Choroplastic diversity in Italian oaks: evidence of a higher genetic richness in southern and insular populations
Forest@ - Journal of Silviculture and Forest Ecology, Volume 1, Pages 82-87 (2004)
doi: https://doi.org/10.3832/efor0220-0010082
Published: Dec 28, 2004 - Copyright © 2004 SISEF
Research Articles
Abstract
Results of the analysis of non-coding regions of the chloroplast genome in Italian population of deciduous oaks (subgenus Quercus) and holm oak (subgenus Scleropyllodris) are reported. Deciduous oak revealed the occurrence of six chloroplast haplotypes, differently distributed in the single geographic areas (northern, central, and southern Italy, and major islands). Moreover, most of the present Italian oak populations seems to derive from two major refugia, located in Sicily and in the Balkans, respectively. The most likely migration dynamics from the refugia towards the central and northern part of the peninsula are discussed. Migration pathways resulted to have been different in the various geographic areas. The presence of an endemic haplotype in Sardinia and Corsica was considered of particular interest. In Italian holm oak populations eight haplotypes were detected, six of them in Sicily. Five out of six Sicilian haplotypes resulted to be endemic. The Italian peninsula was colonized by two major haplotypes, one of them originating from Sicily. Results on deciduous oaks and holm oak in Italy highlight the important role played by the populations from the south and from the major islands in the colonization the Italian peninsula after the last ice age.
Keywords
Quercus, Chloroplastic diversity, Phylogeography, Glacial refugia, Colonization, Conservation of genetic resources
Introduzione
La storia della vegetazione nel nostro continente è stata determinata dalle forti modificazioni ambientali che hanno caratterizzato tutto il Quaternario (gli ultimi due milioni di anni). Durante questo periodo l’alternanza tra fasi fredde o glaciali e fasi temperate ha modificato continuamente la copertura vegetale: questa si è ridotta alle sole aree rifugio durante le fasi glaciali e ha dato vita a nuove migrazioni al sopraggiungere di ogni nuova fase temperata. Questo processo si è verificato anche al termine dell’ultimo periodo glaciale, conclusosi in Europa circa 13.000 anni fa (datazione 14C). L’attuale composizione della vegetazione è il risultato di questo processo.
I reperti fossili, in particolare quelli pollinici, testimoniano la presenza dei rifugi glaciali. L’identificazione tassonomica di granuli pollinici e la relativa datazione al radiocarbonio consentono di determinare la presenza di famiglie e di generi vegetali (o, più raramente, di specie) in un dato sito in epoche successive (spettro pollinico). Le testimonianze fossili raccolte finora indicano le tre penisole del Mediterraneo (la penisola iberica, quella italiana e quella balcanica) come i principali rifugi per molti generi, incluso il genere Quercus ([17], [3], [31]).
La dinamica di migrazione dalle aree rifugio verso le zone più settentrionali del continente segue modelli che sono comuni agli organismi vegetali e a quelli animali (descritti da [2], [15], [16], [30]). Tuttavia, le modalità e la velocità di migrazione variano da specie a specie e dipendono fortemente dalle caratteristiche biologiche delle singole specie, in particolare dai meccanismi di dispersione del seme, che nelle querce è affidata principalmente alla gravità (disseminazione propinqua), e in parte a uccelli e piccoli mammiferi responsabili delle disseminazione a grandi distanze (longinqua). E’ stato stimato che dopo l’ultimo periodo glaciale l’espansione delle piante forestali è stata molto rapida, in particolare per le querce il tasso di migrazione è stato di 500 metri all’anno ([19]).
L’analisi del polline fossile può essere combinata con quella del DNA di origine materna, che, proprio perché trasmesso attraverso le generazioni via seme, è particolarmente indicato per studi di filogeografia (lo studio delle relazioni che intercorrono tra la genealogia dei geni - filogenesi - e la distribuzione geografica delle popolazioni - [1]) e per la definizione delle più probabili vie di migrazioni seguite dagli organismi vegetali.
La diversità cloroplastica è stata studiata a diverse scale geografiche in molte Fagaceae ([5], [7], [8], [9], [10], [11], [18], [20], [22], [24], [25]) e nel genere Quercus l’ereditarietà materna dei cloroplasti è stata dimostrata da Dumolin et al. ([6]). Gli studi di filogeografia realizzati sulle querce europee hanno consentito di definire le dinamiche di colonizzazione e le più verosimili rotte migratorie avvenute alla conclusione dell’ultimo periodo glaciale per le querce decidue (subgenus Quercus) e per il leccio (subgenus Scleropyllodris).
In questo studio vengono riassunti i risultati ottenuti dall’analisi di popolazioni italiane mediante marcatori molecolari, con il principale obiettivo di mettere in evidenza l’importante ruolo avuto dai rifugi localizzati nella penisola italiana per la colonizzazione del continente europea, nell’ottica della conservazione della diversità genetica in specie forestali.
Subgenus Quercus
La diversità cloroplastica delle querce decidue e l’identificazione delle dinamiche di colonizzazione sono state l’oggetto di studio di un progetto europeo (“Synthetic maps of gene diversity and provenance performance for utilisation and conservation of oak genetic resources in Europe”, CT-FAIR1 PL95-0297), che ha coinvolto 12 paesi, compresa l’Italia.
I risultati hanno dimostrato che alcuni tra gli aplotipi (tipi di DNA cloroplastico) che hanno colonizzato il continente europeo si sono originati in Italia, in particolare nelle regioni meridionali e insulari.
E’ interessante notare che la diffusione delle querce che ha interessato la penisola italiana sembra aver seguito un modello di migrazione da sud verso nord e, in minor misura, da est (dai Balcani) verso ovest, mentre non risulta che sia avvenuta migrazione da ovest (penisola Iberica) verso est.
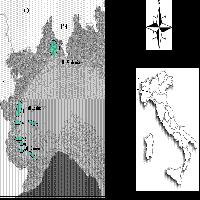
In Fig. 1 è riportata la distribuzione degli aplotipi cloroplastici identificati nella penisola italiana e nelle due isole maggiori, a cui si aggiunge, per motivi geografici, anche la Corsica.
Fig. 1 - Mappa del DNA cloroplastico in popolazioni di querce decidue in Italia e nelle isole maggiori (inclusa la Corsica). Le dimensioni dei singoli cerchi (torte) sono proporzionali al numero di individui campionati in ogni popolazione (min 1, max 5).
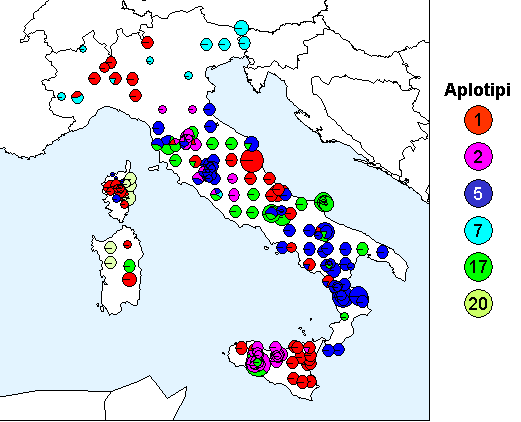
Due aplotipi cloroplastici risultano originari della Sicilia (descritti come aplotipi 1 e 2 da [26]) e sono responsabili della colonizzazione non solo di parte della nostra penisola, ma anche di un’ampia porzione del continente ([7], [12], [25], [26]).
La distribuzione attuale di questi due aplotipi in Italia e il confronto con la situazione europea ci consente di ipotizzare quali possono essere state le modalità di migrazione. Apparentemente, individui caratterizzati da questi aplotipi sono stati in grado di colonizzare le aree appenniniche prima di oltrepassare la barriera alpina migrando verso nord, ma la catena alpina potrebbe essere stata aggirata anziché oltrepassata ([26]).
Le stesse modalità di migrazione possono essere ipotizzate per un altro aplotipo diffuso in Italia (aplotipo 17 secondo [26]), la cui origine risulta difficile da identificare perché potrebbe essere italiana ma anche balcanica.
Un altro aplotipo presente in Italia (aplotipo 5 secondo [26]) potrebbe essersi originato nella parte meridionale della penisola: è opportuno ricordare l’importante area rifugio rappresentata dai Laghi di Monticchio in Calabria, descritta come area di rifugio primario. Si definisce rifugio primario quello localizzato in un’area capace di mantenere l’esistenza di una specie anche durante il periodo di massima estensione glaciale (circa 18000 anni fa). I rifugi primari per le querce sono stati identificati solamente in Europa meridionale ([32]) e uno di questi è l’area dei Laghi di Monticchio. Anche la Sicilia potrebbe rappresentare un’area di rifugio primario, anche se per questa regione non sono disponibili informazioni sui pollini fossili.
Il modello migratorio proposto per le querce caratterizzate dall’aplotipo 5 ([26], [12]) tiene conto della presenza del “ponte adriatico” esistente durante le fasi glaciali, grazie al quale le attuali sponde di Italia e Croazia erano collegate da corridoi terrestri ([30]). Il ruolo che può essere stato svolto dal “ponte adriatico” è sottolineato anche dall’alto numero di specie quercine presenti in Puglia, dove si trovano gli unici popolamenti italiani, che sono anche i più occidentali dell’areale, di Q. macrolepis Kotschy e di Q. troiana Webb ([29]). Queste due specie di querce appartengono alla flora che si suppone sia sopravvissuta in rifugi frammentati nell’Italia meridionale anche durante le fasi glaciali, tra 13000 e 9000 anni fa, che qui erano caratterizzate da una vegetazione di tipo steppico ([17], [29]).
E’ interessante notare che la catena appenninica, in particolare in Calabria, non ha rappresentato un ostacolo alla migrazione di querce con questo aplotipo. Al contrario, non risulta esserci stata migrazione dalla Calabria verso la Sicilia: questo fatto potrebbe essere dovuto dalla profondità del Mar Ionio, che ne ha prevenuto il congelamento anche durante le fasi di massima estensione glaciale, impedendo lo scambio tra le due regioni attraverso un corridoio terrestre.
L’Italia settentrionale è stata colonizzata da uno degli aplotipi siciliani (aplotipo 1) e da un aplotipo che non sembra essersi originato in Italia, ma che probabilmente proviene dal nord dei Balcani (aplotipo 7 secondo [26]). Questo tipo di colonizzazione è comune anche alle regioni alpine degli altri paesi europei ([4]).
Infine appare molto interessante il risultato ottenuto analizzando le popolazioni della Sardegna e della Corsica per la presenza di un aplotipo endemico (aplotipo 20), possibile indicazione dell’esistenza di un’area rifugio in una o in entrambe le isole. Sardegna e Corsica erano congiunte durante le fasi glaciali, pertanto anche la presenza di un unico rifugio può essere stata fonte di colonizzazione per entrambe le isole. La presenza di querce in Corsica alla fine dell’ultimo periodo glaciale è documentata da Reille ([27]), ma questo dato da solo non risulta sufficiente per proporre l’isola come area rifugio.
Alla luce di quanto è stato osservato in Italia, e grazie al confronto che è stato possibile con gli altri paesi europei, possiamo confermare quanto suggerito dalle analisi sui pollini fossili riguardo al ruolo di rifugio svolto dalla penisola italiana e dalle isole maggiori, che ha consentito la conservazione della diversità aplotipica delle querce durante le fasi glaciali, e la loro successiva migrazione.
Subgenus Scleropyllodris
L’analisi del DNA cloroplastico del leccio europeo ha confermato il modello di migrazione già osservato per le querce decidue, da sud verso nord, e ha identificato le tre penisole mediterranee (Iberica, Italiana e Balcanica), come le aree rifugio di questa specie ([20]).
Secondo Huntley & Birks ([17]) le querce decidue erano presenti nel bacino del Mediterraneo durante l’ultimo periodo interglaciale dove sono sopravvissute durante la successiva fase glaciale. La migrazione postglaciale, come testimoniato dagli spettri pollinici, sarebbe iniziata dal Mediterraneo orientale con un ritmo molto rapido, fino a circa 2000 anni fa, quando, in coincidenza con le crescenti attività antropiche, in particolare la deforestazione avvenuta nel Mediterraneo, è registrato una riduzione dei resti pollinici.
La presenza del leccio in Italia è documentata anche durante le fasi interglaciali precedenti a quella attuale ([14], [21]).
Uno studio condotto a livello di tutto l’areale del leccio da Lumaret et al. ([20]) ha messo in evidenza il contributo dato dalle popolazioni italiane alla colonizzazione di alcune aree europee, in particolare della Francia.
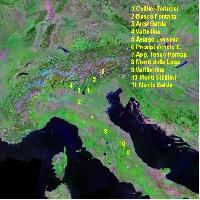
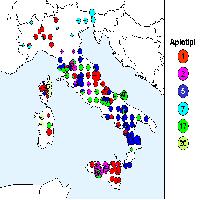
Lo studio delle popolazioni italiane ([13]) ha indicato che la maggior parte della diversità cloroplastica si trova in Sicilia (Fig. 2): degli otto diversi aplotipi identificati in Italia, sei sono presenti sull’isola e cinque di questi (aplotipi 1, 2, 6, 7 e 8) sono endemici ([13]). L’Italia peninsulare è caratterizzata dalla presenza di altri due aplotipi oltre a quello già identificato in Sicilia; mentre uno di questi (aplotipo 5) è diffuso in tutte le regioni, l’altro (aplotipo 4) è limitato alla sola Calabria. La colonizzazione dell’Italia peninsulare sembra avere quindi due diverse origini: la prima come conseguenza di una migrazione dalla Sicilia, l’altra derivante da piccoli rifugi a livello locale che sono ben documentati in tutta l’Italia meridionale e che rappresentano la testimonianza di una più ampia diffusione delle querce sempreverdi durante il precedente interglaciale ([14]).
Fig. 2 - Mappa del DNA cloroplastico in popolazioni di leccio in Italia e nelle isole maggiori. Le dimensioni dei singoli cerchi (torte) sono proporzionali al numero di individui campionati in ogni popolazione (min 1, max 5).
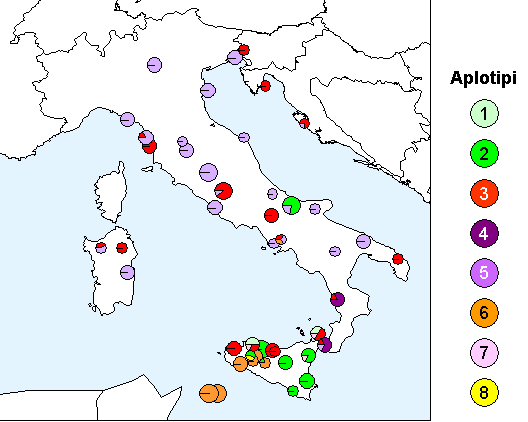
Gli altri aplotipi della Sicilia, che non hanno contribuito alla colonizzazione della penisola, potrebbero rappresentare le ultime testimonianze di un’antica distribuzione del leccio sull’isola, la cui presenza risulta essere stata già cospicua all’inizio dell’Olocene ([28]).
Lo studio sulla diversità cloroplastica del leccio italiano ha consentito di identificare la presenza di due popolazioni non autoctone, in Campania (Avellino) e in Molise (Larino - Fig. 2). Infatti, la presenza di aplotipi siciliani, non rilevati in altre località al di fuori dell’isola, è da attribuire all’introduzione artificiale di materiale di propagazione. In particolare, nel caso di Avellino, l’introduzione di leccio dalla Sicilia è documentato dal botanico Gussone ([23]).
La presenza di sei diversi aplotipi di leccio sull’isola è di supporto all’ipotesi della Sicilia come area rifugio glaciale, come già dimostrato per le querce decidue ([26], [12]). E’ inoltre interessante rilevare che le popolazioni siciliane hanno mantenuto alti livelli di diversità genetica nonostante il forte impatto antropico subito dal paesaggio naturale e forestale dell’isola nel corso degli ultimi millenni.
Conclusione
In base ai risultati ottenuti dall’analisi palinologica e di marcatori molecolari sulle querce decidue e sul leccio, possiamo concludere che alcune regioni della penisola italiana, in particolare quelle meridionali e insulari, sono state aree rifugio (rifugi primari e rifugi secondari) durante i periodi glaciali. Inoltre, queste zone costituiscono un’importante serbatoio di diversità genetica che si è conservata nel tempo nonostante la forte pressione antropica esercitata dalle civiltà che si sono succedute nei secoli nel bacino del Mediterraneo. Una parte di questa diversitàè rimasta circoscritta nelle aree di origine, come testimoniato dalla presenza di aplotipi endemici in Sicilia e in Sardegna e Corsica, mentre una parte consistente di questo patrimonio genetico ha contribuito alla colonizzazione del continente europeo.
Alla luce di questi risultati appare evidente la necessità di potenziare gli sforzi per la conservazione e la protezione di aree che sono importanti bacini di biodiversità, come testimoniano anche la presenza di specie endemiche, sia arboree (come Abies nebrodensis e Zelkova sicula in Sicilia) sia erbacee, nelle maggiori isole italiane.
Questo tipo di studi si basa sull’analisi di marcatori cloroplastici e fornisce informazioni sulla dinamica di migrazione e sulla filogeografia di importanti specie arboree. Tuttavia, questo tipo di informazione deriva solamente dall’esame di marcatori neutrali, i quali non necessariamente descrivono la diversità genetica di natura adattativa. L’obiettivo principale della genetica della conservazione è quello di preservare il potenziale adattativo di una specie attraverso il mantenimento di tutti i livelli di diversità genetica: per raggiungere questo obiettivo è necessaria dunque l’analisi contemporanea di marcatori neutrali e adattativi.
References
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
Online | Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar