Current problems of close-to-nature silviculture in Italy
Forest@ - Journal of Silviculture and Forest Ecology, Volume 1, Pages 59-69 (2004)
doi: https://doi.org/10.3832/efor0232-0001
Published: Dec 28, 2004 - Copyright © 2004 SISEF
Commentaries & Perspectives
Abstract
The main problems of Italian forests are critically reviewed, considering the ecological as well the socio-economic factors that have been responsable of the main evolutionary processes, i.e., the secondary succession under artificial conifer stands, the ageing of the coppice forest, the post-cultivation evolution on abandoned fields and the increase of wild ungulates, which currently represents one of the most dramatic problems to be faced in forest management. Concepts as biodiversity, multifunctionality and sustainable management are the foundations, nowadays, of forest management. The main elements traditionally characterizing the close-to-nature silviculture are discussed and critically examined, warning against ideology and rigidity. Two concepts are more carefully discussed: natural vegetation dynamics and mixed uneven aged forest. An analysis of the priorities of silviculture in Italy concludes the paper, namely: the importance to take advantage of the “natural opportunities” in the cheapest way (the cost of the thinnings is a first rate problem, nowadays); to consider forests ecosystems in the context of surrounding natural and cultural landscape; to define proper management strategies for secondary successions on abandoned fields and pastures; to aim at re-naturalizing artificial conifer stands and increasing/maintaining forest biodiversity; to tackle effectively the wild ungulates problem.
Keywords
close-to-nature silviculture, natural regeneration, forest management
Premessa
Non è intenzione di chi scrive affrontare nel dettaglio aspetti storici dell’argomento trattato, soprattutto perché in proposito è già stato scritto molto (si raccomanda, fra gli altri lavori, La selvicoltura naturalistica nella storia del pensiero forestale, [4]). Vale solo la pena di ricordare che, nell’ambito della selvicoltura naturalistica, in accordo con quanto sostiene Hellrigl (manoscritto inedito) si possono identificare tre nuclei d’origine: uno centro-occidentale (Vosgi, Giura e Selva Nera, dove si è sviluppato il taglio saltuario su piceo-abieteti-faggeti), uno della Germania settentrionale (reso famoso dagli scritti di Moeller sul bosco permanente e basato sul taglio saltuario per pedali, che venne applicato a pinete di pino silvestre in fase di colonizzazione di ex-pascoli), e uno sloveno (basato su criteri di rinnovazione per pedali, nuclei, gruppi e aggregati).
Vale anche la pena di mettere a fuoco le particolarità dell’epoca che i forestali stanno attraversando. Motivi bio-ecologici, ma anche sociali, storici, ideologici ed estetici spingono oggi verso una selvicoltura sempre più attenta alla natura. Nel corso degli ultimi decenni, molte trasformazioni sono avvenute nei boschi del nostro Paese, in gran parte abbandonati alla propria evoluzione: composizione specifica e struttura dei soprassuoli (ma le ricadute sono evidenti su tutte le componenti dei sistemi) ne risultano modificate. Un esempio sono le evoluzioni, orientate verso il bosco misto di latifoglie, dei soprassuoli artificiali di conifere lasciati alla propria evoluzione. Un altro è l’invecchiamento dei cedui, che in assenza di utilizzazioni evolvono più o meno lentamente in direzione di fustaie a struttura irregolare. Un altro ancora è rappresentato dalle evoluzioni di post-coltura di terre abbandonate, con conseguente estensione della superficie forestale a spese di oliveti, vigneti, castagneti da frutto, pascoli. Al quadro fisionomico va aggiunta la forte crescita, in molti territori del Paese, del carico di ungulati selvatici, tendenza iniziata a partire dal II dopoguerra: in alcune aree è uno dei maggiori problemi da affrontare nelle scelte selvicolturali degli anni a venire.
Mentre i boschi si trasformavano, anche la societàè cambiata, e questo si è ripercosso sulla gestione forestale (sebbene sia, in molti casi, più giusto parlare di non-gestione). I motivi dei cambiamenti dei nostri boschi sono sia storico-ecologici sia socio-economici: da un lato l’evoluzione spontanea dei rimboschimenti fatti dai “vecchi forestali” (le fasi successionali cui assistiamo oggi sono conseguenza degli impianti di conifere), dall’altro lo spopolamento delle campagne e delle montagne. Si aggiunga un diverso atteggiamento, da parte dell’opinione pubblica, nei confronti del patrimonio naturale: sviluppo incontrollato, distruzione delle foreste pluviali, erosione genetica e cambiamenti climatici sono all’origine di nuovi timori e di una nuova sensibilità naturalistica. Oggi le parole d’ordine del mondo forestale sono multifunzionalità, biodiversità e gestione sostenibile. La tendenza è di assegnare priorità alla continuità della copertura forestale e di assecondare i processi naturali dei boschi con tecniche eco-compatibili: ne è una conferma il peso acquisito dall’eco-certificazione. La grande crescita degli studi tipologici negli ultimi anni nasce dalla opportunità di interpretare il dinamismo della vegetazione come indagine preliminare alla selvicoltura: non c’è dubbio che tale approccio abbia un senso soprattutto nell’ottica di una selvicoltura su base naturalistica ([11], [18], [23]). Le foreste, inoltre, non sono viste più come ecosistemi autonomi, ma come tessere elementari di un insieme gerarchicamente superiore, il paesaggio, dove ogni ecosistema interagisce con gli altri.
Si commentano ora alcuni pilastri della selvicoltura naturalistica. L’intenzione è di mettere a fuoco i concetti su cui si fonda tale scuola di pensiero, criticando schematismi che appaiono eccessivi.
La selvicoltura naturalistica: breve analisi critica degli elementi che la caratterizzano
1. Tendere alla continuità della copertura del suolo con la rinnovazione naturale . Si tratta, in fin dei conti, delle basi della teoria del bosco permanente di Moeller, che dava grande risalto agli equilibri del bosco nel suo insieme, secondo un approccio organicistico.
Nella selvicoltura naturalistica la rinnovazione naturale è da sempre considerata imprescindibile. Su questo aspetto, tuttavia, è opportuna una riflessione. Anche nell’ottica di una gestione su basi naturali, la rinnovazione artificiale può avere un senso, se il suo significato è quello di integrare la rinnovazione naturale. Viene in mente quanto accade nelle foreste in cui il danno da ungulati impedisce o inibisce fortemente l’evoluzione della vegetazione nel senso di una naturalizzazione (come avviene nella pineta di Pucini presso Camaldoli, in cui il piano delle latifoglie mesofile si attesta attorno a 2-4 m, mentre al di sotto di tale altezza sono pressoché scomparse piante erbaceo-arbustive nonché il novellame di specie arboree, a causa del consumo della selvaggina): in questi casi, assecondare la rinnovazione con piantagioni protette può aiutare a ripristinare un processo naturale, impedito da una attività faunistica innaturale nella sua entità (fatto salvo che le protezioni alle piantine, oltre a essere costose, non possono essere mantenute per lunghi periodi, per cui questo tipo di operazione può essere consigliato in particolari situazioni e su scala limitata). Un altro caso è rappresentato dai rimboschimenti d’alta quota. Per ambienti alpini di quota superiore a 2000 m è stato proposto di eseguire rimboschimenti di abete rosso “per aggregati” ([28]), che consentono sia di sfruttare al meglio la variabilità microstazionale di tali ambienti, sia di creare strutture dotate di grande stabilità nei confronti degli agenti meteorici. La tecnica di rimboschimento, che si basa su osservazioni di modelli di distribuzione naturale dell’abete rosso in ambienti subalpini, nasce dalla considerazione che, soprattutto ad altitudini elevate, la strutturazione in biogruppi andrebbe incoraggiata quale fattore di stabilità.
Artificialità della rinnovazione e impostazione naturalistica convivono anche in questo caso. Ne consegue che a volte può essere opportuno, proprio nella logica di un rispetto degli equilibri naturali, derogare a una delle leggi fondanti della tradizione selvicolturale naturalistica, che è quella di evitarela rinnovazione artificiale.
2. Favorire la formazione e la diffusione di boschi di specie autoctone . Anche su questo assunto si può discutere. Mayr fu il primo a proporre una selvicoltura “su basi naturali” capace di includere le specie esotiche: Mayr metteva al centro della sua concezione selvicolturale la ricchezza di specie, che dovevano essere adatte alla stazione anche se non necessariamente indigene. Pavari riteneva le specie esotiche conciliabili con le esigenze di una selvicoltura naturalistica, sebbene nell’analisi dello studioso entrasse in gioco più che altro la necessità di produzione legnosa: nel contesto di quel tempo si parlava in proposito dell’importanza delle specie esotiche per la realizzazione di una selvicoltura “autarchica” ([24]). Ma anche oggi esistono situazioni in cui non si può escludere a priori l’impiego di specie esotiche nell’ambito di una gestione naturalistica: di esse viene sottolineata l’importanza non solo sotto il profilo economico, ma anche in quanto elementi di arricchimento di biodiversità ([15]). Naturalmente, in una selvicoltura prossima alla natura, l’impiego di rinnovazione artificiale di specie esotiche è soggetta a restrizioni: le specie introdotte devono avere provata adattabilità alle condizioni ambientali, capacità di rinnovarsi per via naturale (senza sopprimere la rinnovazione delle specie native), e inoltre non devono impoverire il suolo né rendere il soprassuolo più suscettibile a danni meteorici, e nemmeno favorire la diffusione di patogeni. A patto che si verifichino queste condizioni, l’impiego di specie esotiche nella selvicoltura prossima alla natura trova giustificazione in una interpretazione della gestione forestale in cui i processi vengono privilegiati rispetto alle strutture originarie ([15]). Infatti, se è vero che l’introduzione di specie esotiche ha, soprattutto in ambienti semi-naturali e su piccola scala, limitato la biodiversità di alcuni ecosistemi forestali (determinando, a esempio, la scomparsa di specie rare del sottobosco), è pure vero che tale introduzione ha permesso di colmare lacune ecologiche: in Britannia settentrionale, la scomparsa di Ulmus glabra (la principale specie arborea tollerante l’ombra in quel settore geografico) è stata compensata dalla introduzione di faggio e acero di monte, che avendo una buona tolleranza dell’ombra hanno finito per occupare proprio la nicchia lasciata libera dall’olmo (Peterken 2000). Per quanto riguarda la selvicoltura italiana, viene in mente non solo il ruolo svolto dal cipresso in Toscana, ma anche quello del cedro dell’Atlante nella Sardegna interna. L’introduzione della conifera montano-mediterranea ha dato buoni risultati sul piano sia paesaggistico sia ecologico: da un lato l’intrusione del cedro è accettata dalla popolazione locale, dall’altro la specie è in fase di naturalizzazione, in quanto capace di rinnovarsi sia all’interno dei popolamenti più vecchi sia allo scoperto ([2]).
3. Ponderare gli interventi sulla base di valutazioni a scala differenziata. Tale opportunità nasce dall’osservazione che il bosco, in molti casi, è un mosaico di situazioni stazionali e strutturali, da gestire con interventi selvicolturali specifici secondo i singoli tratti implicati (quelle che potremmo chiamare le tessere del mosaico). Il dinamismo delle foreste vergini centro-europee ha rappresentato, in questo senso, una grande lezione.L’insieme delle fasi successionali in cui comunemente si distingue il dinamismo delle foreste vergini esprime un ciclo temporale durante il quale i popolamenti arborei caratterizzati da diversa composizione specifica e struttura si succedono sulla stessa superficie. A livello spaziale, nel complesso forestale il ciclo si apprezza con una distribuzione a mosaico delle varie fasi ([17]). Fra le applicazioni selvicolturali dell’analisi della dinamica delle foreste naturali, si segnala quella proposta per le foreste del Comelico, in provincia di Belluno ([3]). Infatti, anche nei boschi coltivati si possono individuare tessere con problematiche differenti, cui assegnare interventi selvicolturali specifici: la fase di rinnovazione è segnata da una forte competizione con le specie erbaceo-arbustive e da fenomeni di predazione da parte di ungulati, quella di costruzione da una spiccata concorrenza intra- e interspecifica, quella di biostasi è invece legata a interventi di utilizzazione, che a loro volta innescano fenomeni successionali nel bosco, mentre la fase di decadenza scompare se con il taglio si asportano alberi adulti prima del loro naturale deperimento. Il grande insegnamento che si trae dalle foreste vergini è che ogni tratto di bosco necessita di un approccio a parte, legato a fattori stazionali (suolo, giacitura ed esposizione), strutturali (composizione specifica struttura spaziale dei soprassuoli), vegetazionali (velocità di accrescimento e capacità di tolleranza della copertura da parte delle piante presenti nel piano di rinnovazione) e storici (età stadiale). Ne consegue che uno dei fondamenti della selvicoltura naturalistica è l’elasticità nelle forme di trattamento: taglio raso, tagli successivi più o meno uniformi, tagli saltuari e forme di trattamento intermedio possono coesistere nello stesso bosco, purché vengano applicati di volta in volta con intensità ed estensioni da calibrare secondo aspetti stazionali e dinamici. Vale la pena di sottolineare che nemmeno i tagli rasi - che pure non garantiscono uno dei principali requisiti tradizionali della selvicoltura su basi naturali, cioè la continuità della copertura arborea del suolo - dovrebbero essere esclusi a priori: niente di così paradossale, se si pensa che incendi, agenti meteorici e insetti sono espressioni della natura capaci di creare ampie aperture nei boschi, con effetti analoghi a quelli che avrebbero i tagli rasi! Già nel 1878, nel suo trattato di selvicoltura, Gayer sosteneva che il taglio raso non andava necessariamente evitato, ma che si trattava solo di limitarne le dimensioni e di studiarne i casi di applicazione. Su posizioni analoghe si pongono anche autori recenti ([33], [6]). Bernetti sottolinea come le tagliate a raso possano rappresentare una fonte di biodiversità sotto vari profili e a vari livelli di scala. Facendo l’esempio di Vallombrosa, l’autore mette in rilievo la ricchezza di specie erbacee, arbustive e arboree presenti nelle fasi iniziali delle successioni nelle tagliate, evidenziando le ricadute positive di queste sull’avifauna e sul paesaggio. Resta il fatto che (corollario della concezione del bosco inteso come mosaico di situazioni stazionali e strutturali), come regola generale, la selvicoltura su basi naturali dovrebbe basarsi su interventi non intensi, su piccole superfici e dilazionati nel tempo. Soprattutto, siccome si parte sempre da un bosco esistente, con le sue tendenze dinamiche in atto, e non da un modello teorico, ogni bosco va visto separatamente dagli altri, in quanto dotato di una propria storia ecologica e selvicolturale.
4. Il bosco misto a struttura disetaneiforme . Si tratta di uno dei luoghi più comuni della selvicoltura naturalistica: non nel senso del modello in se, ma per le pericolose generalizzazioni cui è soggetto. Infatti, una fisionomia del genere non sempre è naturale e, soprattutto, compatibile con l’ambiente (substrato e clima). Fra l’altro, quando si parla di mescolanze e di disetaneità, come minimo è necessario fare riferimento a una scala (una cosa è la disetaneità riferita a una superficie di 2000 m2, altra cosa è la disetaneità a scala di qualche ettaro).
Anche se, nella fascia medio montana e su suoli evoluti, strutture multiplane si osservano in alcuni tratti di territorio appenninico (vedi foreste casentinesi del versante romagnolo), in Italia la maggiore frequenza di boschi con struttura multiplana si registra nelle Alpi.L’origine di queste strutture, e della loro distribuzione su scala geografica, chiama in causa fattori bioecologici e storici.
Strutture complesse si originano soprattutto quando il grado di copertura arborea consente forme di dinamismo accentuato: si possono citare le successioni su coltivi e pascoli abbandonati (soprattutto quelle che avvengono con lentezza e gradualità, vuoi per rigore del clima vuoi per il lento accrescimento delle specie in ingresso: è il caso di colonizzazioni di picea in pascoli e lariceti-pascoli di fascia subalpina), come quelle che avvengono sotto la copertura di impianti di conifere e di castagneti da frutto lasciati alla propria evoluzione.
Non tutte le specie forestali sono adatte a formare consorzi misti a struttura multiplana. L’abete bianco, per esempio, ha questa tendenza perché si rinnova per piccole chiazze di novellame, perché tollera l’ombra e perché, una volta arrivato allo stadio di pianta adulta, la sua chioma non è invadente. Ma il comportamento di una specie varia anche con la stazione. Hofmann (in [12]) osserva che in Friuli l’abete bianco può dar luogo a popolamenti misti pluristratificati, perché essendo nell’ottimo si rinnova bene sotto copertura, al contrario di quanto avviene nell’Appennino meridionale, dove non a caso prevalgono strutture monoplane. D’altro canto il faggio, specie dominante nella montagna appenninica, presenta caratteristiche di rinnovazione e di accrescimento giovanile tali da far sì che i boschi tendano alla monoplanarità già in giovane età.
Per quanto riguarda gli aspetti storici, non si può fare a meno di ricordare che i boschi misti disetanei del Trentino e del Cadore sono legati a doppio filo a forme di trattamento finalizzate a un preciso modello di sfruttamento del bosco. La necessità, nata nell’ambito della piccola azienda, di tagliare con continuità, in modo da avere un reddito più frazionato e una maggiore elasticità gestionale ([26]), di quei boschi spiega forse più cose di quante ne spieghi la natura.
All’atto pratico, nel nostro Paese la gestione della foresta secondo i modelli del bosco disetaneo ha trovato corretta applicazione nei popolamenti delle Alpi che manifestano tendenza strutturale verso il bosco misto e pluristratificato di abete rosso, abete bianco e faggio, mentre clamorosi insuccessi si sono registrati quando si è voluto generalizzare, forzando mescolanze e disetaneizzazioni negli ambienti sbagliati (tipo le faggete montane dei substrati carbonatici). Come dice Hofmann (in [12]), “i trasferimenti di modelli strutturali da una regione all’altra, sia pure per una stessa specie legnosa e per uno stesso consorzio misto, vanno affrontati con una certa prudenza ”.
Fino a che punto una struttura disetaneiforme può essere considerata stabile? Per rispondere a questo quesito torniamo alle foreste vergini del piano medio montano centro-europeo: qui la fase multiplana dei soprassuoli può essere durevole in situazioni a forte microvariabilità stazionale, oppure dove un importante fattore ecologico condiziona il ricambio generazionale. La veritàè che una fase multiplana, in natura, di regola è transitoria. Come osserva Hellrigl (manoscritto inedito), la monoplanarità su ampie superfici esisteva in molte foreste primigenie del nostro continente, in particolare dove una specie si trovava nell’ottimo ecologico (e quindi manifestava maggiore forza concorrenziale, sia per accrescimento che per sopravvivenza), dove era elevata la frequenza di eventi catastrofici (uragani, frane, incendi, valanghe) e nella fase ottimale dei consorzi dei climi temperato-freddi.
Il concetto vale anche per le foreste coltivate: in modo particolarmente veloce sui suoli più fertili e in assenza di tagli di curazione, col tempo la tendenza di molti boschi disteanei del Trentino è verso strutture coetaneiformi, per ingresso nel piano superiore delle piante delle nuove generazioni ([31]). Osserva Hellrigl (inedito) che, taglio raso o taglio saltuario che sia, si tratta pur sempre di trattamenti che interrompono a un certo livello il ciclo naturale proprio delle foreste primigenie: i trattamenti “monoplanarizzanti” su ampie superfici simulano un ricambio generazionale da catastrofe o da decadenza senile accelerata, e interrompono il decorso delle fasi nella fase ottimale; i trattamenti “multiplanizzanti”, d’altro canto, soprattutto nel caso di tagli successivi a gruppi, seguono la via di ricambio più frequente nel bosco vergine, ma cercano di perpetuare una struttura che in natura è transitoria. La conclusione è che il bosco puro monoplano può anche essere naturale, mentre quello misto multiplano non lo è necessariamente. Pecceta subalpina, faggeta di quota e pineta di pino nero su calcari confermano tale convinzione.
In ultima analisi, il bosco misto non deve essere una priorità assoluta, anche se nella fascia medio montana e su suoli di discreta fertilità pare opportuno andare in tale direzione. Secondo una scuola che fa capo agli studiosi di foreste vergini Leibundgut e Mlinsek, il modello selvicolturale nella fascia medio montana sarebbe rappresentato da tagli successivi irregolari (a gruppi e con lungo periodo di rinnovazione, inquadrabili come forme di trattamento combinate fra taglio saltuario e tagli successivi), da cui risultano strutture disetanee a gruppi, con diverso grado di stratificazione.
5. Collegato a quello sul bosco misto è il discorso sulla biodiversità , concetto tuttavia ben più ampio di quello di bosco misto. La biodiversitàè inclusa nel contenitore della complessità dell’ecosistema, dei suoi equilibri e delle sue relazioni con gli altri sistemi: vi entrano perciò in gioco non solo la composizione specifica del soprassuolo arboreo, ma molte altre componenti, da quella microbiologica a quella animale, da quella strutturale a quella paesaggistica ([22]).
Sotto il profilo della composizione specifica delle piante arboree, la selvicoltura naturalistica europea è oggi indirizzata: a) verso la conservazione di foreste miste a struttura complessa; b): verso il “restauro” dei boschi [Nella selvicoltura centro-europea, in particolare per monocolture di pino silvestre, la naturalizzazione acquista il significato di recupero di situazioni deteriorate in seguito al coniferamento di antichi boschi di latifoglie: il recupero in atto si manifesta attraverso un incremento del contenuto di azoto nel suolo, a sua volta conseguenza dell’ingresso delle latifoglie sotto copertura (accumulo autogenico di humus, [34])], favorendo il ritorno delle latifoglie sotto la copertura delle conifere di introduzione artificiale, in modo da ridurne la dominanza e da favorire una crescita di diversità vegetazionale e strutturale. Si tende cioè a mantenere la diversità dove questa è alta, e a incoraggiarla a crescere negli ecosistemi semplificati di origine artificiale (pinete e abetine da rimboschimento ecc.), ma solo dove le condizioni stazionali lo consentono.
Va aggiunto che nel rispetto della biodiversità rientra anche il rispetto delle specie (e delle nicchie) rare. La selvicoltura naturalistica, infatti, deve essere attenta a ogni dettaglio del sistema, come il sottobosco e gli humus (indicatori di fertilità e di attitudine alla rinnovazione naturale ([29])), le radici morte, di cui è stata sottolineata l’importanza come fattore nutrizionale ed energetico nei sistemi forestali ([30]), o gli individui secchi o marcescenti, a terra o in piedi, nicchie adatte per accrescere o mantenere la biodiversità di uccelli, insetti e funghi saprofiti
Per quanto riguarda il legno morto, se il principio generale è condivisibile, non se ne può fare tuttavia una regola fissa. Nei boschi più antropizzati, il legno morto può rappresentare un focolaio per insetti, capaci di attaccare gli alberi vivi: se nella riserva integrale di Sasso Fratino ha un senso tenere in piedi alberi marcescenti, pare non avere molto senso fare altrettanto nelle abetine di Vallombrosa e in quelle dell’Aspromonte, minacciate rispettivamente da Cryphalus piceae e Pityokteines spinidens, agenti responsabili di vistosi fenomeni di deperimento ([25]). Anzi, in sistemi forestali come quello di Vallombrosa, con elevato grado di artificialità e dunque fragili (fragilità aumentata dalle ricorrenti crisi di aridità), sarebbe quanto mai opportuno eliminare tempestivamente i focolai di infezione.
6. Assecondare la multifunzionalità del bosco (protezione di suolo e clima, biodiversità, equilibri biologici, produzione, paesaggio, turismo, ecc.), compresa la funzione di produzione legnosa. In uno scritto di quasi quarant’anni fa, de Philippis, parlando dell’“indirizzo naturalistico della selvicoltura”, sottolinea come gli interventi vadano subordinati alla valutazione degli effetti che essi producono sull’equilibrio bio-ecologico degli ecosistemi forestali ([13]). In ultima analisi si sottolinea che, in selvicoltura naturalistica, il prelievo legnoso deve essere compatibile con l’obiettivo primario delle cure al bosco (in altre parole, l’utilizzazione è contemporaneamente un intervento colturale). La selvicoltura naturalistica non ha mai separato la produzione legnosa dalle altre funzioni del bosco: se c’è una specificità della selvicoltura naturalistica è proprio questa. Semmai, sotto il profilo della produzione legnosa, la selvicoltura naturalistica è particolarmente attenta a sfruttare, sulla scia di quanto a suo tempo sostenuto da Gournaud, le potenzialità produttive locali (microstazionali) e individuali (del singolo albero). Quello della multifunzionalità del bosco è un vecchio principio sempre valido, che oggi è giusto integrare con la presa di coscienza che il bosco è inserito in una rete di ecosistemi interagenti fra loro: come già accennato, il punto di riferimento della selvicoltura deve essere un livello di scala superiore al bosco (il paesaggio).
Alcune puntualizzazioni sul concetto di selvicoltura naturalistica
Dopo avere esaminato criticamente la serie di punti fermi su cui tradizionalmente si appoggia la selvicoltura naturalistica, è doverosa una considerazione: la selvicoltura naturalistica non è una filosofia da contrapporre ad altri approcci di taglio non-naturalistico.
La selvicoltura naturalistica è solo uno strumento con cui si cerca di assecondare il dinamismo della vegetazione in senso “naturalistico” (non sempre le dinamiche spontanee delle foreste sono orientate in tal senso: basta pensare alle invasioni di Robinia pseudoacacia e di Prunus serotina nei querceti del Parco del Ticino!), “una selvicoltura libera, che renda giustizia alle particolarità stazionali e all’assetto temporaneo del bosco un fenomeno unico nello spazio e nel tempo, per la varietà di stazioni che occupa, per la mutevolezza degli eventi naturali occasionali, climatici o patologici” ([16]). Detto questo, la selvicoltura naturalistica è nient’altro che un’opzione, che deve trovare applicazione nei casi che le si confanno. Una selvicoltura non prossima alla natura può comunque avere un senso, e può essere utile alla società umana sotto il profilo economico senza per questo risultare sempre dannosa sotto il profilo ecologico. Né, tanto meno, ha senso dire che una selvicoltura su basi non-naturali non sia una selvicoltura degna di questo nome. Si prenda a esempio la foresta di Vallombrosa. Nonostante l’artificialità del trattamento cui tale complesso è stato soggetto per secoli, gli impianti hanno svolto e tuttora svolgono una funzione multipla tipica dei popolamenti forestali: produttiva (legno, selvaggina, funghi), idrogeologica (copertura arborea continua e stratificata, da parte delle chiome degli abeti, delle specie arboree in ingresso e del sottobosco erbaceo-arbustivo, cui si aggiunge il filtro di un suolo forestale maturo), naturalistica (alto contenuto di biodiversità vegetale e animale), ricreativa e paesaggistica. La selvicoltura, in ultima analisi, “si applica a boschi, anche se in vario grado artificiali, creati e destinati a essere trattati con criteri e finalità forestali” ([13]).
Un’altra questione è quella delle espressioni lessicali. Nella letteratura forestale di lingua inglese e tedesca, ma anche italiana, compaiono termini come “selvicoltura secondo natura”, “selvicoltura vicino alla natura”, “selvicoltura delle opportunità naturali” (libera traduzione di una espressione dello svizzero Schütz: [10]), spesso con la dichiarata intenzione di esprimere approcci alternativi a quello della tradizionale “selvicoltura naturalistica”. Quello delle espressioni lessicali è un problema che nasce in buona parte dalla difficoltà di accordarsi non tanto sul significato del termine naturalistico, quanto addirittura su quello di natura, concetto interpretato in relazione ad aspetti storici e geografici: chi scrive è stato recentemente illuminato da un colloquio con Bernetti, che non ha esitato a sottolineare come Biolley, Moeller e perfino gli studiosi di foreste vergini (che hanno spesso fatto le loro osservazioni su limitate particelle di bosco) abbiano indicato come modelli di bosco naturale le poche decine o centinaia di ettari di foreste su cui di volta in volta hanno concentrato la loro attenzione.
Ma le espressioni lessicali, e i concetti che esse esprimono, non sono il nocciolo della questione che qui interessa affrontare. A parere di chi scrive, è la cosiddetta sostenibilità ecologica, il nocciolo della faccenda. E proprio sulla gestione sostenibile vale la pena di ribattere, perché i nostri boschi è di questo che hanno necessità. E allora non si può fare a meno di sottolineare che nel nostro Paese, oggi, sostenibilità significa soprattutto fare i conti con la componente faunistica dei sistemi forestali. Nell’ottica di una gestione su base naturalistica, il problema degli ungulati selvatici, che minacciano la rinnovazione naturale di molti boschi italiani, è prioritario. Alte densità di ungulati selvatici possono determinare danni che vanno al di là di un semplice ritardo del processo successionale, e possono addirittura modificare strutture e composizioni specifiche dei soprassuoli. Il problema acquista particolare rilevanza nelle foreste montane e alto-montane, visti i lunghi periodi necessari all’affermazione della rinnovazione naturale ([19]). Al contrario di quanto si pensa comunemente, non si tratta solo di un problema di carico faunistico. I nostri boschi avrebbero bisogno di veri e propri piani di assestamento faunistico, che tengano in debito conto non solo il carico complessivo, ma anche le distribuzioni in classi cronologiche e le gerarchie di gruppo ([8]). Per quanto riguarda il selvicoltore, è bene che esso sia consapevole che l’attrattiva di un habitat per la selvaggina non dipende solo dalla disponibilità di cibo, ma anche, e in notevole misura, da fattori ambientali indipendenti dal cibo, quali condizioni del terreno, clima, effetti di margine, impatto di competizione e di disturbo, fattore termico, ricovero e nascondiglio. E che in più di un caso le maggiori densità di selvaggina non si riscontrano nelle aree con maggiore disponibilità di cibo, ma in quelle più ricche di margini, che esprimono (almeno per i cervidi) una grande forza attrattiva. Per quanto riguarda la disponibilità di cibo, la selvicoltura orientata verso il bosco misto accresce l’abbondanza di giovani alberi originati da rinnovazione naturale, diversifica l’offerta alimentare e riduce perciò la predisposizione al danno da brucamento e da scorticamento ([27]). In questo senso, si rifletta sul fatto che, fra le cause che negli ultimi decenni hanno scatenato il problema dei danni da ungulati selvatici in molte foreste europee, si segnala non solo l’assenza di grandi predatori, ma anche una selvicoltura che fino agli anni ’70 ha puntato in gran parte su boschi monospecifici. Un aumento della disponibilità alimentare si registra anche in tutti i boschi sottoposti a cure colturali: una densità ridotta dei soprassuoli consente lo sviluppo di un piano erbaceo-arbustivo, fonte di nutrimento per la fauna ungulata. D’altronde non si tratta solo di un problema selvicolturale: forestali, cacciatori, agronomi e autorità delegate alla gestione del turismo, sono tutti soggetti da coinvolgere, visto che il problema va affrontato su scala territoriale. Non si possono sottovalutare le migrazioni stagionali, come il fatto che se viene creata un’area protetta dalla caccia, gli ungulati tendono a concentrarsi proprio lì. Nel Parco Nazionale delle Foreste Casentinesi, per esempio, molti animali sottoposti alla caccia sono indotti a rifugiarsi all’interno dei confini dove questa è vietata ([14]).
Campi di applicazione della selvicoltura naturalistica attuale
Nel nostro Paese la selvicoltura naturalistica si è sviluppata quasi esclusivamente sulle Alpi. Anche al di fuori della tradizionale cornice esiste oggi, in realtà, più di una opportunità in tal senso. Un esempio sono le terre abbandonate. Che si tratti di castagneti da frutto, oliveti, vigneti, seminativi o pascoli, l’abbandono delle terre ha portato all’espansione di boschi e arbusteti. Fra questi ultimi, i pruneti svolgono un ruolo ecologico importante, sotto il profilo sia del miglioramento delle caratteristiche fisiche del suolo (le radici ne aumentano la porosità ed intaccano eventuali orizzonti induriti, tipo la classica soletta), sia della biodiversità (in quanto habitat di rifugio, alimentazione e riproduzione di uccelli e mammiferi): la loro conservazione è perciò opportuna soprattutto dove la fauna è un patrimonio da tutelare. Ma il discorso sui rimboschimenti naturali va ben al di là degli arbusteti: molte terre abbandonate stanno subendo una evoluzione forestale, e avrebbero necessità di essere gestite, soprattutto nelle situazioni più critiche. Dato che la copertura forestale di regola esercita un’azione positiva nei confronti dell’erosione e del ciclo dell’acqua, l’efficacia idrogeologica del rimboschimento naturale dovrebbe essere fuori discussione. In realtà, l’abbandono delle colture è spesso legato all’abbandono di sistemazioni (terrazzamenti e muretti a secco), il cui degrado può comportare - in corrispondenza di pendenze particolarmente elevate - conseguenze negative, in grado di controbilanciare l’azione positiva del bosco: chi scrive ha osservato più di una situazione problematica negli oliveti delle colline fiorentine. La gestione dei rimboschimenti naturali, fra l’altro, può avere importanti ricadute sul piano economico, soprattutto nel caso che le specie d’ingresso presentino legno pregiato. In ambiente alpino (in cui è frequente l’invasione dell’aceri-frassineto) o in alcune zone appenniniche (in cui si registra l’ingresso del ciliegio o della rovere) può valere la pena di intervenire in modo da valorizzare tali specie.Analoghe considerazioni valgono per certi castagneti da frutto abbandonati, come quelli di molte zone casentinesi, che presentano un piano successionale di latifoglie a legno pregiato. Questi popolamenti meritano di essere assecondati nella loro evoluzione, non solo per motivi naturalistico-paesaggistici, ma anche in prospettiva di una produzione legnosa di qualità ([23]).
Collegato al problema dei rimboschimenti naturali è quello delle modifiche paesaggistiche. Il discorso è piuttosto complesso, e va ben al di là del luogo comune secondo il quale una gestione indirizzata verso “boschi più naturali” comporterebbe sistematicamente arricchimenti sotto il profilo estetico-paesaggistico. Le scelte di pianificazione e di gestione del paesaggio possono essere guidate anche da criteri estetici, che non coincidono necessariamente con quelli naturalistici (Lucas 1991). E’ noto, del resto, che sotto il profilo estetico, le pinete di pino domestico, i castagneti da frutto e i lariceti-pascoli (che notoriamente hanno ben poco di naturale) sono tra i preferiti dai fruitori del territorio. Il rimboschimento naturale, in realtà, ha in più di un caso determinato vistose modificazioni di tradizionali cornici paesaggistiche, con perdita di diversità ad ampia scala, imputabile alla scomparsa di tessere elementari un tempo assai ben connotate al loro interno. Se la biodiversità a livello di singola coltura tende a crescere con i tempi di abbandono (crescita dei taxa vegetali e animali, come conseguenza del passaggio da una monocoltura a un popolamento misto), quella ad ampia scala tende a decrescere: questa omogeneizzazione è la tendenza attuale dell’evoluzione paesaggistica del nostro Paese ([32], [1]). Soprattutto quando lo scopo sia quello di conservare e rivalutare il paesaggio tradizionale, è necessario affrontare il problema della diversità andando oltre gli aspetti legati alle singole tessere, arricchendo le valutazioni di componenti culturali, estetiche e sociali. Per la Toscana, alcuni esempi di paesaggi culturali sono i castagneti da frutto della Montagnola senese, delle Colline Metallifere livornesi e dell’Isola d’Elba, le abetine pure appenniniche e le pinete litoranee di pino domestico. Con loro se ne andrebbe parte della memoria delle zone che li ospitano, qualcosa di molto rilevante in termini di diversità culturale. Insomma, una selvicoltura su basi naturali, finalizzata ad assecondare il dinamismo spontaneo della vegetazione, in certi casi può avere effetti poco apprezzabili nei confronti della diversità paesaggistica. Su questo vale la pena di riflettere, perché la diversità in questione non ha solo un significato estetico e culturale, ma anche un valore ecologico. Basta pensare alle conseguenze sulla fauna selvatica. E’ noto che la scomparsa di spazi aperti, oggi rimboschiti per via naturale, ha determinato una riduzione dell’offerta alimentare, con ricadute sul prelievo in bosco. Parlando non tanto di selvicoltura, quanto di gestione, diviene necessario allargare l’approccio a un livello di scala maggiore: ci si accorgerà che, paradossalmente, proprio in un’ottica di rispetto degli equilibri naturali, alcune tessere elementari potrebbero essere gestite con criteri non strettamente naturalistici.
Sul paesaggio sono peraltro evidenti le conseguenze dell’abbandono delle pratiche selvicolturali: quelle più vistose sono l’invecchiamento dei cedui e la trasformazione di boschi puri di conifere in boschi misti con latifoglie, fenomeno che sta modificando composizione specifica e struttura dei popolamenti, con ricadute su forme e colori dei paesaggi forestali.
La questione si complica quando nella gestione dei boschi di origine artificiale entrano in gioco aspetti sia estetici sia naturalistici. Per esempio, la conservazione delle pinete di pino marittimo sulle serpentiniti dei Monti Rognosi in alta Valtiberina (provincia di Arezzo) è consigliabile proprio in considerazione della particolare fisionomia che i popolamenti in questione assumono (radi e poco sviluppati in altezza), mentre una selvicoltura finalizzata alla eliminazione del pino è caldeggiata da chi punta sulla conservazione della vegetazione arbustiva tipica della steppa-gariga ofiolitica, che l’invadenza del pino sta minacciando (Chiarucci et al. 1995).
Vale ora la pena di fare un breve esame su alcuni casi più tipicamente forestali. In questo senso, un problema cruciale è quello delle conversioni dei cedui in fustaie. Il problema è troppo ampio per affrontarlo in poche righe - si rimanda alla recente trattazione di Ciancio & Nocentini ([9]) - anche se vale la pena di sottolineare che la questione andrebbe analizzata caso per caso, tenendo conto non solo delle condizioni ecologiche, ma anche (soprattutto, verrebbe voglia di dire) delle implicazioni socio-economiche, visto che i diradamenti finalizzati alle conversioni sono nella maggioranza dei casi sconsigliabili in quanto troppo costosi.
Qualche parola in più si può spendere invece sui boschi di conifere, facendo riferimento ad alcuni esempi del territorio toscano. Le pinete di Pinus pinaster delle Cerbaie di Fucecchio e del Monte Serra sono ormai gravemente minacciate da Matsucoccus feytaudi. L’unica soluzione selvicolturale al problema è rappresentata da un incoraggiamento all’ingresso delle latifoglie, con progressiva eliminazione del pinastro, che al momento (al di là del valore paesaggistico della specie), nelle zone di infestazione non pare avere alcun futuro. Si tratta di far affermare gruppi di rinnovazione a scapito di pini deperienti: leccio, corbezzolo, orniello, cerro, castagno, rovere e robinia rappresentano un complesso in cui è necessario controllare l’invadenza della robinia e, in più di un caso, anche di felci, scope e rovi.
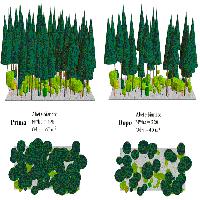
L’ingresso delle latifoglie si registra anche nelle pinete di impianto artificiale di Pinus nigra. Il presupposto per la loro rinaturalizzazione è rappresentato dall’innalzamento delle chiome dei pini e da interruzioni della copertura arborea: è dimostrato che le pratiche selvicolturali accelerano l’evoluzione dei popolamenti ([5], [20]). Il modo di operare si basa su una progressiva riduzione della copertura del pino, con interventi colturali assimilabili a diradamenti, di intensità tanto maggiore quanto più il piano di rinnovazione è affermato. Per le pinete eutrofiche del Casentino, nei casi in cui il piano successionale sia ben sviluppato, il modulo colturale (Fig. 1) potrebbe basarsi su due o tre interventi di alleggerimento della copertura della conifera, in modo da arrivare a un bosco biplano, con 50-70 grosse piante emergenti dai piani inferiori ([18], [7]). Alla fine, le piante di pino potrebbero essere conservate fino a mortalità naturale, oppure abbattute in modo da completare la trasformazione nel bosco di latifoglie, nella cui composizione specifica entrano tiglio, frassino maggiore, olmo montano e acero montano.
Fig. 1 - Esempio di intervento finalizzato ad alleggerire la copertura del pino nero in una pineta casentinese, in modo da favorire l’affermazione delle latifoglie mesofile (castagno, cerro, acero di monte, acero riccio, acero opalo, frassino maggiore, orniello, tiglio, olmo montano, carpino bianco) nel piano inferiore. Le piante segnate con una croce sono quelle che cadono al taglio. Simulazione e grafica a cura di Livio Bianchi.
Le abetine pure appenniniche (come quelle di Vallombrosa e di Camaldoli) sono un altro caso di grande attualità. E’ noto che, a partire dai primi anni ’70, le tradizionali pratiche selvicolturali, adottate a suo tempo dalle comunità benedettine, sono state abbandonate: come conseguenza, l’ingresso di latifoglie (soprattutto faggio, castagno, aceri e tigli) ha avviato una evoluzione delle abetine pure monoplane in boschi misti a struttura complessa. La selvicoltura può assecondare il dinamismo naturale: i tagli dovrebbero interessare strisce di bosco adiacenti ai gruppi di rinnovazione, in modo da creare orli esterni nei quali le condizioni per l’affermazione del piano successionale siano più favorevoli. E’ una selvicoltura che richiede l’abbandono del concetto di turno e di diametro di recidibilità, nonché una frequenza di intervento alta (es. ogni 5-10 anni), e che può ritenersi piuttosto impegnativa.
In Toscana esistono anche casi di abetine miste con latifoglie, legate alla tradizione francescana e diffuse, oltre che alla Verna (Casentino), nel Monte Amiata: è il caso del Bosco della SS. Trinità presso Santa Fiora, caratterizzato dalla mescolanza dell’abete bianco con faggio e latifoglie mesofile (tigli, frassino maggiore, cerro, castagno, aceri, carpini, ecc.). La gestione dovrebbe essere finalizzata al mantenimento della elevata diversità della fitocenosi. La necessità di creare margini utili alla rinnovazione delle varie specie è il punto cardine della selvicoltura proposta, che non si discosta molto da quella suggerita per le abetine pure in evoluzione ([21]).
Conclusioni
L’economicità degli interventi selvicolturali è ormai un problema di primo piano. Fino a cinquant’anni fa, la selvicoltura si sviluppava in un quadro in cui gli interessi della produzione legnosa erano preminenti, mentre il costo del lavoro era basso; oggi le cose sono cambiate, per cui è necessario sfruttare “le opportunità naturali” in modo da “massimizzare contemporaneamente produzione di massa e qualità” nel modo meno costoso: cito una frase tratta da Schütz ([10]), perché esprime bene la scommessa della selvicoltura dei nostri tempi. Gestire i boschi, valorizzandone la produzione legnosa con spese di intervento contenute: è un obiettivo difficile da raggiungere, soprattutto perché la selvicoltura naturalistica richiede un’attenzione e una frequenza di tagli che appaiono in contraddizione con la necessità di contenere le spese. Ma vale la pena di affrontare la sfida, anche perché l’alternativa è l’abbandono del patrimonio forestale a se stesso.
Vengono in mente le parole di un altro vecchio maestro della selvicoltura italiana ([16]), il quale non si stancava di ripetere che affidare un bosco antropizzato alle forze della natura non significa ricostituire un consorzio primitivo: “può sembrare una contraddizione in termini, ma per ottenere boschi secondo natura, si deve intervenire artificialmente, con l’accetta”.
References
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar