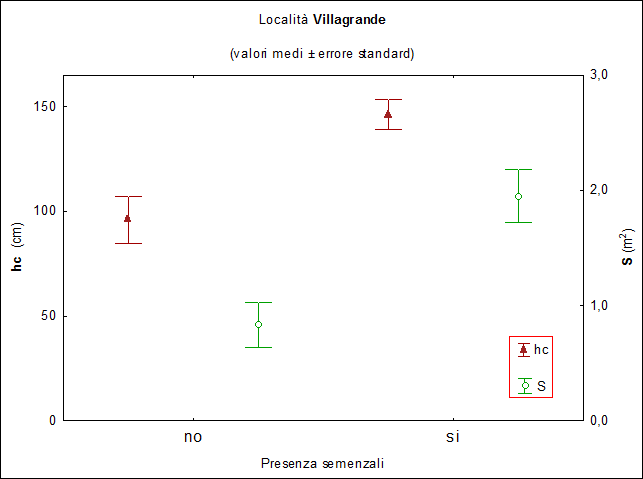
Nell’area di Villagrande, in 127 dei 147 cespugli campionati (86%) sono presenti semenzali di leccio e roverella con differenze molto significative fra le specie arbustive campionate (χ2[3] = 17.82; P > 0.01). I cespugli con la più alta frequenza di semenzali all’interno sono di rovo (95%) seguiti da quelli di erica e ginepro (89 e 87% rispettivamente) mentre per quelli di perastro e biancospino, considerati assieme, la frequenza è significativamente inferiore (45%). Escludendo questi ultimi (per la scarsa rappresentatività del campione), emergono differenze dimensionali significative fra cespugli con e senza semenzali all’interno. Infatti, come riportato nella Fig. 2, la superficie media dei primi con 1.95 m2 risulta significativamente superiore (F[1, 145] = 3.94; P < 0.05) a quella dei secondi 0.83 m2, così anche l’altezza media risulta significativamente (F[1, 145] = 7.3; P < 0.02) più elevata negli arbusti ospitanti semenzali (146.4 cm) rispetto agli altri (96.0 cm). Facendo riferimento ai cespugli che ospitano semenzali emergono differenze significative (F[2, 118] = 6.03; P < 0.01) rispetto al numero medio di semenzali per cespuglio fra le tre specie rappresentative. In particolare il test di Duncan evidenzia che il numero medio semenzali è significativamente più alto nei cespugli di rovo e ginepro (10 e 9 rispettivamente, omogenei fra loro) rispetto a quelli di erica (5). Anche rispetto all’altezza media del semenzale più alto per cespuglio emergono differenze significative fra le tre specie campionate (F[2, 118] = 15.44; P < 0.001). In base al test di Duncan i semenzali più alti si trovano nei cespugli di rovo (153.8 cm) rispetto a quelli di erica e ginepro (43.2 e 43.7 cm, rispettivamente). Inoltre i semenzali collocati nella zona centrale del cespuglio mostrano un’altezza media significativamente (F[1, 145] = 23.79; P < 0.001) superiore a quelli collocati in periferia (133.8 cm i primi e 42.5 cm i secondi)
Fig. 2 - Località Villagrande: valori medi di altezza (hc, cm) e superficie (S, m2) degli arbusti distinguendo fra quelli con (si) e senza (no) semenzali all’interno. Le barre verticali indicano l’errore standard.
Emerge una correlazione lineare positiva (Tab. 3) fra altezza del cespuglio e altezza del semenzale ospitato (R = 0.31; P < 0.01), tuttavia in questo caso tale tendenza appare marcata solo nel caso del rovo (R = 0.83) mentre per le altre due specie prese singolarmente tale correlazione è molto meno evidente e non significativa.
Tab. 3 - Località Villagrande Strisaili: correlazione lineare fra hc e hs. (ns): non significativa.
| Specie | Casi | R | P |
|---|---|---|---|
| Erica scoparia | 74 | 0.18 | n.s. |
| Ginepro comune | 26 | 0.2 | n.s. |
| Rovo | 21 | 0.83 | P<0.001 |
| Totale | 126 | 0.31 | P<0.001 |
Discussione
Caratteristiche dei cespugli
Se si esclude l’erica scoparia la cui diffusione in entrambe le aree è localizzata legata alle zone con terreno più superficiale, ciò che accomuna tutte le specie arbustive presenti (rovo, biancospino, ginepro, rosa canina, prugnolo e perastro) è di essere più o meno provviste di spine, come risultato selettivo dell’azione degli animali al pascolo ([38], [39]). E’ noto infatti che, in ambienti a clima mediterraneo, il carico animale molto elevato incide dapprima sulle specie erbacee con la scomparsa di quelle perenni maggiormente appetite e la loro sostituzione con graminacee annuali (con meccanismi di disseminazione più efficaci in queste situazioni). Successivamente, ma la tempistica dipende anche dalle caratteristiche delle specie (razze) pascolanti, l’azione selettiva incide anche sulla componente arbustiva. Come ultimo stadio di regressione si ha la completa scomparsa della vegetazione e l’insorgenza di fenomeni erosivi più o meno spinti ([30]).
Caratteristiche del “novellame”
La composizione specifica del piano arboreo si riflette in quella dei semenzali censiti: i semenzali di leccio sono gli unici presenti nell’area di Gairo e sono i più frequenti a Villagrande dove possiamo sporadicamente trovare anche semenzali di roverella esclusivamente nelle immediate vicinanze di alcune piante portaseme.
Il microclima luminoso all’interno dei cespugli è un fattore che può risultare selettivo per i semenzali delle specie arboree ([20], [27]). Nel nostro caso la tolleranza all’ombra del leccio può favorirne la sopravvivenza all’interno della chioma degli arbusti. Inoltre non è da sottovalutare l’anatomia della foglia dei semenzali di leccio che, in condizioni di ripetuto pascolamento tende a divenire più coriacea e spinescente, espressione di una maggiore adattabilità al pascolo ([6]). Tuttavia occorre evidenziare che in entrambe le aree di studio il leccio è l’unica specie in grado di disseminare efficacemente.
L’età media dei semenzali campionati, seppur con una certa variabilità, è comunque elevata (10 anni) in entrambe le aree indagate, a conferma di quanto osservato da Alias et al. ([3]) ovvero la tendenza all’invecchiamento dei semenzali all’interno dei cespugli e al contenimento del loro accrescimento longitudinale.
L’azione degli erbivori non incide sul numero dei semenzali che si sono insediati, come del resto dimostrato in altri lavori ([37], [3]), per questo il numero medio dei semenzali non differisce significativamente fra le due aree.
Facilitazione dei cespugli
In entrambe le aree, i cespugli rappresentano gli unici ambienti in cui si possono osservare i processi di rinnovazione naturale della componente arborea: non sono stati rilevati semenzali allo scoperto. In realtà in entrambe le aree si sono osservate ceppaie di leccio con la fisionomia di cespugli, potrebbero essere anch’essi semenzali che, a seguito dell’intensa e ripetuta brucatura, hanno assunto tale portamento. Questo risultato conferma che l’azione di facilitazione è particolarmente importante quando il carico degli erbivori è elevato ([7]). L’interazione fra arbusti e rinnovazione avviene in maniera diversa nei vari stadi del processo di rinnovazione ([36]).
A causa delle caratteristiche morfologiche e della ricchezza di sostanze di riserva, i semi delle specie quercine sono sottoposti con maggiore probabilità all’azione di predatori naturali ([35], [8]). Gli stessi autori, mettono in risalto il ruolo svolto da roditori ed uccelli nella dispersione di questi semi: tra questi il Garrulus glandarius, risulta abbondante nelle zone oggetto di studio. L’allontanamento dei semi dalla pianta madre e la deposizione degli stessi in piccoli serbatoi per opera di questi “agenti di disseminazione secondaria” è tanto più importante quanto minore è il numero di siti sicuri presenti ([33]). È stato ampiamente dimostrato che i cespugli sono siti molto importanti sia nella fase di dispersione ([18], [13], [34]) sia di insediamento, in quanto i semi e le plantule vengono protetti dalla predazione e dal calpestamento degli animali domestici e selvatici (Garfì 1987 in [32], [25], [26]), ciò anche se la sottrazione del seme causata da insetti, roditori ed uccelli può restare elevata proprio all’interno dei cespugli ([28], [40]). All’interno degli arbusti, inoltre, si creano le condizioni microambientali più favorevoli per la germinazione dei semenzali ([23], [24]): minore compattazione del terreno, maggiore umidità, minore insolazione, maggiore disponibilità di nutrienti, minore competizione con le specie erbacee ([10], [16], [22]).
Come anche osservato in altri studi ([5], [3]), la dimensione dei cespugli influisce sulla probabilità di sopravvivenza dei semenzali. Nei cespugli più grandi aumentano le difficoltà per gli animali (soprattutto suini) di predare ghiande e plantule, tale fenomeno è più evidente a Gairo rispetto che a Villagrande. Esistono comunque differenze legate alla specie: le coperture di Rubus ulmifolius e Juniperus communis costituiscono i siti più sicuri per l’insediamento dei semenzali, mentre i cespugli di Erica scoparia lo sono solo nell’area di Gairo. Questo fatto può essere dovuto sia alla scarsa compattezza della chioma di questi arbusti e all’assenza di spine che li rende più facilmente accessibili agli animali in condizioni di maggiore carico.
Nella fase di affermazione, la sopravvivenza e l’accrescimento delle piantine sono soprattutto condizionati dalla brucatura degli animali ([29], [11], [21], [9], [15]). Importanti divengono in questa fase anche i fenomeni di competizione intraspecifica fra cespuglio e semenzale per luce, acqua e nutrienti che, secondo alcuni autori, si traducono in un minore accrescimento delle specie arboree ([9], [34]).
L’altezza media dei semenzali è significativamente più alta nei cespugli di Rubus ulmifolius e Crataegus monogyna rispetto agli altri, anche se in ogni caso le dimensioni sono tali da rimanere entro l’altezza di brucatura degli animali valutata in circa 2 m.
I semenzali hanno manifestato la tendenza ad accrescersi proporzionalmente all’accrescimento longitudinale dei cespugli, in questo senso tale relazione è evidente in quasi tutti i cespugli nell’area di Gairo (anche se più marcata nel caso del rovo e del biancospino) mentre a Villagrande si osserva solo nel caso dei cespugli di rovo. Gli arbusti di Rubus ulmifolius tendono ad accrescersi avvolgendo i semenzali e seguendoli nella crescita e proteggendoli con le spine, stesso comportamento avviene nel caso dei meno frequenti cespugli di rosa canina. Anche i cespugli di biancospino, seppur poco diffusi, dimostrano spesso una buona attitudine nel favorire l’accrescimento dei semenzali.
I semenzali più alti si trovano sempre nella zona centrale del cespuglio, questo fenomeno conferma che l’azione protettiva svolta da quest’ultimi è comunque limitata alle zone più difficilmente accessibili dagli animali.
Siti sicuri per l’insediamento e la sopravvivenza dei semenzali delle specie arboree, si sono occasionalmente dimostrati anche le spaccature delle rocce, fenomeno già descritto nei pascoli arborati nella vicina Orgosolo ([3]). Tuttavia le piantine rimangono confinate all’interno delle fessure della roccia senza possibilità di accrescersi, a causa sia della puntuale brucatura degli animali, che dello scarso spessore del suolo.
Conclusioni
I risultati dei rilievi condotti nelle aree campione di Villagrande e Gairo evidenziano l’efficacia dell’interazione fra cespugli e semenzali delle specie arboree e come questa interazione facilitativa sia specie-specifica ([12]). Il rovo e il biancospino (anche se poco diffuso) sono le specie più efficienti nel favorire l’accrescimento dei semenzali delle specie arboree, probabilmente a causa della loro capacità di formare cespugli alti e poco accessibili. Le altre specie arbustive svolgono invece una azione facilitativa meno efficace, soprattutto nelle condizioni di carico più intenso.
La densità e la distribuzione dei cespugli con all’interno semenzali di leccio risulta relativamente elevata e ben distribuita in entrambe le aree e tale, teoricamente, da assicurare la continuità dei processi di rinnovazione. Di contro l’età media e le ridotte dimensioni dei semenzali, praticamente tutti costretti all’interno dei cespugli, inducono a pensare che la loro possibilità di sfuggire al morso del bestiame, considerati gli attuali livelli di carico, sia praticamente nulla.
Anche se la periodica sospensione del pascolo che avviene nell’area di Gairo offre qualche risultato apprezzabile in termini di densità e accrescimento dei semenzali, la maggior parte dei cespugli ha un’altezza tale da rimanere marcatamente al di sotto della linea di brucatura di bovini e caprini, così come riscontrato nei boschi comunali di Orgosolo ([3]).
Il contesto sociale ed economico che caratterizza i due territori comunali rende improponibile la sospensione, anche temporanea, dell’attività di pascolo, ma la riduzione e la regolamentazione del carico è una misura indispensabile se l’obbiettivo è quello della perpetuazione di questi soprassuoli. Il livello di senescenza è infatti abbastanza elevato e c’è quindi da aspettarsi, oltre a una progressiva diminuzione della copertura delle chiome, anche una costante diminuzione nella produzione di ghiande fertili.
Potrebbero essere individuate senza difficoltà anche altre tecniche di intervento, come l’integrazione della rinnovazione naturale attraverso piantagione di semenzali all’interno di arbusti di dimensioni adeguate, come positivamente sperimentato per la Q. pirenaica in Spagna ([15]) o, in alternativa, recinzioni localizzate in prossimità dei nuclei di rinnovazione esistenti.
La conservazione di questi ambienti è necessaria e possibile ma non può prescindere da una presa di coscienza degli amministratori locali che porti alla razionalizzazione delle attività di utilizzo del bosco e delle altre risorse. In questo senso diviene fondamentale il coinvolgimento partecipativo gli allevatori, magari compensando la minore resa dell’allevamento, con una maggiore rimuneratività dei loro prodotti combinata alla valorizzazione dell’attrattiva turistica di questi ambienti, che facilmente potrebbe diventare strumento di reddito.
Ringraziamenti
Gli autori esprimono un vivo ringraziamento al dott. Stefano Alias per la collaborazione nei rilievi floristici e all’Ente Foreste di Lanusei, in particolare al dott. Salvatore Mele, per il supporto logistico.
References
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar