The conservation of pedunculate oak (Quercus robur) stands in the territory of Parco Lombardo Valle del Ticino (province of Milano and Pavia): main results and management proposals after three years of testing
Forest@ - Journal of Silviculture and Forest Ecology, Volume 10, Pages 1-13 (2013)
doi: https://doi.org/10.3832/efor0825-010
Published: Feb 04, 2013 - Copyright © 2013 SISEF
Research Articles
Abstract
The conservation of pedunculate oak (Quercus robur) stands in the territory of Parco Lombardo Valle del Ticino (province of Milano and Pavia): main results and management proposals after three years of testing.. In the territory of “Parco Lombardo della Valle del Ticino” an experiment, aimed to assess the silviculture pattern suitable for the conservation of stands dominated by peduncolate oak, was set up in 2008. Four experimental areas were located, both in “meso-hygrophilous” and “mesophilous” sites, respectively corresponding to: (a): less intense seed felling; (b): more intense seed felling (both of them aimed to promote the natural regeneration); (c): opening of gap (800 m2) followed by artificial regeneration (both protected and non-protected by shelters); (d): control. Three years after the set up of the experiment, the main results may be summarized as follows: (i): both the seed fellings (the more and the less intense felling) produced a process of natural regeneration, at present insufficient to ensure the continuity of the forest canopy cover; (ii): the branchwood released after utilization produced a positive effect on settlement and growth of peduncolate oak seedlings, attributable to the protection both from the roe deer browsing and wild boar acorn consumption; (iii): ventilated shelters (Tubex Ventex®) produced a positive effect on planting stock, both in terms of survival and growth; (iv): monitoring over time, especially as regards the analysis of the effects of different types of treatment on the processes of regeneration, can provide indications management extensible in similar populations in the territory of the Parco Lombardo Valle del Ticino.
Keywords
Quercus Robur, Regeneration, Secondary Successions, Silviculture, Sustainable Management
Premessa
Nel territorio del Parco del Ticino, il “deperimento” delle piante di farnia ha iniziato a manifestarsi alla fine degli anni ’90. Si tratta di uno stato di sofferenza che, a partire dalla metà degli anni ’80, era stato segnalato per altri boschi quercini del nostro paese ([30]). I sintomi del deperimento si manifestano con trasparenza della chioma, ingiallimenti fogliari, morte delle estremità delle branche (a volte dell’intera chioma), emissione di rami epicormici lungo branche e tronco ([1]) mentre per quanto riguarda le cause del fenomeno, si concorda sul fatto che esse sono attribuibili a un’interazione sinergica di fattori biotici e abiotici ([32]). Per meglio monitorare il fenomeno si sta cercando di mettere a punto una metodologia basata su tecniche di telerilevamento ([31]).
In presenza di questi preoccupanti sintomi, la Regione Lombardia e il Parco del Ticino hanno promosso e finanziato, nell’ambito del programma regionale in campo agricolo 2001-2003, il progetto di ricerca DEPFAR (“Indagini diagnostiche sul deperimento della farnia nei boschi della valle del Ticino”), a cui è seguita, a partire dal 2008, la prosecuzione dal titolo “DEPFAR 2 - Il deperimento della farnia: monitoraggio, indagini eziologiche e proposte di intervento”, nell’ambito della quale si inserisce la presente ricerca (“Il deperimento della farnia: monitoraggio e proposte di intervento selvicolturale”).
Il progressivo deperimento della farnia, al di là degli aspetti fitopatologici, si inserisce nel più ampio problema della conservazione dei boschi a dominanza di questa specie, che rappresentano un elemento caratterizzante del Parco del Ticino, e più in generale, del paesaggio padano. Il problema sta acquisendo rilievo soprattutto alla luce dell’estendersi del deperimento e delle difficoltà di rinnovazione della quercia.
Allo scopo di trovare soluzioni gestionali del problema si è avviata una sperimentazione mirata alla valutazione degli effetti di differenti forme di trattamento selvicolturale.
Il trattamento classico delle fustaie di farnia è rappresentato dai tagli successivi basati su alleggerimenti moderati e progressivi della copertura per favorire l’insediamento e l’affermazione della rinnovazione naturale ([14], [15], [3], [13], [18]).
In assenza di gravi problemi di carattere fitopatologico, i tagli di rinnovazione potrebbero iniziare quando i soprassuoli hanno raggiunto un’età di 150-200 anni, come avviene in popolamenti simili in Slovenia e in Francia ([15], [16]). Sebbene i boschi del Parco Lombardo Valle del Ticino siano tutti molto più giovani, il progressivo deperimento che sta interessando buona parte dei soprassuoli in questione - in aggiunta alla carenza di conoscenze sull’effettiva capacità di risposta dei popolamenti agli interventi selvicolturali - evidenzia la necessità (e l’urgenza) di avviare una sperimentazione finalizzata a individuare forme di trattamento idonee alla conservazione dei querceti.
La presente ricerca, frutto della collaborazione avviata nel 2008 tra un gruppo di ricerca del DEISTAF con il Parco Lombardo Valle del Ticino, si propone di valutare modelli colturali in grado di garantire la conservazione dei popolamenti a dominanza di farnia.
I boschi di farnia del Parco Lombardo Valle del Ticino
La farnia, elemento caratterizzante del bosco planiziario, è ormai confinata in aree relitte, anche a causa dell’attività agricola e industriale che ha eliminato molti boschi di pianura.
La maggior parte dei boschi a dominanza di farnia presenti nel Parco risale al periodo post-bellico, e sono il risultato di una intensa quanto irregolare attività antropica. Sulla base di un’analisi bibliografica (Furlanetto, in [6]), oltre che della consultazione dei piani di gestione presenti nell’archivio del Parco, si è potuto accertare che questi boschi non sono stati oggetto di regolari forme di gestione, in quanto i tagli avvenivano occasionalmente in base alle richieste di legname da opera e di legna da ardere. Fra gli interventi eseguiti in passato vanno segnalati quelli mirati ad ottimizzare la fruibilità dei popolamenti per il pascolo e, soprattutto, per la caccia.
In molti casi, la rinnovazione naturale della farnia era favorita dal controllo della densità dei piani inferiori, mediante la loro periodica utilizzazione (per ricavarne fascina e legna da ardere, e/o per favorirne la percorribilità per i cacciatori) o tramite il passaggio del fuoco ([23]) e spesso integrata, se non addirittura sostituita, da semine e/o piantagioni protette.
Il processo di rinnovazione appare oggi sempre più problematico - oltre che per la giovane età dei soprassuoli e i fenomeni di concorrenza (delle specie del piano arboreo e, soprattutto, di quello arbustivo e erbaceo) e predazione (sia del seme che delle giovani piantine) evidenziati da studi pregressi ([27]) - per il progressivo deperimento delle piante di farnia. Infatti quest’ultimo fenomeno, oltre a diminuire la disponibilità di seme, determina dinamiche evolutive favorevoli a specie arboree molto invadenti come robinia e, soprattutto, ciliegio tardivo. Il problema dell’invasione delle specie esotiche, di difficile soluzione anche in soprassuoli in buone condizioni vegetative, diviene praticamente irrisolvibile in presenza di gravi fenomeni di deperimento a carico della farnia.
In alcuni casi si possono osservare piccoli nuclei (su superfici comprese fra 5 e 100 m2) di piante affermate di farnia, originate da rinnovazione naturale, sempre localizzati al margine dei soprassuoli o delle chiarie che si creano al loro interno, lungo strade o su substrati sabbiosi. La presenza di tali nuclei conferma che, in condizioni di nicchia favorevoli, la farnia non ha problemi a rinnovarsi: resta il fatto che la formazione spontanea di questi gruppi appare insufficiente a mantenere l’attuale fisionomia dei soprassuoli. Il ruolo degli interventi selvicolturali è proprio quello di mirare a creare, laddove le caratteristiche stazionali lo consentano, condizioni analoghe a scala di popolamento.
L’ambiente di studio: Il Parco Lombardo della Valle del Ticino
Il Parco Lombardo della Valle del Ticino ha una superficie di 91410 ettari, di cui circa 1/4 a Parco Naturale e 3/4 a Parco Regionale.
I dati meteorologici della stazione di Abbiategrasso indicano una temperatura media annua di 13.3 °C, una temperatura media del mese più freddo superiore a 0 °C e il gradiente termico annuo di 24 °C. Le precipitazioni annue ammontano a 989 mm (massimo equinoziale con punte massime autunnali, minimo solstiziale con minimo assoluto nel mese di luglio). Secondo l’indice di Gaussen, mediamente non si verificano mesi aridi.
Secondo la classificazione di Pavari ([25]) l’area di studio ricade nella zona fitoclimatica del Castanetum (sottozona calda, I tipo, senza siccità estiva), mentre in accordo con Blasi ([5]), l’area può essere inquadrata nel clima temperato, regione mesoxerica, sottoregione ipomesoxerica, tipo B. La fascia vegetazionale di riferimento corrisponde alla medio-europea di Pignatti ([26]) e alla serie planiziaria della farnia di Ozenda ([24]).
I soprassuoli esaminati vegetano su suoli di tipo Haplic Arenosol ([10]) e sono originati da depositi fluviali.
Nella fase preliminare del lavoro sono stati esaminati diversi popolamenti per definire l’inquadramento tipologico dei popolamenti a dominanza di farnia. Si tratta di soprassuoli che presentano strutture spaziali tendenzialmente semplificate, per lo più biplane, la cui fisionomia riflette la diffusa sospensione delle utilizzazioni e del pascolo, che ha determinato la progressiva affermazione di un piano inferiore di origine agamica.
In sintesi i tipi di riferimento sono:
- Mi: tipo mesoigrofilo, che fa riferimento ai popolamenti delle migliori fertilità.
- Me: tipo mesofilo, il più diffuso, che presenta al suo interno una grande variabilità di fisionomie, in gran parte legate al gradiente di umidità stazionale (in relazione anche alla profondità del suolo).
- Mxe: tipo mesoxerofilo, che caratterizza i popolamenti ubicati al limite inferiore di tolleranza per le disponibilità idriche.
Per maggiori informazioni si rimanda a Bianchi et al. ([4]).
Materiali e metodi
Vista la complessità della problematica, la sperimentazione è stata preceduta da una fase conoscitiva che ha permesso di definire il successivo protocollo sperimentale. Per maggiori dettagli sulla metodologia adottata si rimanda a Bianchi et al. ([4]).
Fase 1: definizione e realizzazione del protocollo sperimentale
Sono state individuate due zone di indagine in popolamenti dominati dalla quercia, localizzate a:
- Motta Visconti (Milano, UTM 32T 498853 E; 5013889 N, quota 77 m s.l.m.), del tipo mesoigrofilo, che fa riferimento ai popolamenti delle migliori fertilità (altezza dominante: 28.20 m)
- Cassolnovo (Pavia, UTM 32T 487075 E; 5025774 N, quota 100 m s.l.m.), del tipo mesofilo, espressivo di fertilità intermedie (altezza dominante: 20.7 m)
I popolamenti individuati sono in buone condizioni sanitarie. Il motivo della scelta è che, pur essendo questa sperimentazione inserita in un progetto sul deperimento della farnia, i meccanismi di rinnovazione naturale della quercia sono poco noti, in particolare in boschi giovani e mai sottoposti a regolare gestione selvicolturale: ne consegue che la valutazione del successo del trattamento in condizioni ottimali è indispensabile prima di valutarne l’applicazione in popolamenti interessati al fenomeno del deperimento.
In ogni zona sono state delimitate 3 aree permanenti di forma quadrata di 2500 m2 ciascuna, separate da corridoi di circa 10 m. In due parcelle sono state sperimentate forme di trattamento mirate a promuovere la rinnovazione naturale della farnia. Una terza area funge da area di controllo per seguire l’evoluzione naturale e per monitorare la produzione di ghianda. In Tab. 1 sono riassunte le caratteristiche dei soprassuoli di Motta Visconti e Cassolnovo.
Tab. 1 - Composizione specifica e principali valori dendrometrici nelle parcelle sperimentali prima del taglio. Il termine “Altre specie” indica rispettivamente: biancospino, ciliegio e nocciolo (zona a), biancospino, ciliegio, eleagno, sambuco, sanguinella, pado e ciliegio tardivo (zona b). (G): area basimetrica; (N): numero di piante.
| Zona | Specie | Area 1 | Area 2 | Area 3 | |||
|---|---|---|---|---|---|---|---|
| N/ha | G/ha, m2 | N/ha | G/ha, m2 | N/ha | G/ha, m2 | ||
| Motta Visconti | Quercus robur | 120 | 9.71 | 132 | 13.93 | 172 | 13.87 |
| Alnus glutinosa | 252 | 11.64 | 224 | 11.76 | 180 | 8.83 | |
| Pinus strobus | 40 | 3.06 | 60 | 3.05 | 56 | 4.33 | |
| Prunus padus | 184 | 0.70 | 264 | 1.05 | 112 | 0.52 | |
| Altre specie | 44 | 0.18 | 88 | 0.38 | 84 | 0.37 | |
| Totale | 640 | 25.29 | 768 | 30.2 | 604 | 27.93 | |
| Cassolnovo | Quercus robur | 252 | 16.16 | 264 | 16.51 | 224 | 12.99 |
| Alnus glutinosa | - | - | 24 | 1.37 | 40 | 3.39 | |
| Populus spp. | 8 | 1.06 | 16 | 2.19 | 28 | 2.36 | |
| Robinia pseudoacacia | 228 | 5.96 | 148 | 3.69 | 32 | 0.61 | |
| Ulmus campestris | 56 | 0.90 | 12 | 0.16 | 52 | 0.61 | |
| Platanus spp. | - | - | - | - | 24 | 3.37 | |
| Altre specie | 160 | 0.62 | 76 | 0.51 | 188 | 1.03 | |
| Totale | 704 | 24.70 | 540 | 24.43 | 588 | 24.35 | |
In tutte le aree è stato eseguito il censimento delle piante, con rilievo del diametro a 1.3 m e dell’altezza di un campione rappresentativo. In ognuna delle tre aree al fine di verificare le condizioni di omogeneità fra le parcelle è stato inoltre eseguito un censimento speditivo dei semenzali di farnia presenti (con misura dell’altezza), in due transect di 20 m x 1m.
Le variazioni del microclima luminoso indotte dal trattamento selvicolturale sono state valutate in termini di irradianza relativa nel campo della PAR (R. I.). Per le misure è stata utilizzata una coppia di ceptometri (Sun Scan Canopy Analysis System, Delta-T Devices Ltd). Le misure sotto copertura sono state eseguite campionando casualmente circa 100 punti al suolo all’interno di ciascuna parcella (Fig. 1). I valori di irradianza in piena luce sono stati rilevati simultaneamente ai rilievi nelle aree, in ampi tratti scoperti adiacenti alle parcelle. Le misure sono state eseguite a 1 m da terra, durante le ore centrali del giorno, in un arco di tempo compreso fra le 11.30 e le 12.30 (ora solare), in giornate serene (situazione corrispondente mediamente a 1850 μmol m-2 sec-1 in piena luce).
Fig. 1 - Misurazione dell’irradianza all’interno di un’area sperimentale.

A Motta Visconti, nel febbraio 2010, è stato progettato ed eseguito un taglio di sementazione (di diversa intensità in ognuna delle due parcelle destinate al confronto delle forme di trattamento), che non può essere tuttavia ricondotto a quello che si esegue per tagli successivi uniformi, dal momento che la distribuzione spaziale delle farnie nel soprassuolo principale era irregolare, così come quella delle altre latifoglie (ontano nero, pado e nocciolo) e della vegetazione arbustiva ed erbacea. Il criterio è stato, per entrambe le tesi (sebbene con intensità di intervento diverse), quello di isolare le chiome delle migliori piante portaseme di farnia. Nel corso dell’intervento sono stati asportati quasi tutti gli individui di pino strobo (in quanto specie esotica) ed il sottobosco. In totale l’area basimetrica è stata ridotta rispettivamente del 48% e del 40% nelle aree F (taglio di sementazione forte) e D (taglio di sementazione debole). La Tab. 2 riporta in dettaglio i dati dell’intervento.
Tab. 2 - Motta Visconti, taglio di sementazione - Numero e area basimetrica ad ettaro delle piante presenti prima dell’intervento, cadute al taglio e presenti dopo il taglio. (F): taglio di sementazione forte; (D): taglio di sementazione debole. (G): area basimetrica; (N): numero di piante.
| Tesi | Specie | PRIMA | TAGLIATE | DOPO | |||
|---|---|---|---|---|---|---|---|
| N/ha | G/ha, m2 | N/ha | G/ha, m2 | N/ha | G/ha, m2 | ||
| F | Quercus robur | 132 | 13.93 | 48 | 3.24 | 84 | 10.69 |
| Alnus glutinosa | 224 | 11.76 | 116 | 6.96 | 108 | 4.80 | |
| Pinus strobus | 60 | 3.05 | 52 | 2.92 | 8 | 0.13 | |
| Altre specie | 352 | 0.38 | 352 | 1.43 | 0 | 0 | |
| Totale | 768 | 30.2 | 568 | 14.55 | 200 | 15.62 | |
| D | Quercus robur | 172 | 13.87 | 32 | 2.48 | 140 | 11.39 |
| Alnus glutinosa | 180 | 8.83 | 76 | 3.65 | 104 | 5.18 | |
| Pinus strobus | 56 | 4.33 | 52 | 4.19 | 4 | 0.14 | |
| Altre specie | 196 | 0.89 | 196 | 0.89 | 0 | 0 | |
| Totale | 604 | 27.93 | 356 | 11.21 | 248 | 16.71 | |
Da ciascuna delle piante tagliate sono state prelevate due rotelle, una alla base per valutare l’età ed una all’altezza di 4 m, per valutare il tempo occorso per arrivare all’altezza superiore a quella del piano arbustivo.
Nella seconda località, a Cassolnovo, si è deciso di rinviare il taglio di sementazione a causa della scarsissima produzione di ghianda.
Il monitoraggio della produzione di ghianda (presupposto della rinnovazione naturale) è stato portato a termine a cadenza mensile nel periodo di caduta delle ghiande, utilizzando 12 trappole permanenti circolari di superficie pari a 0.5 m2 ciascuna, disposte nell’area di controllo (C). Nel Laboratorio Semi del DEISTAF dell’Università di Firenze sono stati successivamente determinati i principali valori quantitativi e qualitativi del seme: numero e peso fresco di ghiande per unità di superficie e capacità germinativa.
In parallelo alle indagini sulla rinnovazione naturale, è stata valutata la possibilità di ricorrere alla rinnovazione artificiale. A tale scopo, in ciascuna zona, alla fine dell’inverno 2008/09 è stato applicato un trattamento a taglio raso a buche su una superficie di 800 m2 (superficie massima prevista dal Piano forestale del Parco) in cui è stato realizzato un impianto con sesto rettangolare (3x2m), alternando file con e senza protezione individuale (shelter Tubex, mod. Ventex®). Sono stati utilizzati semenzali di farnia di 1 anno, allevati con pane di terra, prodotti nel vivaio ERSAF di Curno (Bergamo). Sono stati effettuati due interventi all’anno di decespugliamento e diserbo meccanico.
Fase 2: indagini svolte successivamente agli interventi selvicolturali
Per valutare lo sviluppo della rinnovazione naturale, in ciascuna area sottoposta a taglio di sementazione sono stati realizzati tre transect lineari (materializzati con due picchetti metallici all’inizio e alla fine) di 50 m x 2 m con orientamento Nord-Sud e posizionati uno al centro della parcella e gli altri due rispettivamente a 10 m dai margini Est e Ovest. Nella parcella di controllo sono stati realizzati due transect delle stesse dimensioni, con lo stesso orientamento e posizionati casualmente all’interno della parcella stessa. Ciascun transect è stato suddiviso in sub-plot quadrati di 1 m di lato, per ognuno dei quali sono state osservate e indicate le caratteristiche che avrebbero potuto influenzare la sopravvivenza o lo sviluppo dei semenzali. Sono state così individuate tre categorie:
- presenza di ramaglia, residuo delle utilizzazioni. La ramaglia può rappresentare un ostacolo per l’arrivo del seme sul terreno, rappresenta un ostacolo allo sviluppo dei semenzali ma contiene lo sviluppo della vegetazione concorrente e assicura una maggiore protezione al seme dalla predazione e alle giovani piantine da danni causati da animali o dal passaggio delle persone;
- zona di ristagno dell’acqua nei periodi piovosi, favorita dalla presenza di depressioni del terreno. Il ristagno di acqua determina un ambiente asfittico per lunghi periodi, sia pure in gran parte coincidenti con i periodi di riposo vegetativo delle plantule,
- terreno scoperto (la superficie che non presentava i caratteri già citati).
Nel mese di ottobre di ciascun anno è stato rilevato il numero dei semenzali di farnia e l’altezza di ciascun semenzale; i dati hanno consentito la successiva determinazione per ciascun sub-plot dell’indice di rinnovazione Ir ([20]), comunemente utilizzato per valutare sinteticamente densità e sviluppo della rinnovazione e pari al prodotto del numero di semenzali per m2 (n°) per l’altezza media dei semenzali espressa in cm (h).
I parametri sono stati saggiati con il test ANOVA con successivo test di Duncan (con trasformazione logaritmica per n° e Ir). Per l’anno 2009, precedente all’intervento selvicolturale, visto l’esiguo numero di semenzali osservati, i dati sono stati analizzati con test non parametrici (Kruskal Wallis ANOVA).
Per quanto concerne gli studi sulla rinnovazione artificiale, i rilievi sulle piante messe a dimora sono stati condotti la settimana successiva all’impianto e ripetuti nel mese di ottobre dal 2009 al 2011. Durante i rilievi, per ogni pianta è stata rilevata l’altezza e valutato lo stato di vitalità. Anche in questo caso è stato utilizzato il test ANOVA per testare, per ogni anno e tra le diverse località, l’effetto della presenza/assenza di shelter sull’accrescimento longitudinale.
Risultati
Motta Visconti - Analisi dendrocronologica
L’età delle piante di farnia varia da 32 a 42 anni, il valore medio è di 38 anni (± 0.7); il tempo impiegato mediamente dalle piante esaminate per raggiungere l’altezza di 4 m è di 8.1 anni (± 0.6), variando da un minimo di 6 ad un massimo di 12 anni.
Per quanto riguarda l’ontano nero, l’età varia da 26 a 43 anni (media= 35.3 ± 1.1 anni); il tempo impiegato mediamente per raggiungere l’altezza di 4 m è di 10.9 anni (± 0.6), con un minimo di 6 e un massimo di 16 anni.
Per entrambe le specie non si evidenziano relazioni tra età e/o diametro della pianta e velocità di accrescimento.
Motta Visconti - Microclima luminoso
Nel 2009, prima dell’intervento selvicolturale, non sono state rilevate differenze statisticamente significative tra i valori medi di irradianza relativa (R.I.) nelle tre aree (F2.393=2.99, n.s.). Il valore medio rilevato al suolo è pari a 3.4% (Fig. 2).
Fig. 2 - Variazione, nel tempo, dei valori di irradianza relativa al suolo (R.I.) nelle aree di studio a Motta Visconti. Valori medi e limiti fiduciari (p<0.05).
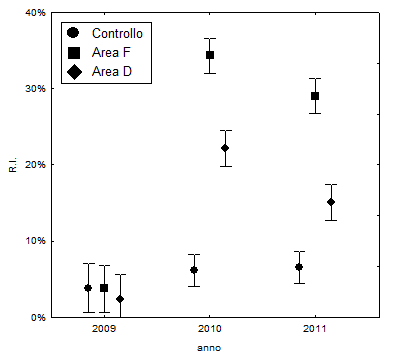
Nell’anno successivo, dopo il taglio, sono emerse differenze molto significative (F2.805=115.91, p<0.001) fra le tesi. In base al test di Duncan risultano significativamente diversi tra loro i valori medi delle tre aree (area F: 34.3%; area D: 22.2%; area C: 6.2%) .
Anche nel 2011 si conferma la tendenza emersa nel 2010, benché su valori leggermente decrescenti (F2.781=93.16, p<0.001). In base al test di Duncan i valori medi delle tre aree sono ancora significativamente diversi tra loro (area F = 29.0%; area D = 15.1%; area C = 6.6%). La diminuzione osservata nel 2011 è probabilmente dovuta alla rapida espansione delle chiome delle piante in seguito al loro isolamento, a sua volta riconducibile alla giovane età delle piante.
Nell’area di controllo i valori medi di R.I. evidenziano un leggero aumento nel 2010 (rimanendo di fatto costanti nel 2011): ciò è dovuto, verosimilmente, alla caduta di due piante a seguito di eventi meteorici.
Monitoraggio fruttificazione della farnia
A Motta Visconti, nel 2009 sono state raccolte, mediamente, 207.7 ghiande per m2 (con una facoltà germinativa del 46.9%), per un peso di 462.6 g/m2. Nei due anni successivi il numero medio di ghiande per m2 è sceso rispettivamente a 16.8 e 13.2; inoltre si è ridotta drasticamente la facoltà germinativa delle ghiande (rispettivamente 4% e 17%).
Diverso è l’andamento registrato a Cassolnovo, dove, mediamente, nel 2009 sono state rilevate al suolo 24.2 ghiande per m2, con una facoltà germinativa del 3%; il numero aumenta nel 2010 (69.5 ghiande per m2, con una facoltà germinativa che sale al 39.1%) pur rimanendo nettamente inferiore a quello rilevato l’anno precedente a Motta Visconti, per poi crollare nuovamente nel 2011 con 15.3 ghiande per m2 (mentre la facoltà germinativa scende al 3.3% - Tab. 3).
Tab. 3 - Principali dati relativi alle ghiande prodotte nelle due località di studio.
| Località | Anno | Numero/m² germinabili | Numero/m² non germ. |
|---|---|---|---|
| Motta Visconti | 2009 | 97.5 (± 95.07) | 110.17 (± 77.10) |
| 2010 | 0.67 (± 1.25) | 16.17 (± 15.91) | |
| 2011 | 2.25 (± 3.39) | 11.00 (± 8.94) | |
| Cassolnovo | 2009 | 0.83 (± 1.72) | 23.33 (± 22.14) |
| 2010 | 27.17 (± 21.22) | 42.33 (± 18.88) | |
| 2011 | 0.50 (± 1.12) | 14.83 (± 7.74) |
Motta Visconti - Analisi della popolazione di semenzali di farnia
Il numero di semenzali di farnia rilevati prima del taglio era molto basso (Tab. 4). È interessante notare come tale valore sia rimasto pressoché immutato nell’area di controllo (C) dal 2009 al 2011, mentre è aumentato nelle due aree sottoposte a intervento selvicolturale, pur rimanendo su valori di densità non elevati.
Tab. 4 - Numero di semenzali di farnia per m2 presenti nelle tre aree di Motta Visconti: confronto fra le aree prima del taglio e nei due anni successivi, (C): controllo; (F): taglio di sementazione forte; (D): taglio di sementazione debole.
| Area | 2009 | 2010 | 2011 | ||
|---|---|---|---|---|---|
| C | 0.45 | 0.45 (±0.76) a | F2.787=90.803 p<0.001 |
0.41 (±0.91) a | F2.795=36.337 p<0.001 |
| F | 0.10 | 1.27 (±1.95) b | 1.09 (±1.84) b | ||
| D | 0.12 | 2.97 (±3.66) c | 1.67 (±2.27) c | ||
I valori maggiori, già nel 2010, si sono osservati nell’area D. Nel corso del 2011 è stata evidenziata una diminuzione del numero di semenzali, probabilmente per effetto della mortalità non compensata da nuove nascite, che è stata direttamente proporzionale alla densità osservata nel 2010.
La distribuzione dei semenzali sul terreno è molto localizzata (Fig. 3): nel 2011 quasi il 60% della superficie campionata nell’area F e il 40% nell’area D erano prive di semenzali, mentre la percentuale sale a quasi l’80% nell’area di controllo.
Fig. 3 - Semenzali di farnia rilevati nelle aree sperimentali di Motta Visconti: frequenza in relazione alla densità.
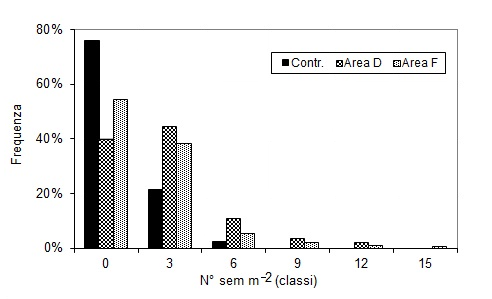
L’altezza media dei semenzali è superiore prima del taglio in tutte le aree; successivamente all’intervento il valore è diminuito nel 2010 per tornare ad aumentare, sia pure di poco, nel 2011 (Tab. 5)
Tab. 5 - Altezza dei semenzali di farnia nelle aree sperimentali di Motta Visconti: confronto fra anni e tra trattamenti (C): controllo; (F): taglio di sementazione forte; (D): taglio di sementazione debole.
| Area | 2009 | 2010 | 2011 | |||
|---|---|---|---|---|---|---|
| C | 37.5 (± 25.9) a | Kruskal-Wallis H: (2, N= 27) = 6.93 p <0.05 |
12.4 (± 7.5) a | ANOVA F2.1482=10.10 p<0.001 |
14.7(± 8.43) a | ANOVA F2.911=33.94 p<0.001 |
| F | 52.5 (± 34.0) ab | 12.9 (± 5.8) ab | 19.4(± 9.70) b | |||
| D | 21.0 (± 5.5) ac | 11.4 (± 5.1) ac | 14.7(± 6.92) a | |||
Nella valutazione di questo dato occorre considerare che, prima del taglio, tra i pochi semenzali presenti nelle aree C e F, alcuni avevano un’altezza di circa 1 m, contribuendo così a innalzare il valore medio. Questi semenzali non sono stati ritrovati nella seconda fase. Nelle aree sottoposte ad intervento è ipotizzabile la loro asportazione o il forte danneggiamento durante le diverse fasi dell’utilizzazione, mentre nell’area C, dal momento che i transect avevano una posizione non coincidente con quelli delimitati prima del taglio, è possibile che non siano stati compresi nei nuovi rilievi.
Nel 2011, l’altezza dei semenzali varia da un minimo di 4 cm a un massimo di 69 cm, tuttavia il 95.4% dei semenzali presenta un’altezza inferiore a 31 cm. L’incremento maggiore è stato registrato nell’area F.
La posizione all’interno della tagliata, che determina differenze in termini di disponibilità luminosa, sembra non influire in modo particolare sull’accrescimento dei semenzali: in entrambe le aree il margine Ovest (che risente in misura maggiore dell’ombreggiamento della parte non tagliata) presenta altezze medie minori rispetto alla parte centrale e alla parte più illuminata, per quanto solo nell’area D le differenze siano statisticamente significative.
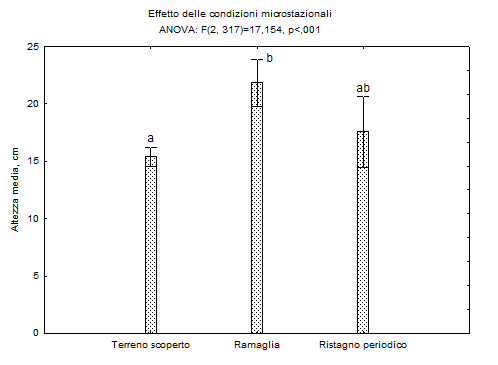
Le condizioni microstazionali sembrano invece influire maggiormente sull’altezza dei semenzali (Fig. 4), con effetti positivi dovuti alla presenza di ramaglia ma anche al ristagno di acqua, sebbene in questo caso si osservi una più marcata variabilità.
Fig. 4 - Effetto delle condizioni microstazionali (ramaglia, ristagni di acqua, terreno scoperto) sull’altezza dei semenzali nelle aree di studio di Motta Visconti: le lettere indicano i gruppi omogenei.
Per quanto riguarda l’effetto dei tagli di sementazione sull’avvio del processo di rinnovazione (espresso da Ir), l’insediamento e la prima affermazione dei semenzali di farnia risultano nettamente più marcati nelle aree in cui si è intervenuto rispetto al controllo (Fig. 5). Solo nelle tesi D e F si osservano sub-plot con valori di Ir superiori a 40.
Fig. 5 - Valori di Ir nelle tre aree di studio di Motta Visconti, riferiti al 2011. Valori medi e limiti fiduciari (p<0.05): le lettere indicano i gruppi omogenei.
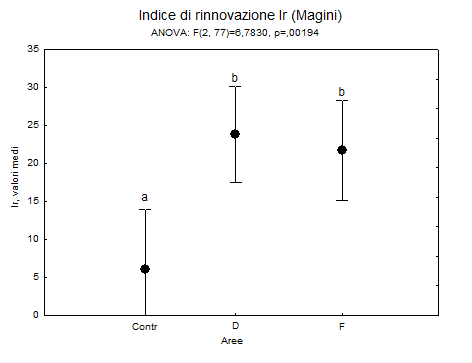
Il test ANOVA relativo al confronto tra i valori medi del numero di semenzali rilevati nei vari sub plot, evidenzia differenze statisticamente molto significative fra le tesi (Fgl 2;77=6.78; P<0.02). In base al test di Duncan i valori medi sono significativamente più bassi nel controllo (Ir = 6) rispetto alle tesi D ed F (24 e 23 rispettivamente), che risultano omogenee fra loro.
Rinnovazione artificiale
Sono emerse differenze notevoli tra le due località in termini di sopravvivenza delle piante messe a dimora. Mentre a Motta Visconti la protezione delle piante ha fatto registrare una superiorità abbastanza contenuta rispetto al testimone, a Cassolnovo (ambiente mesofilo tendente al mesoxerofilo, ma soprattutto caratterizzato da una ben superiore presenza di caprioli), la protezione è stata decisiva nel limitare la mortalità delle piante. Nel 2011, a Cassolnovo il valore di sopravvivenza registrato per le piante protette è quasi 9 volte superiore rispetto al testimone, mentre a Motta Visconti solo 1.3 volte (Tab. 6). In entrambe le località (Cassolnovo: χ21=114.5, p<0.01; Motta Visconti: χ21=12.54, p<0.01) si registrano comunque differenze significative mettendo a confronto la presenza e l’assenza di protezione.
Tab. 6 - Andamento della sopravvivenza delle piante messe a dimora, durante i 3 anni di osservazione, nelle due località in cui si è condotta la sperimentazione: confronto tra piante protette con shelter e piante non protette.
| Località | Protezione | 2009 | 2010 | 2011 |
|---|---|---|---|---|
| Cassolnovo | Shelter | 93.2% | 85.5% | 69.2% |
| Nessuna | 36.2% | 12.9% | 8.0% | |
| Motta Visconti | Shelter | 94.1% | 86.6% | 81.5% |
| Nessuna | 89.5% | 71.8% | 62.4% |
Da sottolineare che, mettendo a confronto solo le piante protette, nei primi due anni, in base all’analisi del χ2 non risultano differenze significative di sopravvivenza tra le due località. Solo nel 2011 si osserva una mortalità significativamente superiore a Cassolnovo (31%) rispetto a Motta Visconti (18.5%). Più in particolare, nell’anno in questione l’analisi del χ2 ha evidenziato differenze significative tra le due località sia per le piante senza protezione (χ21=109.6, p<0.01) che per quelle protette da shelter (χ21=4.82, p<0.05).
Nell’area di Motta Visconti, gli incrementi longitudinali medi annui sono risultati crescenti dal 2009 al 2011, sia per le piante protette da shelter che per quelle senza protezione. In particolare, l’analisi della varianza ha evidenziato differenze statisticamente significative di accrescimento a favore delle piante protette rispetto a quelle non protette nel 2009 (F1.208=55.08; p<0.001) e nel 2010 (F1.208=67.38; p<0.001), mentre le differenze risultano non significative nel 2011 (F1.208=3.66; n.s.).
Considerando le altezze raggiunte dalle piante a tre anni dall’impianto, si può notare che soprattutto le piantine protette hanno raggiunto valori elevati. A Motta Visconti, ove non si sono registrate brucature da parte dei caprioli, anche le piante non protette hanno fornito in questo senso buone performance (Tab. 7), per quanto inferiori a quelle delle piante cresciute all’interno degli shelter (ANOVA: Motta Visconti, F1.208=79.00; p<0.001; Cassolnovo F1.92 =58.40; p<0.001).
Tab. 7 - Altezza media e massima (in cm) dei semenzali di farnia rilevati nel 2011 nelle due località di studio, distinguendo la presenza o meno di shelter nelle prove di rimboschimento.
| Shelter | Motta Visconti | Cassolnovo | ||
|---|---|---|---|---|
| H media | H max | H media | H max | |
| Con shelter | 137.9 (± 43.2) | 224 | 113.2 (± 44.1) | 206 |
| Senza shelter | 86.3 (± 40.7) | 218 | 17.9 (± 20.0) | 80 |
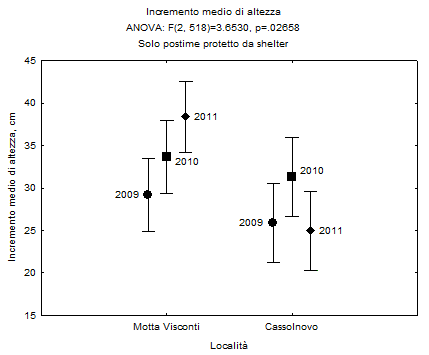
Va infine messo in evidenza (Fig. 6) che l’accrescimento longitudinale delle piante cresciute negli shelter è simile nelle due località nel 2009 (F1.175=0.86; n.s.) e nel 2010 (F1.175=0.24; n.s.), mentre nel 2011 risulta significativamente superiore a Motta Visconti (F1.175=15.42; p<0.001).
Fig. 6 - Incremento di altezza delle piante protette con shelter, separatamente per località e anni di osservazione. Valori medi e limiti fiduciari (p<0.05).
Discussione
I boschi oggetto di indagine sono significativi di alcune fisionomie tipiche dei boschi del Parco. In considerazione del deperimento in atto, per garantire il mantenimento dei popolamenti a dominanza di farnia del Parco Lombardo Valle del Ticino, è necessario quindi avviare un processo di rinnovazione che porti a sostituire le piante morte o fortemente deperienti.
La nostra indagine ha evidenziato alcuni elementi critici nel processo di rinnovazione.
Il primo è rappresentato dalla disponibilità di seme con elevata capacità germinativa. Se è vero che, in generale, la farnia presenta una spiccata alternanza nel tempo per quanto riguarda la produzione di seme, a ciò va aggiunto che, nel caso specifico, si tratta di popolamenti giovani che non hanno raggiunto ancora un livello di fruttificazione sufficientemente abbondante. Il problema è aggravato dalle condizioni di deperimento, che potrebbero richiedere interventi selvicolturali urgenti a prescindere dalla disponibilità di seme. Dato il lungo periodo di attesa necessario per ottenere una sufficiente rinnovazione da seme, è stata valutata la possibilità di integrare la rinnovazione naturale con semine o piantagioni.
Un altro ostacolo è rappresentato dai fenomeni di concorrenza che la vegetazione spontanea, costituita da specie del piano arboreo, arbustivo ed erbaceo, esercita sui semenzali. Per questo, una possibile soluzione è quella di associare ai tagli successivi ripuliture del sottobosco ([8]). In realtà, nelle aree di studio, a distanza di due anni dagli interventi, le vegetazione spontanea (dominata, oltre che da ricacci di Corylus avellana, Prunus padus e Alnus glutinosa, da Rhamnus frangula, Rubus spp. e Osmunda regalis) non ha raggiunto né una densità né uno sviluppo in altezza tali da compromettere la sopravvivenza dei semenzali di farnia.
In questo momento, tuttavia, il principale fattore di disturbo per le dinamiche di insediamento e affermazione del novellame sembra essere rappresentato dalla pressione degli ungulati selvatici (capriolo e cinghiale) che appare crescente negli ultimi anni in tutto il territorio del Parco: la predazione interessa sia il seme che le giovani piantine ([27]).
In realtà, in assenza di fenomeni di predazione da parte della fauna selvatica e di concorrenza della vegetazione spontanea, la farnia è capace di rinnovarsi con successo, fino a dimostrare sorprendente capacità di colonizzazione di spazi aperti, come confermano indagini recenti condotte in campi abbandonati del Piemonte ([22]). Una ulteriore conferma in questo senso proviene dalle nostre indagini, ed è fornita dai i dati di sopravvivenza e accrescimento delle piantine non protette da shelter a Motta Visconti.
Vale la pena, a questo proposito, di fare una breve rassegna sull’ecologia della rinnovazione della specie.
Secondo Calvo et al. ([7]) il seme di farnia germina con una certa facilità, e i semenzali sopravvivono bene, almeno inizialmente. Come evidenziato anche nel Bosco delle querce di Seveso e Meda, nelle prime fasi di insediamento la densità dei semenzali può effettivamente essere molto alta ([21]) soprattutto a seguito di annate di pasciona, ma dopo pochi anni inizia la mortalità, imputabile alla scarsa luminosità a livello del suolo, che favorisce altre specie più adattate all’ombra a scapito della farnia. In particolare, studi condotti da Welander & Ottosson ([33]) indicano che nel primo anno i semenzali si adattano all’ombra, per cui è possibile il loro insediamento anche sotto densa copertura; è negli anni successivi che la disponibilità luminosa diventerebbe decisiva. La mancanza delle operazioni colturali, svolte in passato per il contenimento della vegetazione concorrente, condiziona perciò negativamente la rinnovazione della quercia.
Un fattore determinante è dunque la disponibilità luminosa ([19], [9], [7]). Secondo studi condotti da Von Lupke ([18]), all’interno di querceti misti, per assicurare lo sviluppo giovanile dei semenzali di farnia occorre un’irradianza relativa (R.I.) superiore al 15-20%, altrimenti a livelli inferiori le piantine muoiono entro 2-6 anni. Sulla base dei nostri risultati, in entrambe le aree in cui si è intervenuto si sono oltrepassati tali valori minimi. Tuttavia nei due anni seguenti l’intervento si è osservata una progressiva riduzione dei valori di irradianza al suolo, fattore probabilmente legato alla grande reattività delle chiome delle piante (dovuto sia alla giovane età sia alla fertilità della stazione).
Possiamo affermare che a Motta Visconti il taglio di sementazione è stato efficace nel creare le condizioni favorevoli all’inizio di un processo di rinnovazione, anche se non ancora sufficiente a garantirne la piena affermazione. Queste prime osservazioni suggerirebbero, perciò, la necessità di procedere in tempi brevi a eseguire un taglio secondario, magari in concomitanza con un’annata di buona produzione di ghianda.
La luce non può essere considerato il solo fattore chiave per la rinnovazione della farnia; è stato dimostrato che, per la necessità di massimizzare l’accrescimento in altezza, il novellame della quercia è molto condizionato anche dallo stress idrico ([28]), sebbene, piuttosto che l’azione separata dei singoli fattori ecologici, sembra sia l’interazione della luce con il contenuto idrico del suolo a influenzare sia la sopravvivenza che l’accrescimento giovanile dei semenzali di farnia ([17], [29]).
Studi condotti nel Bosco Fontana (Mantova) hanno dimostrato che l’accrescimento dei semenzali è controllato principalmente dalla luce, mentre la disponibilità idrica esercita influenza significativa solo in condizioni di disponibilità radiative non limitanti ([13]).
Per contro, al crescere dell’afflusso radiativo, aumenta l’aggressività di specie del sottobosco, come il rovo, che rappresentano in questi casi i principali agenti di mortalità a causa della concorrenza per le risorse, indipendentemente dall’umidità del suolo.
Fra i risultati di maggiore interesse emersi da questi primi anni di sperimentazione, vale la pena di segnalare l’effetto positivo esercitato dalla ramaglia sulla rinnovazione della farnia. L’accumulo di ramaglia favorisce i semenzali verosimilmente a causa di un’azione protettiva, che si esprime in termini sia di un microclima più umido (favorevole a sviluppo e sopravvivenza di plantule esigenti di umidità atmosferica come quelle della farnia), sia di difesa dal morso degli animali, come è stato dimostrato, per altre specie arboree, in boschi cedui della Maremma ([12]). Nel nostro caso è ipotizzabile che la presenza della ramaglia abbia ridotto la predazione delle ghiande da parte dei cinghiali o di altri animali.
Come è stato anticipato, il ricorso alla rinnovazione artificiale, anche solo come integrazione alla rinnovazione naturale, appare necessario per garantire la dominanza della farnia. Nel nostro caso, per proteggere il postime dagli ungulati sono stati utilizzati shelter areati che, oltre alla protezione, garantiscono un effetto positivo in termini di performance, attribuibile alla creazione di un microclima caratterizzato da maggiore umidità atmosferica e al cosiddetto “effetto camino”, cioè al flusso continuo di aria attraverso i fori di ventilazione della parte inferiore del tubo, capace di stimolare la fotosintesi e la traspirazione, come è stato dimostrato anche nel caso di postime di Prunus avium ([11]). A Motta Visconti, situazione espressiva delle condizioni ottimali di vegetazione della farnia (ambiente mesoigrofilo, ridotta presenza di cervidi), l’effetto positivo in termini di performance degli shelter è evidente soprattutto nei primi anni: già a partire dal terzo anno si osserva infatti un netto recupero delle piante non protette. Rimane tuttavia importante la possibilità per le piantine di portare velocemente la porzione apicale oltre l’altezza di accessibilità degli ungulati. A Motta Visconti, a tre anni dall’impianto, l’80% circa delle piante protette si trova in queste condizioni, contro il 32% di quelle non protette, che pure non hanno subito danni dovuti a brucature. E’ pur vero, però, che in presenza di una consistente popolazione di caprioli, si possono osservare brucature sulla parte apicale delle piantine che fuoriescono dagli shelter, fenomeno osservato nell’impianto di Cassolnovo, dove alla brucatura sono in parte imputabili le differenze di performance registrate il terzo anno rispetto all’impianto di Motta Visconti.
L’efficacia degli shelter ventilati sulle performance del postime si manifesta in misura ancora maggiore in ambienti relativamente più xerici ([2]), come Cassolnovo, dove gli incrementi di altezza, almeno nei primi due anni, risultano in linea con quelli osservati a Motta Visconti.
Conclusioni
La situazione attuale dei boschi a dominanza di farnia del Parco Lombardo Valle del Ticino è problematica al punto da presentare aspetti di vera e propria emergenza a causa del deperimento che, da alcuni anni, minaccia la sopravvivenza di interi popolamenti. Questo fenomeno è aggravato da altri fattori, fra cui assumono particolare importanza l’invadenza delle specie esotiche e della vegetazione spontanea, che in ambienti mesofili e mesoigrofili come quelli studiati assume notevole vigore, e la crescente pressione degli ungulati selvatici, che limita i processi di rinnovazione della quercia.
Il bosco a dominanza di farnia rappresenta una tessera importante del paesaggio della pianura lombarda: il punto chiave della questione è rappresentato dal futuro dei querco-carpineti, con una partecipazione più o meno consistente di farnia, la cui conservazione richiede la progressiva sostituzione delle piante deperienti (in certi casi di interi popolamenti) attraverso processi di rinnovazione che però si verificano, normalmente, in boschi di età ben più avanzata.
In questo senso la sperimentazione descritta nel presente lavoro presenta interessanti prospettive in termini di risultati applicabili sul piano gestionale. Dal monitoraggio futuro e dall’analisi degli effetti del trattamento sulla rinnovazione potranno scaturire indicazioni gestionali estendibili a popolamenti simili nel territorio del Parco. Al momento, a soli tre anni dall’inizio della sperimentazione, la valutazione dei processi di rinnovazione non può essere considerata esaustiva. Tuttavia, in base ai primi risultati, appare chiaro che la rinnovazione naturale dovrà essere integrata con quella artificiale. In questo modo dovrebbe essere garantito il futuro dei boschi a partecipazione di farnia, anche se non necessariamente dominati da tale specie.
Ringraziamenti
Il lavoro è stato svolto con contributo del Parco lombardo della Valle del Ticino. Gli autori ringraziano Roberto Castrovinci per la collaborazione ai rilievi in campo.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar