Impact of traffic on δ15N, δ13C and δ18O of needles and annual tree rings of Norway spruce (Picea abies L.)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 3, Pages 437-445 (2006)
doi: https://doi.org/10.3832/efor0389-0030437
Published: Sep 20, 2006 - Copyright © 2006 SISEF
Research Articles
Guest Editors: 5° SISEF Congress (Grugliasco, TO - 2005)
« Forests and Society - Changes, Conflicts, Sinergies »
Collection/Special Issue: E. Lingua, R. Marzano, G. Minotta, R. Motta, A. Nosenzo, G. Bovio
Abstract
Increase of nitrogen depositions, as oxidized (NOx) and reduced (NHx) compounds, has important implications on ecosystem nitrogen cycle. NOx comes predominantly from fossil fuel combustion in car engines and industrial processes, while agricultural activities (cattle farming, fertilizations) are the main sources for NHx emissions. Such fertilisations could stimulate growth and, therefore, productivity of forests, especially in temperate forests, where nitrogen is a limiting factor. On the other hand an excess of nitrogen can lead to an acidification of the soil and have a negative impact on the microbial fauna and structure of plants. NOx and NHx depositions can be separated with the help of stable isotopes with different 15 N-values reported for NOx originating from combustion sources. Consequently it was observed that the nitrogen isotopic composition of the vegetation reflects the isotopic signature of nitrogen sources. Our preliminary results on needles of Norway spruce trees exposed to NOx emissions along a transect perpendicular to a highway close to Faido and Brittnau show a clear isotopic enrichment in 15 N in trees growing closer to traffic pollution, indicating an uptake of atmospheric nitrogen by stomatal pathway. Carbon and oxygen isotopic composition measured in tree rings reveals the physiological response in plants. Trees more exposed to air pollution from traffic show an increase in δ13 C and δ18 O in tree rings, since mid sixties next to Erstfeld. This could mean a higher photosynthetic activity, enhanced by NOx traffic emissions, under low or not changed stomatal conductance. Our results confirm that stable isotopes of carbon, oxygen and nitrogen measured in needles and tree rings represent an important tool to monitor the impact of NOx load on tree physiology.
Keywords
Spruce, Traffic, Nitrogen, Deposition, Growth rings, Needles, Stable isotopes
Introduzione
L’aumento delle deposizioni di azoto osservato negli ultimi decenni ha importanti implicazioni sul ciclo globale dell’azoto e sui processi biogeochimici che avvengono negli ecosistemi forestali. Pitcairn et al. ([14]) riportano un aumento delle deposizioni di azoto in Europa da 2-6 a 15-60 kg ha-1 anno-1, relativamente ai due decenni precedenti. L’azoto è presente nell’atmosfera in forma ossidata (NOx) e ridotta (NHx) ed il suo apporto negli ecosistemi terrestri ed acquatici può avvenire mediante deposizioni umide o secche ([5]). Le emissioni di NOx sono causate principalmente dalla circolazione di autoveicoli e da attività industriali, che, secondo quanto riportato da Bragazza et al. ([5]), costituirebbero più del 70% delle emissioni totali di NOx. Quelle di NHx derivano principalmente da attività agricole di natura intensiva (allevamenti animali e fertilizzazioni di monoculture specializzate). Nonostante recenti progressi tecnologici abbiano consentito una notevole riduzione delle emissioni, soprattutto grazie all’introduzione della benzina senza piombo e della marmitta catalitica ([13]), le emissioni di NOx costituiscono ancora un serio problema per l’impatto che continuano ad avere sull’ambiente, provocando alte concentrazioni di ozono nella troposfera e processi di acidificazione e di eutrofizzazione degli ecosistemi terrestri ed acquatici ([10]).
Negli ecosistemi forestali, queste deposizioni hanno importanti implicazioni legate al ciclo globale dell’azoto. Nelle foreste temperate, in particolare, dove spesso l’azoto rappresenta il fattore limitante, le addizioni di azoto hanno un effetto fertilizzante, perlomeno in una fase iniziale. Deposizioni croniche, tuttavia, provocando un eccesso di azoto rispetto alla domanda da parte delle piante e della microflora tellurica, comportano squilibri nei cicli biogeochimici, con possibili alterazioni dei processi fisiologici delle piante. La maggiore disponibilità di azoto per gli alberi, ad esempio, provoca una riduzione del rapporto della biomassa radici/chioma, alterando l’equilibrio sia strutturale che fisiologico della pianta (in quanto una minore biomassa radicale in proporzione a quella fogliare comporta una scarsa compensazione delle perdite traspirative di acqua). Un altro effetto è stato evidenziato da Bolsinger & Flückiger ([3]) che, accanto ad un’autostrada, hanno osservato un cambiamento della composizione di aminoacidi all’interno delle foglie di Viburnum opulus L. e Phaseolus vulgaris L. che favorirebbe lo sviluppo e la crescita di alcune specie di afidi.
Un aspetto importante da considerare è quello dell’impatto sulle caratteristiche del suolo e sul conseguente cambiamento vegetazionale nel piano sottostante quello arboreo, all’interno di una foresta. In uno studio condotto da Kirchner et al. ([12]) in una foresta di abete rosso nei pressi dell’autostrada A95 in Germania è stata osservata, ad esempio, una diminuzione di Rubus fructicosus L. e Mycelis muralis L. procedendo dal margine verso l’interno della foresta.
L’assorbimento di azoto da parte delle piante avviene generalmente attraverso le radici (sotto forma di ioni NH4+ e di NO3- presenti nel suolo), ma bisogna considerare anche l’assorbimento di NO2 che avviene per via stomatica e che contribuisce in maniera significativa al budget complessivo di azoto ([1], [18]). In questo caso, affinché il gas assorbito possa essere utilizzato per la sintesi degli aminoacidi, viene convertito in nitrato e nitrito, attraverso l’attività della nitrato e nitrito-reduttasi, e in ammonio ([20]).
La variazione della concentrazione dell’isotopo dell’azoto meno abbondante nel materiale vegetale viene espressa come composizione isotopica dell’azoto δ15N ed è data dal rapporto (eqn. 1):
dove come standard si considera l’N2 atmosferico.
Nell’ambito di un ecosistema forestale, il grado di arricchimento o impoverimento di 15N è influenzato dai frazionamenti isotopici durante le trasformazioni biochimiche a livello dei diversi pools (suolo, pianta, atmosfera). In particolare, il δ15N nel materiale vegetale riflette principalmente quello del suolo (derivante da mineralizzazione della sostanza organica, attività microbiche e perdita di azoto) ed è associato ai processi di assimilazione, traslocazione a livello di pianta, nonché alla composizione isotopica dell’azoto nell’atmosfera.
La composizione isotopica dell’azoto misurata nelle foglie e negli anelli legnosi fornisce informazioni sull’input di azoto atmosferico per gli alberi, dal momento che il segnale isotopico delle due forme di deposizioni di azoto, NOx e NHx, è difatti differente: il primo mostra un valore δ15N più alto del secondo ([1], [19]). La combinazione di δ13C, δ18O e δ15N consente di interpretare da un punto di vista fisiologico gli effetti delle deposizioni di azoto sugli alberi. La composizione isotopica del carbonio, infatti, è strettamente legata ai processi biochimici e fisiologici che avvengono durante la fotosintesi (a scala fogliare) e l’accrescimento secondario (traslocazione ed investimento strutturale dei fotosintati nel fusto). Il segnale derivante dal δ18O è espressione, invece, di processi fisico-chimici che hanno luogo a scala fogliare durante la traspirazione. Carbonio e ossigeno consentono di distinguere tra limitazioni biochimiche o del controllo stomatico in risposta a fattori ambientali ([17]).
In studi condotti nei pressi di un’autostrada in Svizzera, sono stati osservati valori di δ15N più positivi in aghi ed anelli legnosi di alberi di abete rosso esposti alle emissioni di NOx derivanti dal traffico, piuttosto che in alberi di controllo ([1], [16]).
L’obiettivo del presente lavoro è quello di i) verificare se vi è un segnale dell’assorbimento di NOx negli aghi di alberi di abete rosso lungo transetti perpendicolari all’autostrada; ii) studiare i possibili effetti dell’aumento del traffico sugli alberi, in seguito alla costruzione dell’autostrada, attraverso uno studio retrospettivo basato su misure dendrocronologiche ed isotopiche (δ13C e δ18O) degli anelli legnosi.
Materiali e metodi
Area di studio e campionamento
Perpendicolari all’autostrada A2, in Svizzera, sono stati individuati tre transetti all’interno di foreste di abete rosso (Picea abies L. Karsten), nei pressi di Brittnau (cantone Argovia), Faido (canton Ticino) ed Erstfeld (cantone Uri - Fig. 1).
Fig. 1 - Area di studio nei pressi di Erstfeld (fonte: ©AlpTransit San Gottardo SA. [online] URL: ⇒ http://www.alptransit.ch/).

Per le analisi isotopiche del δ15N sono stati campionati aghi di due anni da due alberi all’interno di cinque plots a Faido (P1 = 50 m, P2 = 125 m, P3 = 175 m, P4 = 400 m; P5 = 500 m dall’autostrada) e otto plots a Brittnau (P1 = 12.5 m, P2 = 25 m, P3 = 50 m, P4 = 62.5 m; P5 = 75 m, P6 = 87.5 m, P7 = 200 m, P8 = 300 m dall’autostrada). Considerando lo studio sugli aghi preliminare, volto, cioè, ad accertare la presenza o meno di un segnale dell’assorbimento dell’azoto derivante dalle emissioni, si è ritenuto opportuno considerare solo due alberi per ciascun plot.
Tre carote di legno sono state estratte da sei alberi individuati in due aree poste a differente altitudine (approssimativamente 200 m di dislivello) vicino all’autostrada nei pressi di Erstfeld (Fig. 1). In questo caso, tre alberi (indicati con T1, T2 e T3) sono stati scelti nel plot P1 alle pendici di una parete rocciosa, mentre gli altri 3 (indicati con T4, T5 e T6), 200 metri più in alto, nel plot P2. La distanza di ciascun albero dall’autostrada è di: T1 = 99 m, T2 = 127 m, T3 = 193 m, T4 = 474 m, T5 = 495 e T6 = 525 m. L’esposizione della pendice rocciosa è NE, con una pendenza di 30°. Nel plot a quota più elevata il popolamento si presenta più denso, con frequenti consociazioni tra abete rosso e larice (Larix decidua Mill). Nel plot S1, invece, è stata riscontrata la presenza di altre specie quali Abies alba Mill. e Carpinus betulus L.
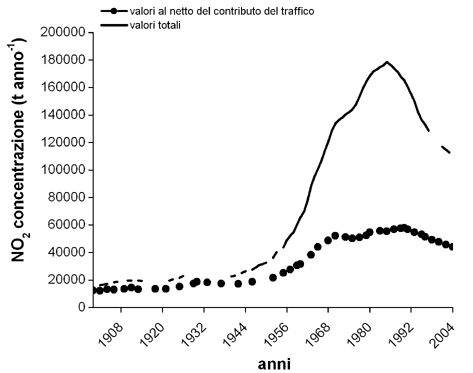
I dati disponibili di emissioni di NO2 (a partire dal 1900) sono relativi all’intera Svizzera e derivano dalla rete nazionale d’osservazione degli inquinanti atmosferici (NABEL), gestita dall’EMPA, su incarico dell’Ufficio Federale dell’Ambiente e delle Foreste ([6]). I valori di NO2 causati dal traffico vengono stimati considerando la differenza tra i valori totali di NO2 derivanti da diverse fonti quali attività industriali, traffico, usi domestici (riscaldamenti) e i valori senza il contributo del traffico. Come è possibile osservare in Fig. 2, si registra un forte aumento della concentrazione di NO2 a partire dagli anni ’60, con un picco intorno agli anni ’80. Da questo punto in poi si osserva una diminuzione delle emissioni, legata probabilmente all’introduzione della marmitta catalitica e della benzina senza piombo (per maggiori informazioni si veda Buwal 1995).
Fig. 2 - Andamento della emissioni di NO2 in Svizzera dal 1900 al 2004. Con la linea continua si rappresentano le emissioni totali di NO2 derivanti da diverse fonti (traffico, attività industriali, riscaldamenti) mentre i simboli indicano l’andamento dei valori di NO2 senza il contributo del traffico. È chiaro il notevole apporto delle emissioni di NO2 derivante dal traffico sulle emissioni totali. Dati riportati dall’Ufficio Federale dell’Ambiente (UFAM).
Preparazione dei campioni
Delle tre carote prelevate da ciascun albero, due sono state utilizzate per le analisi isotopiche ed una per le misure dendrocronologiche. Per essere sicuri dell’esatta datazione di ciascun anello destinato alle analisi isotopiche, abbiamo datato, misurato e crossdatato anche le carote destinate alle analisi isotopiche.
Misure dendrocronologiche
Per ciascuna carota ogni singolo anello è stato identificato, datato e misurato, partendo dalla corteccia fino al midollo, lavorando sotto uno stereomicroscopio. Per le misure delle ampiezze anulari, è stato utilizzato il sistema combinato di tavola misuratrice Lintab e software TSAP (Time Series Analysis and Presentation), entrambi della Frank Rinn, Heidelberg (Germania).
Le curve delle ampiezze anulari ottenute per ogni pianta sono state plottate e, ad una prima fase di confronto visivo (in modo da poter individuare possibili errori), è seguita l’analisi statistica, mediante la quale le curve di ciascuna pianta sono state confrontate con la cronologia media (per maggiori dettagli si veda, ad esempio, [7]). I parametri statistici ottenuti sono indicati in Tab. 1.
Tab. 1 - Parametri derivanti dal confronto statistico (t-student) delle curve delle ampiezze anulari misurate per ciascun albero rispetto alla curva media. L’overlap indica il numero di anni considerati; Gleichläufigkeit rappresenta il coefficiente di concordanza, ovvero la misura della corrispondenza esistente di anno in anno tra andamenti di un determinato intervallo di due cronologie, basata sulla concordanza di segno della variazione (positivo, negativo o nullo - [11]).
| Id campione | Overlap | Gleichläufigkeit | P | t Student |
|---|---|---|---|---|
| T1 | 101 | 74 | 0.001 | 5.3 |
| T2 | 149 | 66 | 0.001 | 9.9 |
| T3 | 116 | 69 | 0.001 | 7.8 |
| T4 | 125 | 63 | 0.05 | 4.3 |
| T5 | 109 | 61 | 0.05 | 3.0 |
| T6 | 96 | 70 | 0.001 | 5.5 |
Analisi isotopiche
Da ciascuna delle due carote destinate alle analisi isotopiche, gruppi di tre anelli sono stati individuati, separati e tagliati. Prima delle misure isotopiche, gli aghi e i gruppi di anelli sono stati macinati con un mulino (Retsch, Germania) e successivamente pesati in capsule di stagno. Il pooling di anelli consecutivi è stato necessario dal momento che spesso il materiale relativo ad un singolo anello non era sufficiente per le misure isotopiche.
La quantità di materiale usata per le analisi è stata, per gli aghi, di 3.8-4.2 mg sia per il carbonio che per l’azoto, mentre per l’ossigeno 1.1-1.3 mg. Per quanto riguarda gli anelli legnosi è stata analizzata una quantità di 0.6-0.8 mg sia per l’ossigeno che per il carbonio. Considerata la bassa concentrazione di azoto nel legno, per la misura del δ15N è stato necessario considerare una quantità di campione pari a 20 mg. I campioni sono stati bruciati in un analizzatore elementare (EA-1100, Carlo Erba, Milano, Italia) per le analisi isotopiche di carbonio e azoto (nel caso del materiale fogliare) e di carbonio (per i campioni legnosi). Dopo la combustione, l’acqua è stata rimossa mediante trappola contenente perclorato di magnesio, mentre N2 e CO2 sono stati separati in una colonna gascromatografica e convogliati successivamente nello spettrometro di massa (Delta - S Finnigan MAT, Bremen, Germania) mediante un’interfaccia (EA-1108, Carlo Erba, Milano, Italia) connessa all’analizzatore elementare.
La composizione isotopica dell’ossigeno è stata misurata separatamente, utilizzando un differente analizzatore elementare, connesso allo stesso spettrometro di massa, che decompone i campioni in assenza di ossigeno, mediante pirolisi termica ad una temperatura di 1080°C ([15]). H2O and CO2 derivanti da questa reazione sono stati rimossi mediante due differenti trappole, perclorato di magnesio e carbosorb, rispettivamente per l’acqua e per l’anidride carbonica, mentre CO e gli altri gas prodotti sono stati separati mediante colonna gascromatografica e convogliati nello spettrometro di massa. Il rapporto 18O/16O è stato calcolato sulle molecole di gas di CO. Per maggiori dettagli metodologici si vedano ad es. [4]; [8]; [9].
Risultati e discussione
Aghi
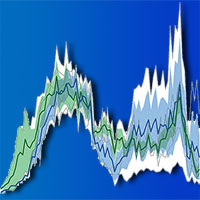
In Fig. 3 è possibile osservare un arricchimento isotopico in 15N negli aghi maggiormente esposti alle emissioni di NOx provenienti dall’autostrada, soprattutto a Brittnau. Questo risultato conferma quanto osservato da Ammann et al. ([1]) su campioni di aghi e di suolo, nei pressi della stessa autostrada, vicino a Solothurn. Nel suddetto lavoro vengono riportati anche i valori di δ15N misurati per NO2 e NO, rispettivamente di + 5.7 ± 2.8 ‰ e di +3.1 ± 5.4 ‰.
Fig. 3 - Andamento del δ15N misurato su aghi campionati dalla porzione alta delle chiome di alberi di abete rosso lungo un transetto perpendicolare all’autostrada nei pressi delle località di Faido e Brittnau.
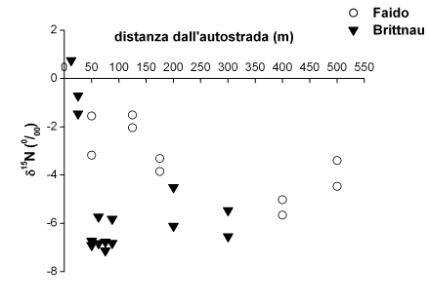
Allontanandosi dall’autostrada il valore di δ15N misurato negli aghi diminuisce. In particolare, a Brittnau, l’impatto delle emissioni provenienti dall’autostrada si riduce drasticamente già a 50 metri di distanza dall’autostrada. In uno studio condotto da Kirchner et al. ([12]), è stata messa in evidenza una diminuzione della concentrazione di NO2 e di NH3 lungo un transetto perpendicolare all’autostrada in una foresta di abete rosso ed è stato osservato un effetto barriera da parte degli alberi immediatamente prossimi all’autostrada, che ridurrebbero notevolmente l’impatto delle emissioni man mano che si procede verso l’interno della foresta. L’arricchimento isotopico osservato soltanto nelle immediate vicinanze dell’autostrada sembra confermare che l’assorbimento di azoto possa avvenire anche per via stomatica ([18]).
Anelli di accrescimento degli alberi
1) Ampiezza anulare
Ciascun punto della curva rappresenta la media dei valori misurati su gruppi di tre anelli consecutivi (in accordo con le misure isotopiche). Si osserva una notevole variabilità dell’ampiezza anulare tra gli alberi in entrambi i plots considerati (Fig. 4), all’interno di una foresta di abete rosso nei pressi di Erstfeld. In generale, si verifica una riduzione dell’accrescimento sino agli anni ’50 (normale trend dovuto all’età) e una successiva e lieve ripresa, soprattutto per le piante 1 e 5. In generale gli alberi non hanno mostrato grosse variazioni delle ampiezze anulari, eccetto per l’albero T1 più vicino all’autostrada, indicando un possibile effetto fertilizzante delle emissioni di NOx provenienti dall’autostrada. Se si considerano i valori medi per ciascun plot (dati non mostrati) si evidenziano valori di accrescimento maggiori negli alberi del plot posto alle pendici della parete rocciosa, più vicino all’autostrada.
Fig. 4 - Andamento delle ampiezze anulari di alberi di abete rosso in due differenti plot (P1 e P2) lungo l’autostrada A2 nei pressi di Erstfeld. Ogni punto rappresenta la media dei valori misurati su 3 anelli consecutivi.
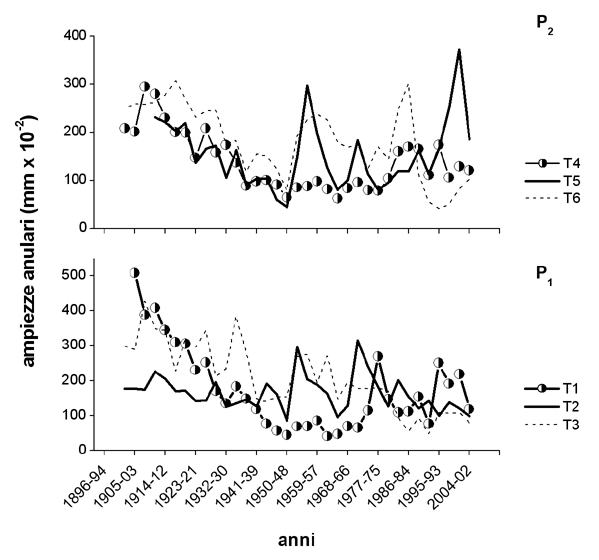
Le basse correlazioni trovate tra ampiezza anulare e dati climatici misurati a Zurigo (Tab. 2) indicano che né le precipitazioni né le temperature sono limitanti per l’accrescimento degli alberi nell’area di studio considerata. Solo per due alberi (T3 e T6) è stata trovata una correlazione significativa (P<0.05) tra ampiezze anulari e temperatura media annua.
Tab. 2 - Coefficiente di correlazione (R) e grado di significatività (P) derivante da correlazioni, per ciascun albero, tra i valori delle ampiezze anulari e temperatura (Temp) e precipitazioni (Prec) medie. I dati climatici sono relativi alla stazione più vicina all’area di studio (Zurigo).
| Id Campione |
Prec | Temp | ||
|---|---|---|---|---|
| R | P | R | P | |
| T1 | 0.01 | 0.95 | -0.20 | 0.24 |
| T2 | -0.20 | 0.24 | -0.30 | 0.07 |
| T3 | -0.27 | 0.20 | -0.58 | 1.8e-4 |
| T4 | -0.09 | 0.60 | -0.25 | 0.13 |
| T5 | 0.42 | 0.01 | 0.14 | 0.44 |
| T6 | -0.24 | 0.15 | -0.63 | <0.0001 |
2) Cronologie isotopiche
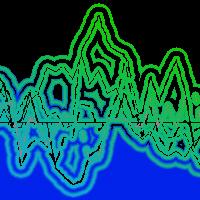
Analizzando i risultati relativi ai valori di composizione isotopica di carbonio e di ossigeno misurati su gruppi di tre anelli consecutivi si nota che nel plot posto a quota più elevata, il trend del δ18O e, soprattutto, del δ13C non subisce grosse variazioni, né evidenzia un segnale di disturbo da parte delle emissioni provenienti dall’autostrada (Fig. 5).
Fig. 5 - Andamento del δ13C e del δ18O misurati su gruppi di tre anelli consecutivi di alberi di abete rosso in due differenti plots lungo l’autostrada A2 nei pressi di Erstfeld.
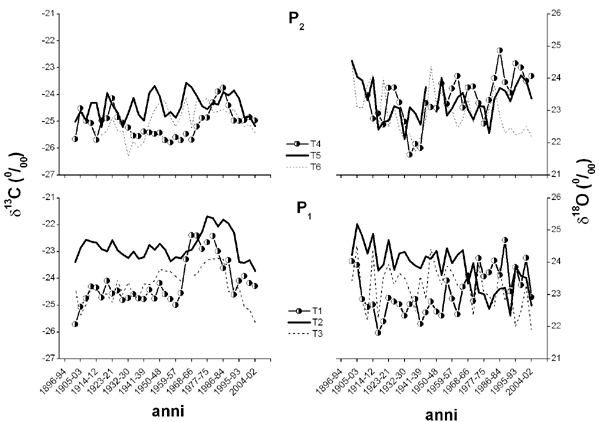
Nel plot alle pendici della parete rocciosa, invece, l’andamento del δ13C è concorde per tutte le piante, mentre più variabile risulta quello del δ18O. In particolare, l’albero più vicino all’autostrada (indicato con T1), mostra un evidente aumento del δ13C e, meno pronunciatamente, del δ18O, a partire dagli anni ’60, fino agli anni ’80. Questo risultato potrebbe indicare una minore attività stomatica ([2], [17]), con conseguente riduzione delle perdite di acqua per traspirazione, ed una maggiore efficienza dell’attività assimilativa, suggerendo un’elevata efficienza d’uso dell’acqua. Una delle spiegazioni di questa risposta positiva della pianta potrebbe essere in parte legata all’aumento delle emissioni di NO2 registrato in Svizzera proprio a partire dagli anni ’60 (Fig. 2). In uno studio condotto da Siegwolf et al. ([18]), infatti, è stato osservato un aumento del δ13C per piante di pioppo sottoposte a fumigazioni di NO2. La maggiore concentrazione di NO2 potrebbe avere un duplice effetto: fertilizzante da una parte, stimolando l’assimilazione di CO2 e tossico dall’altra parte, contribuendo ad una maggiore sensibilità e, quindi, chiusura stomatica.
L’albero T2 mostra un simile andamento del δ13C e del δ18O fino agli anni ’70 dopodiché, ad un aumento del δ13C corrisponde una diminuzione del δ18O. Il minore arricchimento isotopico dell’ossigeno suggerirebbe un elevato tasso traspirativo, accompagnato da un maggiore “Peclet effect” ([2]). Questo potrebbe indicare una minore sensibilità stomatica alle emissioni si NO2 per quest’albero che si trova più distante dall’autostrada rispetto a T1. I valori più elevati di δ13C e di δ18O nei primi 50 anni per questo albero indicherebbero una ridotta attività stomatica probabilmente indotta da una scarsa disponibilità idrica per la pianta, dovuta o alla particolare tipologia di suolo, a matrice prevalentemente rocciosa, o ad un poco sviluppato apparato radicale.
Per l’albero T3 si osserva solo un lieve cambiamento dell’andamento dei valori di δ18O e, soprattutto di δ13C. L’impatto delle emissioni di NOx derivanti dal traffico sembra, quindi, attenuarsi man mano che ci si allontana dall’autostrada, come osservato anche per gli aghi.
Conclusioni
Nelle aree di studio di Faido e Brittnau, gli alberi di abete rosso più vicini all’autostrada hanno mostrato un chiaro arricchimento isotopico in 15N, confermando la possibilità per le piante di assorbire l’azoto anche per via stomatica. È interessante osservare come gli alberi più vicini all’autostrada fungano da barriera, diminuendo l’impatto delle emissioni di NO2 legate alla circolazione dei veicoli, procedendo lungo il transetto, verso l’interno della foresta. Successive analisi su carote estratte dagli alberi considerati per il campionamento degli aghi consentiranno di capire se e con quale entità il segnale osservato negli aghi possa essere trasferito negli anelli di accrescimento.
Dallo studio retrospettivo sugli anelli di accrescimento degli alberi nei pressi di Erstfeld è emerso un segnale interessante, attribuibile in parte al possibile impatto dell’aumento delle emissioni di NOx registrato in Svizzera a partire dagli anni ’60. Questo segnale non è tanto visibile nelle ampiezze anulari (ad eccezione della pianta T1) quanto nei valori di composizione isotopica dell’ossigeno e, soprattutto del carbonio. In quest’ ultimo caso, infatti, il trend crescente di δ13C osservato tra gli anni ’60 e ’80 per le piante T1 e T2 denoterebbe una maggiore attività fotosintetica, quindi un aumento dell’efficienza d’uso dell’acqua per queste piante. Questo risultato suggerirebbe un effetto di fertilizzazione da parte delle emissioni di NOx (e probabilmente anche di CO2) derivante dalla circolazione dei veicoli. L’andamento crescente dei valori di δ18O osservato negli anelli legnosi della pianta T1, più vicina all’autostrada, indica, tuttavia, una riduzione dell’attività stomatica, suggerendo un effetto tossico da parte delle emissioni di NO2.
Questo studio conferma la potenzialità della misura degli isotopi stabili di carbonio, ossigeno ed azoto quale utile strumento per il monitoraggio dell’impatto delle deposizioni di azoto sulla risposta fisiologica degli alberi negli ecosistemi forestali.
Ringraziamenti
L’attività di ricerca è stata svolta da M.R.G. durante il periodo di stage presso il Laboratory of Atmospheric Chemistry - Stable Isotopes Group (Paul Scherrer Institute) - nell’ambito del Dottorato di Ricerca Internazionale in “Crop Systems, Forestry and Environmental Sciences” (Università degli Studi della Basilicata). Si ringrazia l’Istituto Federale di Ricerca per la Foresta, la Neve e il Paesaggio (WSL) per il campionamento degli aghi e per l’utilizzo del laboratorio di dendrocronologia; la Dr. Maya Jaeggi, il Prof. Marco Borghetti e il Prof. Antonio Saracino per il supporto scientifico. La presente ricerca è stata in parte finanziata dall’Ufficio Federale dell’Ambiente - UFAM (Bundesamt für Umwelt - BAFU).
References
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar