Application of isotope discrimination techniques to evaluate the functional response of Mediterranean coppices to high-forest conversion cut
Forest@ - Journal of Silviculture and Forest Ecology, Volume 2, Pages 367-377 (2005)
doi: https://doi.org/10.3832/efor0313-0020367
Published: Dec 19, 2005 - Copyright © 2005 SISEF
Research Articles
Abstract
Discrimination of stable isotopes of carbon and hydrogen provides an effective tool for interpreting time-integrated responses of plants to environmental conditions and delineate sources of plant water uptake. In this work, isotopic analyses were carried out in two Mediterranean oak forests in which a thinning experiment on replicated plots has been performed. Changes in carbon isotope discrimination suggests an increase of water use efficiency soon after thinning. Together with changes in the hydrogen isotopic composition in xylem sap, this may suggest that trees are able of a rather prompt physiological acclimation to cope effectively to new environmental conditions and changes in resource availability.
Keywords
Stable isotopes, Carbon, Hydrogen, Water-use efficiency, Coppice, Silviculture, Treatment
Introduzione
La gestione dei boschi cedui nell’area mediterranea, dopo la crisi degli anni ’50, si è andata caratterizzando per successivi adattamenti ai mutamenti economici e sociali, sia con adeguamenti del trattamento, sia con la scelta di opzioni colturali diversificate. Queste ultime si sono tradotte nella sospensione delle utilizzazioni, che hanno determinato il cosiddetto “invecchiamento” dei cedui, e nell’avviamento all’alto fusto, che ha interessato sopratutto le zone di proprietà pubblica ([1]).
L’attuale presenza sul territorio di queste realtà colturali non è ancora consistente, ma la tendenza in atto a valorizzare anche le funzioni non produttive del bosco fa presumere un loro incremento nel prossimo futuro, unitamente ad una revisione complessiva del trattamento selvicolturale delle formazioni di origine agamica. Numerosi studi sono stati focalizzati sugli aspetti tecnici di questa opzione selvicolturale, con importanti applicazioni pratiche sulla gestione forestale pubblica. Per contro, le conoscenze scientifiche dell’impatto dei trattamenti di conversione sulla funzionalità e l’efficienza d’uso delle risorse di questi ecosistemi (efficienza d’uso dell’acqua, efficienza fotosintetica ed efficienza d’uso dell’azoto) sono limitate. Quando, invece, l’opportunità di applicare opzioni gestionali diversificate impone la definizione di tecniche di trattamento fondate sulla valutazione di parametri non solo produttivi ma anche di ordine funzionale.
Oggigiorno sono disponibili nuovi strumenti di indagine per studiare la risposta degli alberi e dei popolamenti forestali alla variazione dei fattori ambientali. Tra questi, di particolare rilevanza è l’uso degli isotopi stabili del carbonio e dell’idrogeno, per la loro capacità di integrare nel tempo le risposte a livello di processi fisiologici e per l’idoneità ad essere utilizzati anche in schemi di indagine che prevedano campionamenti intensivi.
I valori di discriminazione dell’isotopo stabile del carbonio 13 rispetto al carbonio 12, più abbondante in natura, e del deuterio rispetto all’idrogeno, possono fornire importanti indicazioni, rispettivamente sul rapporto tra assimilazione fotosintetica (A) e traspirazione (E) (efficienza d’uso dell’acqua, WUE=A/E) e sulle strategie di approvvigionamento idrico (falde profonde, falde superficiali, nebbie), come ampiamente dimostrato in letteratura ([20], [32], [13]). Più in dettaglio, la discriminazione del carbonio 13 è direttamente dipendente dal rapporto tra le concentrazioni della CO2 presente negli spazi intercellulari della foglia e di quella presente nell’atmosfera (Ci /Ca), riflettendo in questo modo il bilancio tra il tasso di consumo interno di CO2 dovuto all’attività fotosintetica e l’ingresso di CO2 mediato dal grado di apertura stomatica. È possibile, quindi, mettere in relazione la WUE con la composizione isotopica del carbonio (δ13C - [19]). Prodotti fotosintetici a più rapido turn-over (zuccheri solubili) riflettono nel breve periodo le fluttuazioni della disponibilità idrica e dell’intercettazione luminosa, mentre il δ13C delle foglie e del legno riflette solo parzialmente il segnale delle condizioni ambientali del momento ([10], [22]). A livello fogliare il segnale del δ13C è riferito in parte ai carboidrati prodotti e immagazzinati negli organi di riserva della pianta durante il precedente anno e successivamente rimobilitati, e in parte a quelli prodotti durante l’anno corrente. Pertanto, il δ13C misurato a livello fogliare rappresenta un’integrazione temporale di lungo termine di WUE della pianta, poiché parte del carbonio strutturale è stato verosimilmente assimilato da foglie più vecchie e poi successivamente ritraslocato alle foglie più giovani. È quindi necessario tenere conto della traslocazione del carbonio verso gli apici meristematici e gli organi di accumulo ([6]).
L’efficienza d’uso dell’acqua, definita come rapporto delle moli di CO2 fissata (A) rispetto alle moli di H2O traspirata (E) è inversamente correlata con la disponibilità idrica del suolo ([16], [17], [25]), poiché l’incremento di E è maggiore di quello di A in condizioni di migliore disponibilità idrica ([26]). Nelle foreste sottoposte a trattamenti selvicolturali questa risposta può essere mediata o anche ’sopraffatta’ da un simultaneo incremento dei nutrienti o della disponibilità di radiazione luminosa associato alla perturbazione ([33]). Per contro, in termini assoluti il rapporto A/E potrebbe rimanere invariato in seguito al diradamento ([23], [27]), sebbene i due parametri possano variare in funzione dell’incremento della disponibilità di luce o di azoto, che può determinare un aumento di A, e per effetto dell’incremento di disponibilità idrica del suolo, che può determinare un aumento di E.
Analogamente, gli isotopi stabili dell’idrogeno nell’acqua possono essere impiegati per tracciare le risorse idriche utilizzate dalle piante e per completare le informazioni sui modelli spaziali di sviluppo radicale ([11], [34], [30], [21], [31], [14], [12], [15]), poiché durante l’assorbimento di acqua dalle piante non intervengono fenomeni di frazionamento isotopico dell’idrogeno. Durante il trasporto dell’acqua all’interno della pianta, la composizione isotopica dell’idrogeno (δD) mantiene lo stesso valore di quella dell’acqua nel suolo, fino alla superficie fogliare, dove si innescano i processi evapotraspirativi. A livello fogliare l’acqua si presenta arricchita in deuterio (D). Di conseguenza conoscendo il valore del δD dell’acqua xilematica, delle precipitazioni, delle sorgenti e delle falde è possibile risalire al tipo di risorsa idrica che la pianta sta utilizzando.
Obiettivo di questo lavoro è stato quello quello di verificare l’impatto di un intervento di conversione all’alto fusto, condotto in due cedui quercini mediterranei, su alcuni importanti processi fisiologici, determinanti la capacità di acclimatazione degli alberi ai fattori ambientali e il loro accrescimento. Come indicatori dei processi di acclimatazione sono stati valutati: i) morfologia fogliare e contenuto di clorofille e azoto, per le variazioni indotte nel regime radiativo e nel profilo termico; ii) isotopi stabili del carbonio e potenziale idrico, per le variazioni di WUE e delle relazioni idriche albero-suolo in seguito alla modificazione del bilancio idrologico; iii) isotopi stabili dell’idrogeno, per eventuali variazioni della capacità di approvvigionamento idrico degli alberi in seguito alla riduzione della competizione e alla riduzione dell’intercettazione delle precipitazioni.
Materiali e metodi
Siti sperimentali e trattamenti selvicolturali
Foresta di Pula (CA)
L’area di studio è localizzata a Is Cannoneris, Pula-CA (39°03’ N, 8°50’ E), alla quota di 900 m s.l.m. Il clima è sub-umido ([2]), con moderata deficienza idrica estiva (35 mm). La temperatura media annua è di 15 °C e le precipitazioni medie annue sono di 1000 mm. Il regime pluviometrico è influenzato dalla vicinanza del mare ed è caratterizzato da un massimo autunnale e uno primaverile.
L’area studiata è un ceduo invecchiato di 53 anni, dominato da Quercus ilex L. e Arbutus unedo L., con sporadica presenza nel piano inferiore di Phillyrea latifolia L. e Juniperus communis L. Nove aree sperimentali quadrate, ciascuna di 40 x 40 m sono state istituite e rese permanenti, secondo uno schema a blocchi randomizzati, nel periodo del primo taglio preparatorio di avviamento all’alto fusto, a cura dell’Istituto Sperimentale per la Selvicoltura di Arezzo (CRA-ISSA).
Nel mese di marzo 1994 sono stati realizzati interventi selvicolturali su soprassuoli ceduati 45 anni prima. Il diradamento per l’avviamento all’alto fusto, di tipo misto (dal basso e selettivo), ha asportato dal 50% (grado A) al 60% (grado B) dell’area basimetrica, privilegiando il leccio rispetto alle altre specie componenti e costituendo soprassuoli tendenzialmente monoplani per l’eliminazione del piano sottoposto ([2]).
I due trattamenti selvicolturali oggetto del presente studio sono stati:
- controllo, evoluzione naturale del ceduo senza applicazione di alcun trattamento selvicolturale (C);
- conversione ad alto fusto attraverso un primo taglio di avviamento che ha rimosso circa l’80% dei polloni presenti e il 60% dell’area basimetrica (grado B).
Al momento dell’inizio del presente lavoro, l’ultimo intervento selvicolturale risaliva a più di 50 anni prima nelle parcelle controllo e a circa 8 anni nelle parcelle avviate all’alto fusto.
Foresta di Poggio Pievano (GR)
L’area studiata è un ceduo invecchiato di 43 anni localizzato a Poggio Pievano-GR (43°09’ N, 10°54’ E), a una quota di 570 m s.l.m. Il clima è umido (Amorini et al 1996a), con moderata deficienza idrica estiva. La temperatura media annua è di 14.5 °C, il regime pluviometrico è tipicamente Mediterraneo (942 mm per anno con 105 giorni piovosi).
All’interno del popolamento Quercus cerris L. rappresenta circa l’80% del numero totale dei polloni. Nel piano dominato è presente Fraxinus ornus L. e Acer campestre L. e alcune specie del genere Quercus (Q. ilex L. e Q. pubescens Willd.).
Ampie parcelle sperimentali (di dimensione pari a circa 0.5 ha) sono state permanentemente istituite all’interno della foresta, durante il periodo del primo intervento di conversione ad alto fusto, dall’ISSA ([3]).
Una delle parcelle sperimentali è stata selezionata come controllo (C), mentre l’altra è stata sottoposta a un primo taglio di conversione all’alto fusto (febbraio 1995) all’età di 35 anni, con rimozione di circa l’80% dei polloni e circa il 50% dell’area basimetrica (grado A).
Pertanto, all’inizio del presente lavoro l’ultimo intervento selvicolturale risaliva a più di 40 anni nelle parcelle controllo e a circa 7 anni nelle parcelle diradate.
Rilievi e analisi
Campionamento
Il campionamento è stato effettuato durante i mesi di giugno (foresta di Pula) e luglio (foresta di Poggio Pievano) del 2002. Nel sito sperimentale di Pula sono state campionate tre specie caratteristiche delle foreste sclerofille sempreverdi quali il leccio, la fillirea e il corbezzolo mentre nel sito sperimentale di Poggio Pievano è stato campionato solamente il cerro. In tutte le parcelle di entrambi i siti sperimentali, da alberi appartenenti al piano dominante, sono stati prelevati campioni fogliari dalla parte superiore della chioma (foglie di luce) mentre per il solo sito di Poggio Pievano sono state prelevate anche le foglie d’ombra dei polloni del piano dominato per la determinazione del rapporto tra clorofille a e b, per l’analisi del rapporto isotopico13 C/12C e del contenuto di azoto e carbonio. Gli alberi campione, in numero di 6-10 per trattamento e per specie, sono stati scelti tra quelli dominanti e codominanti e, nelle parcelle controllo di Poggio Pievano, anche nel piano dominato. Inoltre, per la determinazione del rapporto isotopico deuterio/idrogeno (D/H), sono stati prelevati campioni xilematici (per entrambi i siti), e di acqua di falda (solo per il sito di Pula). Nel sito sperimentale di Poggio Pievano non è stato possibile campionare l’acqua di falda, tuttavia alcuni campioni di acqua di sorgente prelevati in prossimità delle parcelle sperimentali sono tuttora in fase di analisi. Pertanto lo studio dell’analisi isotopica dell’idrogeno è stato esteso al continuum suolo-albero-atmosfera solo per il sito sperimentale di Pula.
Contenuto di azoto, di carbonio e δ13C
I campioni fogliari, sigillati in buste di plastica ed essiccati in stufa ventilata a una temperatura di 70 °C per 48 ore, sono stati finemente macinati in azoto liquido. Successivamente, i campioni sono stati analizzati con uno spettrometro di massa (Finnigan delta S, ThermoQuest, Brema, Germania) per determinare la composizione isotopica del carbonio (δ13C) e quindi il rapporto di discriminazione isotopica: Δ = (δ13Caria - δ13Cfoglia)/(1+δ13Cfoglia), che può essere interpretato come proxy dell’efficienza di uso dell’acqua ([18]); il δ13C dell’aria è stato assunto pari a -8‰ e costante fra i diversi trattamenti ([7]).
Concentrazione di clorofille
I campioni fogliari, conservati in azoto liquido, al buio a 80 °C, sono stati sottoposti a estrazione delle clorofilla a (Chl a) e b (Chl b) e dei carotenoidi, mediante una soluzione di Etanolo al 95%. La lettura di assorbanza è stata effettuata con uno spettrofotometro Lambda 3B (Perkin Elmer, Norwalk, USA) alle lunghezze d’onda di 664.2 nm e di 648.6 nm. Il calcolo delle concentrazioni delle clorofille è stato realizzato in accordo con quanto descritto da Licthenthaler ([24]).
Morfologia fogliare
Sui campioni fogliari sono state misurate le seguenti caratteristiche morfometriche: superficie fogliare mediante misuratore di area fogliare a diodi (LI-3100, Li-Cor Inc, Lincoln, NE), peso secco fogliare e rapporto superficie fogliare-peso secco (SLA). Per la determinazione del peso secco i campioni sono stati posti in stufa ventilata, a una temperatura di 70 °C, fino a peso costante.
Rapporto isotopico D/H
Per il calcolo della composizione isotopica dell’idrogeno δD lungo il continuum suolo-albero-atmosfera sono stati analizzati dei campioni di acqua provenienti rispettivamente dalla falda, dalle precipitazioni e da tessuti xilematici di rami. Al fine di evitare eventuali perdite per dispersione ed evaporazione dell’acqua, i campioni, posti in barattoli a tenuta stagna, sono stati conservati ad una temperatura di 4 °C fino al momento dell’estrazione. La linfa grezza presente nei tessuti xilematici è stata criogenicamente estratta sotto vuoto, applicando il procedimento descritto da Dawson & Ehleringer ([11]). Successivamente, l’idrogeno è stato ridotto nella forma diatomica a una temperatura di 500 °C, utilizzando lo zinco come catalizzatore ([8]). I campioni così ottenuti sono stati analizzati in uno spettrometro di massa e δD è stato espresso rispetto allo standard internazionale Vienna Mean Ocean Water (V-SMOW). Il valore medio di δD delle precipitazioni è stato calcolato, per il periodo 1973-1991, utilizzando i dati della stazione meteorologica IAEA (International Agency Energy Atomic) di Genova. Questo valore è stato usato come riferimento per le precipitazioni del sito sperimentale di Pula (CA).
Potenziale idrico
Per ogni specie, il potenziale idrico di base (Ψb) è stato misurato con una camera a pressione ([28]) su tre (Pula) e cinque (Poggio Pievano) foglie raccolte prima del sorgere del sole da alberi differenti nelle due tesi sperimentali. Tali misure sono state condotte nello stesso periodo di campionamento delle altre variabili ecofisiologiche.
Risultati
Clorofille e carotenoidi
Le quattro specie studiate presentavano contenuti di clorofille totali per unità di superficie fogliare, che variavano fra i 58.3 ± 2.6 µg cm-2 del cerro e gli 82.4 ± 3.0 µg cm-2 della fillirea. Nelle tre specie di Pula il rapporto Chla/Chlb aveva un valore medio di 1 e raggiungeva valori superiori a 3 nel cerro.
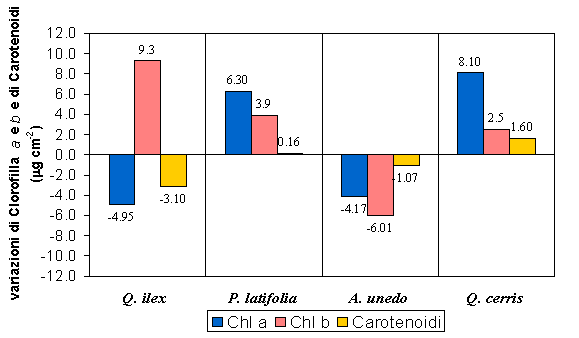
La concentrazione di clorofilla totale appariva generalmente più elevata nelle tesi trattate (Fig. 1), suggerendo un aumento della capacità fotosintetica venutosi a determinare dopo il diradamento (Q. ilex P < 0.05, P. latifolia P < 0.05 e Q. cerris P < 0.05). A. unedo era l’unica specie a mostrare una riduzione significativa di tale parametro (P < 0.01), evidenziando dopo l’intervento una coordinata riduzione delle concentrazioni di clorofilla e di carotenoidi (Fig. 2). Q. cerris e P. latifolia hanno mostrato un incremento delle tre variabili e in particolare della concentrazione di clorofilla a. In un contesto di variazioni complementari tra i pigmenti fotosintetici, Q. ilex è stata la sola specie a mostrare delle differenze contrastanti: ad un sensibile aumento delle clorofille b si è contrapposto un decremento delle clorofille a e dei carotenoidi.
Fig. 1 - Confronto tra il contenuto di clorofilla totale nelle due tesi sperimentali per tutte le specie studiate. Le barre indicano l’errore standard della media (Pula n=6 per specie, Poggio Pievano n=10).
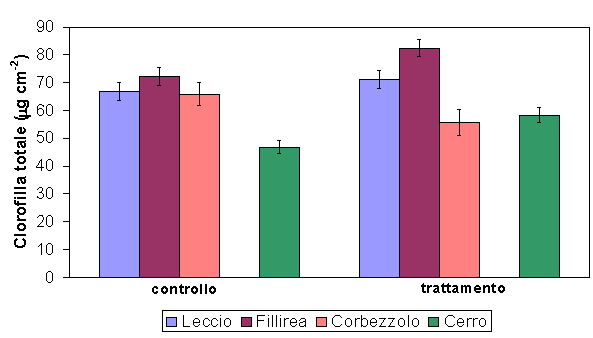
Fig. 2 - Differenze tra i contenuti dei principali pigmenti fotosintetici osservate dopo 7-8 anni dal diradamento.
In entrambi le tesi sperimentali il contenuto di clorofilla a risultava correlato con la concentrazione di azoto fogliare (R2 = 0.71 nelle parcelle di controllo e R2 = 0.87 nelle tesi di trattamento), come mostrato in Fig. 3: si osservi che il leccio e il corbezzolo rappresentano le specie con i più bassi contenuti di clorofilla a e di azoto, con valori non significativamente più bassi dopo il diradamento, mentre il cerro raggiungeva valori più alti e presentava un incremento significativo (P < 0.05) dopo l’esecuzione del diradamento.
Fig. 3 - Relazione tra il contenuto di clorofilla a e azoto fogliare. I simboli aperti indicano le parcelle diradate, i simboli chiusi indicano le parcelle di controllo; le barre indicano l’errore standard della media (Pula: n =6 per specie, Poggio Pievano: n=10).
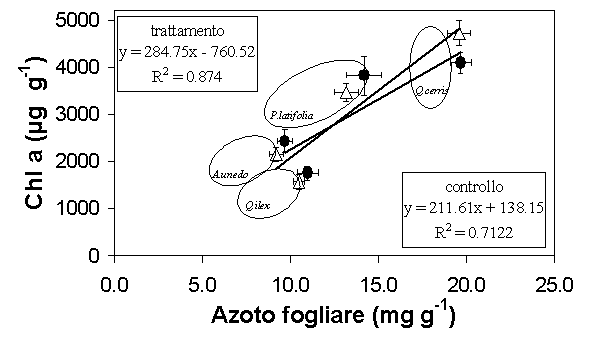
In questo modo dopo 7-8 anni dall’esecuzione del diradamento si è verificato un aumento della pendenza della retta di regressione (P < 0.001) in coincidenza di un incremento lineare della concentrazione di clorofilla a e del contenuto di azoto.
Morfologia fogliare
Il cerro presentava un’area fogliare di dimensioni mediamente maggiori rispetto a quella delle tre specie principali costituenti la lecceta di Pula. Tuttavia, i valori osservati nel cerro erano sensibilmente ridotti nella tesi di controllo (20.9 ± 0.9 cm2), con i valori minimi registrati nelle piante del piano dominato (17.7 ± 0.9 cm2). Valori significativamente più alti (P < 0.05) sono stati osservati nelle parcelle sottoposte al trattamento selvicolturale (22.5 ± 0.5 cm2). Una risposta simile, seppure in misura non significativa, è stata osservata anche nelle tre specie studiate a Pula.
Il rapporto superficie-peso fogliare (area fogliare specifica, SLA), risultava mediamente più basso nelle tre specie sempreverdi di Pula, rispetto al cerro; tale osservazione è coerente con la diversa anatomia delle specie studiate. Per questo parametro non era verificabile un effetto univoco del trattamento, probabilmente a causa dell’interazione di più fattori di controllo. La fillirea è stata l’unica specie a mostrare una diminuzione significativa del parametro SLA (P < 0.01) dopo 8 anni dal diradamento, probabilmente in relazione al maggior grado di plasticità fogliare caratteristico di questa specie. A Poggio Pievano sono state osservate differenze significative (P < 0.001) del parametro SLA tra piano dominante (111.5 ± 8.6 cm2 gr-1) e dominato (143.5 ± 5.2 cm2 gr-1) e tra il piano dominato del controllo e trattamento (P < 0.001), come conseguenza del diverso regime luminoso.
Contenuto di carbonio e azoto
Il contenuto di azoto fogliare, espresso per unità di peso fogliare, era mediamente più elevato nel cerro di Poggio Pievano (20 ± 0.1 mg g-1), rispetto alle specie di Pula (11 ± 1 mg g-1). Fra queste ultime i valori più elevati sono stati registrati per la fillirea (14 ± 0.9 mg g-1). Il contenuto di azoto fogliare degli alberi campionati nelle parcelle di controllo era sempre leggermente superiore, anche se non in modo significativo, a quello misurato nelle parcelle diradate, per tutte le specie studiate. Il contenuto di carbonio non variava sensibilmente nelle diverse specie e in funzione del trattamento. Significativamente differenziati risultavano invece i valori del rapporto C/N nelle diverse specie esaminate: la specie con più basso valore C/N fogliare risultava il cerro (23.7%), mentre quella con i valori più elevati era il corbezzolo (50.3%). Il rapporto C/N era sempre leggermente inferiore nelle parcelle controllo, rispetto a quelle trattate.
Potenziale idrico di base
I valori di potenziale idrico prima dell’alba non risultavano mai inferiori a 1.5 MPa, a testimonianza di un livello di stress idrico relativamente moderato. I potenziali risultavano meno negativi nelle piante del piano dominato e/o delle parcelle controllo, anche se non è stato evidenziato un effetto significativo del trattamento.
Composizione isotopica del carbonio 13 e composizione isotopica dell’idrogeno
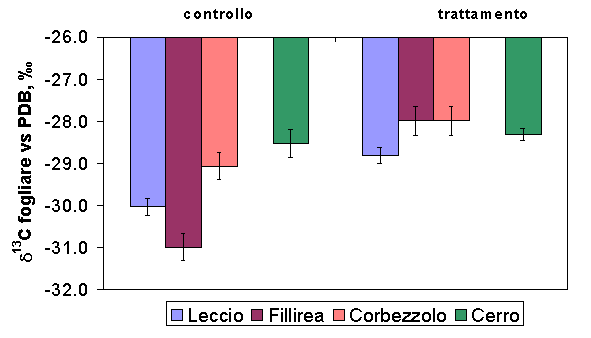
Per quanto riguarda le abbondanze isotopiche del carbonio, i valori di δ13C osservati nelle differenti specie e per entrambi i siti rientrano nella parte bassa dell’intervallo riportato per le specie ad assimilazione fotosintetica C3. Assumendo che il δ13C dell’aria non vari all’interno di uno stesso sito sperimentale, è possibile imputare le differenze osservate a una diversa discriminazione del carbonio nel corso dell’attività fotosintetica. I valori ottenuti per le specie della foresta di Pula e di Poggio Pievano si attestano intorno ad un valore medio di 29‰. La specie quercina dominante della foresta sempreverde (Q. ilex) è caratterizzata da valori più bassi di δ13C (-30.03 ± 0.2‰), in confronto con la specie decidua Q. cerris (-28.53 ± 0.33‰). Per entrambi i siti sperimentali, le specie del piano dominato, come la fillirea (Pula) e i polloni dominati di cerro (Poggio Pievano) mostravano valori di δ13C più negativi a confronto delle specie del piano dominante (-31 ± 0.32‰ per la fillirea e 30 ± 0.13‰ per il cerro). Dopo il diradamento, il δ13C di tutte le specie e per entrambi i siti risultava aumentato, di 1.21‰ per Q. ilex (P < 0.05), 3‰ per P. latifolia (P < 0.01), 1.08‰ per A.unedo (P < 0.01), e 0.22‰ per Q. cerris (n.s. - Fig. 4). Questi risultati possono essere interpretati come segno di un aumento della WUE, possibile effetto della riduzione della densità del popolamento, che comporta a sua volta un aumento della disponibilità di luce e di acqua per le piante.
Fig. 4 - Confronto tra la composizione isotopica del carbonio (δ13C) fogliare nelle due tesi sperimentali per tutte le specie campionate. Le barre indicano l’errore standard della media (Pula: n=6 per specie, Poggio Pievano: n=10).
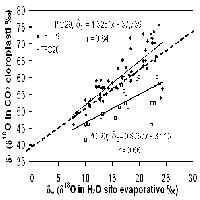
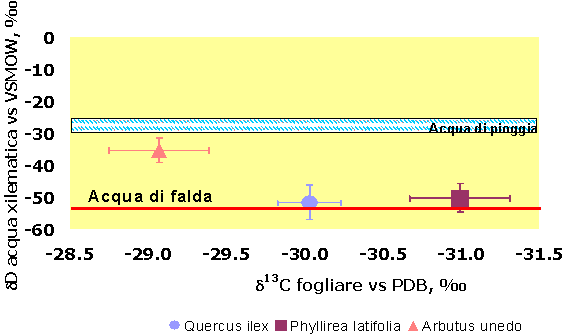
Per quanto riguarda le abbondanze isotopiche dell’idrogeno, i valori di δD misurati sulle quattro specie e in entrambi i siti, evidenziano due gruppi di piante con differente modalità di accesso alla risorsa idrica (Fig. 5). Le specie con apparato radicale profondo come Q. ilex, P. latifolia, e Q. cerris presentano bassi valori di δD, mentre A. unedo, caratterizzato da apparato radicale superficiale, presenta valori sempre significativamente più alti. Dopo il diradamento, i valori di δD sono aumentati in tutte le specie: i cambiamenti sono stati di circa il 5’ per le specie sempreverdi e di più del 10’ per la specie decidua (Q. cerris). Dopo 7-8 anni dall’esecuzione del diradamento la linfa grezza risultava arricchita in deuterio, con δD compreso nell’intervallo di assorbimento tra acqua di falda e acqua di pioggia (miscela falda-pioggia). Valori molto negativi di δD sono generalmente osservati in specie con apparato radicale profondo, capaci di accedere al livello della falda acquifera, come confermato dal segnale misurato nelle parcelle di controllo del sito di Pula (Fig. 6 - [31]).
Fig. 5 - Confronto tra la composizione isotopica del deuterio (δΔ) della linfa grezza nelle due tesi sperimentali. Le barre indicano l’errore standard della media (Pula: n=6 per specie, Poggio Pievano: n=10).
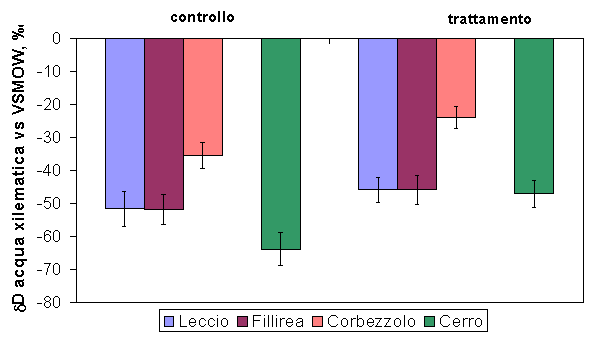
Fig. 6 - Comparazione del segnale del δ13C fogliare e del δΔ dell’acqua di pioggia, di falda e xilematica, all’interno delle parcelle di controllo. I livelli di acqua di pioggia e acqua di falda sono indicati dalle linee tratteggiate. Le barre indicano l’errore standard della media (n=6 per specie).
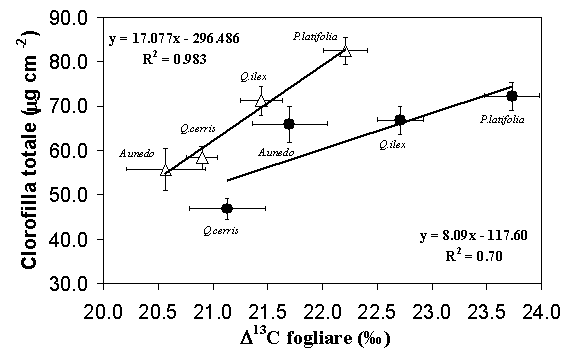
In Fig. 7 sono mostrate le relazioni trovate fra il valore di discriminazione fogliare del carbonio 13 (Δ13C) e il contenuto di clorofilla totale, per i due trattamenti sperimentali. Nelle parcelle di controllo la relazione mostra un coefficiente di regressione pari a 0.70 con un intervallo di Δ13C più ampio, rispetto a quello delle parcelle diradate. Come già visto il contenuto di clorofilla era mediamente più basso in queste parcelle. Diversamente, nelle parcelle diradate si osservava una più stretta dipendenza fra le due variabili (R2 = 0.983) e un incremento della pendenza della retta di regressione.
Fig. 7 - Valori medi di clorofilla totale (Chltot) verso discriminazione isotopica del carbonio 13 fogliare (δ13C). Le parcelle diradate sono indicate con i simboli aperti (y = 17.077 x -296.48) e le parcelle di controllo (y = 8.09 x 117.6) con i simboli chiusi. Le barre indicano l’errore standard della media (Pula: n=6 per specie, Poggio Pievano: n=10).
Discussione
Un elevato valore del rapporto Chla/Chlb è frequentemente associato a condizioni di maggiore illuminazione ([4]), in accordo con quanto verificato nella cerreta. Tale diversa condizione era evidenziata anche a livello delle concentrazioni fogliari dei principali pigmenti fotosintetici (clorofilla a, clorofilla b e carotenoidi) che risultavano sensibilmente superiori nella cerreta rispetto a quelli della lecceta mediterranea.
Per quanto riguarda gli effetti dello stato idrico, la debole correlazione tra δ13C e potenziale idrico di base (Ψb - R2 = 0.109) suggerisce che circa il 90% delle variazioni di δ13C sono attribuibili a variabili diverse dallo stato idrico istantaneo.
Nell’insieme, effetti positivi del diradamento sullo stato idrico sono stati evidenziati nel corso dei primi tre-cinque anni successivi all’intervento; in seguito si è osservato un progressivo peggioramento dello stato idrico delle piante per la progressiva chiusura della copertura arborea ([9]). A livello epigeo il ripristino della copertura arborea potrebbe aver provocato un maggiore incremento del tasso di assimilazione fotosintetico (A) rispetto a un parallelo incremento della traspirazione (E). Allo stesso tempo, un incremento della traspirazione complessiva del popolamento a seguito del diradamento potrebbe aver determinato una riduzione della disponibilità idrica del suolo, che potrebbe quindi spiegare le variazioni osservate di δ13C e di δD.
Nel corso di precedenti studi su foreste temperate, Bréda et al. ([5]) e Geβler et al. ([22]) hanno messo in evidenza che i trattamenti selvicolturali causano un immediato decremento dell’indice di area fogliare e dell’intercettazione delle precipitazioni, incrementando in questo modo la disponibilità idrica nelle piante rilasciate. Tali osservazioni si ripercuotono con una produzione di tessuti fogliari impoveriti in δ13C.
Negli ambienti mediterranei sembrerebbe, invece, che l’effetto di maggiore importanza del diradamento sia quello sul regime radiativo, che determina un incremento del tasso di assimilazione fotosintetico con contestuale incremento dell’efficienza di uso idrico. I risultati ottenuti con il presente studio paiono infatti indicare che dopo 7-8 anni dall’esecuzione del diradamento si è instaurato un nuovo equilibrio fra le quantità di azoto fogliare e di luce trasmessa dalle chiome e del contenuto di umidità del suolo, spostato a favore di una maggiore WUE. Analoghe conclusioni sono state riportate da Sparks & Ehleringer ([29]), che hanno rilevato una robusta correlazione tra δ13C e contenuto di azoto fogliare, dimostrando come ad elevate concentrazioni di azoto fogliare corrisponda un incremento della capacità fotosintetica e una riduzione delle concentrazioni intercellulari della CO2 fogliare.
I diversi valori di δD delle tre specie sempreverdi di Pula, possono essere associati alle variazioni dei valori di δ13C. Indipendentemente dal potenziale idrico di base, le specie che utilizzavano principalmente acqua di falda (Q. ilex e P. latifolia) presentavano valori molto negativi di δ13C. Diversamente, A. unedo che utilizzava maggiormente l’acqua di precipitazione, presentava valori meno negativi e quindi un’elevata WUE. La differente efficienza d’uso dell’acqua delle tre specie potrebbe avere quindi una base adattativa, legata alla diversa morfologia radicale. La risposta al diradamento, con incremento dei valori di δD e contestuale riduzione dei valori di δ13C, dimostra l’instaurarsi di un nuovo equilibrio fra disponibilità idriche e efficienza nel loro uso e rappresenterebbe una risposta di acclimatazione delle piante alle nuove condizioni ambientali.
In conclusione, lo studio ha dimostrato l’efficacia delle metodologie di discriminazione isotopica nella definizione della risposta funzionale a interventi selvicolturali di conversione ad alto fusto; ha inoltre evidenziato che, in sistemi forestali complessi come quelli mediterranei, gli effetti dei trattamenti si protraggono per più anni, suggerendo anche che una ripetizione cadenzata dei diradamenti possa contribuire alla realizzazione di strutture più efficienti nell’uso delle risorse ambientali.
Ringraziamenti
Lavoro svolto nell’ambito del programma di ricerca del Ministero per le Politiche Agricole e Forestali RI.SELV.ITALIA - Sottoprogetto 3.2.1: “Efficienza funzionale e interventi selvicolturali in boschi cedui mediterranei”, coordinato dal Dr. Gianfranco Fabbio. Gli autori ringraziano l’Azienda Foreste Demaniali della Regione Sardegna per aver consentito l’accesso alle aree di ricerca e per aver contribuito con uomini e mezzi all’attività in campo. In particolare il dott. Tiziano Mei, il tecnico forestale Venanzio Basciu e il personale del Cantiere Forestale “Is Cannoneris”. Per le aree di ricerca in Toscana, gli autori ringraziano la Comunità Montana “Colline Metallifere” e l’Istituto Mareografico e Oceanografico di Pisa per avere gentilmente fornito i dati termopluviometrici della stazione di Massa Marittima. Un ringraziamento particolare ai ricercatori dell’Istituto Sperimentale per la Selvicoltura di Arezzo: Gianfranco Fabbio, Emilio Amorini, Andrea Cutini, Maria Chiara Manetti. Si ringrazia inoltre la Sig.ra Matilde Tamantini (tecnico presso il DISAFRI) per il sostanziale contributo apportato durante la fase di analisi di laboratorio e il Sig. Ermenegildo Magnani (tecnico presso il laboratorio Isotopi Stabili del Consorzio Agrital-Ricerche di Maccarese) per il sostanziale contributo durante la fase delle analisi isotopiche.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar