Observations on the felling of standards in oak coppices, Central Italy
Forest@ - Journal of Silviculture and Forest Ecology, Volume 3, Pages 397-406 (2006)
doi: https://doi.org/10.3832/efor0390-0030397
Published: Sep 20, 2006 - Copyright © 2006 SISEF
Research Articles
Guest Editors: 5° SISEF Congress (Grugliasco, TO - 2005)
« Forests and Society - Changes, Conflicts, Sinergies »
Collection/Special Issue: E. Lingua, R. Marzano, G. Minotta, R. Motta, A. Nosenzo, G. Bovio
Abstract
Recently, in the coppices of Tuscany and many other regions of central Italy, a large number of standards are commonly released, often leaving all the oldest ones. This practice is linked to the concern for inability to sprout by the old standards stumps. The research was carried out in two coppices with standards of Quercus cerris located near San Gimignano (Siena, Italy), and harvested in the winter 2002/2003. The research focus on the evalutation of the effects of the felling of both different age standards (2, 3 or more rotations) and shoots (1 rotation); the stumps vitality and the shoots growth were studied. The results, two years after logging, show little differences about vitality between 2-3 rotations standards stumps and shoots stumps, even if the mortality of the oldest standards is highest (about 20%). Significative differences are observed as related to the increment both in height and diameter of the shoots. The performances are different in each plot, however behaviour of standards are equal or higher than shoots. The results suggest that the low mortality of the 2-3 rotations standard stumps is wide compensated by growth increase of theirs shoots, therefore the harvest of older standard can enhance the development of the shoots by limiting the shading effect carried out by their crowns.
Keywords
Coppice, Standards, Release, Shoot, Stump, Re-growth, Vitality, Tuscany
Introduzione e scopi
Negli ultimi anni, nel governo dei boschi cedui in Toscana così come in altre regioni dell’Italia centrale, si è manifestata la tendenza a rilasciare un elevato numero di matricine, anche molto superiore a quello previsto dalle vigenti leggi. L’intensificazione della matricinatura è stata attuata, spesso, attraverso il rilascio in piedi di quasi tutte le matricine più vecchie, oltre che di numerosi allievi. Diverse sono le motivazioni che spingono in questa direzione, dalla tutela idrogeologica a quella paesaggistica, compreso il tentativo di attuare una sorta di mascheramento degli effetti del taglio nei periodi immediatamente successivi all’intervento. Inoltre, questa pratica è anche la conseguenza di atteggiamenti prudenziali da parte degli utilizzatori, motivati, tra l’altro, dalla preoccupazione del mancato ricaccio delle matricine di più turni.
La scelta delle matricine è stata da sempre considerata l’azione caratterizzante il governo a ceduo e a ceduo composto ([21], [11], [20], [4], [22], [9]). Il ruolo delle matricine nei boschi cedui italiani ha subito variazioni anche in seguito al variare delle condizioni socio-economiche ([26], [25]).
Il ceduo composto (un tempo si distingueva “la fustaia sopra ceduo” ed “il ceduo sotto fustaia”, a seconda che prevalesse l’una o l’altra componente), sia pure con critiche e dubbi ([21], [4]) trovava una sua giustificazione nella diversificazione dei prodotti legnosi offerti e dall’importanza delle matricine quali produttrici di seme appetito dal bestiame. Ad oggi queste funzioni hanno perso quasi completamente la loro importanza e il ceduo composto viene spesso assimilato ad un ceduo intensamente matricinato, dal quale si differenzia per la presenza di matricine di più classi d’età.
Attualmente, per effetto dell’allungamento dell’intervallo tra le utilizzazioni, le matricine di 3 turni hanno oltre 70-75 anni (invece di 45-50 di quando i turni erano più brevi e regolari), e l’aumento dell’età, in particolare nelle specie quercine, è indicato in letteratura come uno dei possibili fattori che determinano la diminuzione o la cessazione della facoltà pollonifera.
Numerosi Autori hanno evidenziato che la forte copertura esercitata dalle chiome delle matricine può ridurre considerevolmente l’accrescimento dei nuovi polloni, provocando una forte diminuzione di vitalità - e conseguentemente di produzione - della componente ceduo (tra gli altri: [21], [11], e più recentemente: [3], [22], [2], [6], [9]).
Il presente lavoro si propone di valutare gli effetti del taglio delle matricine, prendendo in considerazione sia la capacità di ricaccio sia la vitalità dei polloni, confrontando il comportamento delle matricine e delle ceppaie di un turno.
Materiali e metodi
Ambiente di studio
Le osservazioni sono state condotte in due differenti boschi del Comune di San Gimignano, in provincia di Siena, a prevalenza di cerro (Quercus cerris L.), ambedue classificati come cedui composti ed ascrivibili alla cerreta mesoxerofila secondo la tipologia di Mondino & Bernetti ([18]).
Dati climatici di lungo periodo di San Gimignano indicano una piovosità media annua di 836 mm, con massimo a novembre (108 mm) e minimo a luglio (28 mm); temperatura media annua di 14.7°C, il mese più freddo è gennaio (4.5°C), il più caldo è luglio (25°C). Il clima può essere definito come mesotermico umido a carattere mediterraneo (Csa), secondo la classificazione di Köppen.
Il primo dei due boschi, dove è ubicata la prima area di saggio (A1), si trova sul versante Nord del Poggio del Comune, in località Settefonti, ad una quota di circa 400 m s.l.m. (Fig. 1). Il bosco si trova su terreno con modestissima pendenza, esposizione Nord. Il substrato geologico è rappresentato da conglomerati e ghiaie prevalentemente calcarei con sabbie e sabbie argillose, con suoli profondi e privi di scheletro.
Fig. 1 - Area 1: panoramica dopo il taglio.

La seconda area (A2) si trova in località Marcignanella, ai confini con il Comune di Certaldo, ad una quota di 100-130 m s.l.m., esposizione Nord (Fig. 2). Il substrato geologico è rappresentato da sabbie stratificate con argille e ciottoli, di ambiente litoraneo o salmastro, con suoli profondi e privi di scheletro. La pendenza media è circa il 30%. Nella zona di rilievo il terreno presenta le tracce di una antica sistemazione a gradoni, in parte scomparsa. Fino a circa un secolo fa il terreno era destinato a vigneto (con viti maritate all’olmo), con successiva fase a pascolo arborato (Di Benedetto, com. pers.).
Fig. 2 - Area 2: panoramica dopo il taglio.

Le utilizzazioni sono avvenute durante la stagione silvana 2002/03, ad opera di ditte locali.
Nelle due aree, per motivi diversi, è stato rilasciato un numero di matricine inferiore a quello previsto dall’attuale Regolamento Forestale della Toscana (150 ad ettaro). Sono state rilasciate al taglio circa 120 matricine ad ettaro nell’area 1 e circa 80 matricine ad ettaro nell’area 2. Nel primo caso (Settefonti) si è trattato di un taglio eseguito in seguito a vecchie autorizzazioni precedenti l’entrata in vigore del Regolamento, con prescrizioni comunque verificate e approvate dal locale comando CFS, mentre nel secondo caso (Marcignanella) si è trattato di una violazione delle norme, successivamente denunciata e perseguita legalmente.
La scelta delle aree è stata effettuata dopo l’utilizzazione; di conseguenza la caratterizzazione delle aree è stata effettuata sulla base della composizione specifica desumibile dalle ceppaie e dalle matricine.
Le aree di studio
In ciascuna delle due zone di studio è stata realizzata un’area di saggio di 2400 m2 (60 x 40 m) all’interno della quale per ogni ceppaia è stata rilevata la specie, per ogni matricina è stato rilevato: specie, diametro a 1.30m, altezza e proiezione a terra della chioma (stimata misurando quattro raggi ortogonali orientati in direzione dei punti cardinali) ed è stata attribuita una classe di età (in turni) in base ad un esame visivo delle caratteristiche della chioma (profondità, ampiezza, angolo di inserzione dei rami principali) e del fusto. Le caratteristiche della matricinatura sono riassunte nella Tab. 1 e nella Fig. 3.
Tab. 1 - Caratteristiche della matricinatura.
| Caratteristica | Turni | Area 1 | Area 2 |
|---|---|---|---|
| Num. ha-1 | T1 | 33 (27.6%) |
25 (31.6%) |
| T2 | 71 (58.6%) |
25 (31.6%) |
|
| >= T3 | 17 (13.8%) |
29 (36.8%) |
|
| TOT | 121 | 79 | |
| Altezza media, m (±SD) | - | 16.7 (± 2.1) |
16.8 (± 3.8) |
| Copertura, m2 ha-1 | T1 | 898.8 (13.6%) |
410.4 (9.8%) |
| T2 | 3515.2 (61.0%) |
1234.7 (29.5%) |
|
| >= T3 | 1344.1 (23.3%) |
2537.8 (60.7%) |
|
| TOT% | 57.6 | 41.8 | |
| Età, anni | T1 | 24 | 24 |
| T2 | 48 | 42 | |
| >= T3 | 70-75 | 70-105 |
Fig. 3 - Matricine: distribuzione in classi diametriche. Pannello superiore: Area 1 (Settefonti); pannello inferiore: Area 2 (Marcignanella).
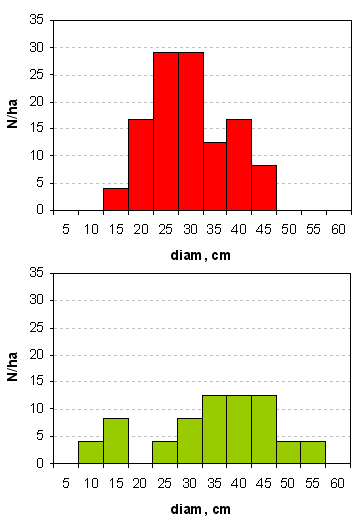
Su tutta la superficie interessata dall’utilizzazione sono state campionate casualmente alcune ceppaie di cerro che sono state localizzate planimetricamente ed etichettate con targhetta metallica. Le ceppaie sono state distinte in categorie in funzione dell’età (turno -T-, determinato tramite analisi degli anelli sulle ceppaie), ovvero: ceppaie di polloni (T1), di matricine di 2 turni (T2) e di 3 o più turni (T3). Sono state campionate per A1: 30 ceppaie di T1, 31 di T2 e 12 di T3, mentre per A2: 30 ceppaie di T1, 6 di T2 e 30 di T3.
Nell’area 1 il numero ad ettaro di ceppaie di cerro è 525, 163, 46 rispettivamente per T1, T2 e T3, mentre le ceppaie di altre specie (acero campestre, biancospino, carpino nero, corniolo, ligustro, orniello, pero selvatico, sorbo ciavardello, sorbo degli uccellatori) sono 1900 ad ettaro.
Nell’area 2 il numero delle ceppaie ad ettaro di cerro ad ettaro risulta di 279, 13, 63 rispettivamente per T1, T2 e T3, mentre le ceppaie di altre specie (acero campestre, biancospino, corniolo, ligustro, olmo campestre, orniello, pero selvatico, prugnolo, sorbo degli uccellatori) risultano oltre 3100 ad ettaro.
Rilievi sulla rinnovazione agamica
Sono stati effettuati 5 rilievi nel 2003 (15/03; 4/06; 23/06; 4/09; 21/10) e 3 nel 2004 (18/03; 4/09; 20/10).
Ad eccezione del primo rilievo del 2003, su ciascuna ceppaia è stato rilevato:
- l’altezza del pollone dominante (Hd, altezza del pollone più alto della ceppaia);
- il numero totale di polloni (Nt),
- il numero di polloni dominanti (Nd, numero di polloni con altezza superiore ai 2/3 di Hd).
Nell’ultimo rilievo (ottobre 2004) è stato misurato anche il diametro (d) a 10 cm dalla base del pollone dominante.
Elaborazione dei dati
Le elaborazioni dei dati sono state eseguite con i comuni test statistici (chi-quadro, ANOVA), considerando come fonti di variazione le categorie di ceppaia: di polloni, (T1, indicate con “p”), di matricine (riunendo T2 e T3, indicate con “M”). Questa scelta è dovuta alla considerazione che per alcune categorie (T3 nell’area 1 e T2 nell’area 2) il limitato numero di ceppaie presenti non consente un adeguato confronto; inoltre anche dal punto di vista applicativo, il regolamento forestale della Regione Toscana non fa distinzione fra le matricine di due o più turni[1].
Risultati
Vitalità delle ceppaie
Nell’autunno del 2003, in A1 aveva ricacciato il 92% delle ceppaie, quota che è rimasta praticamente invariata l’anno successivo, senza differenze significative fra le diverse categorie (2004: χ2[1] = 0.88; n.s.). Alla fine del 2004 le ceppaie vitali sono l’87% e il 93% rispettivamente per p ed M, e tra le matricine l’assenza di ricaccio si osserva soprattutto tra quelle di almeno tre turni
In A2, alla fine del primo anno tutte le ceppaie p hanno ricacciato mentre tra le matricine il 32% non ha ricacciato (χ2[1] = 8.71; p<0.01); questa percentuale scende nel secondo anno al 23%, pur restando significativamente superiore a quella dei polloni (χ2[1] = 6.67; p<0.01); anche in quest’area la mortalità maggiore è a carico delle ceppaie delle matricine più vecchie.
Nell’area 1, durante il secondo anno alcune ceppaie p, che avevano ricacciato l’anno precedente, sono morte per cui, complessivamente, alla fine del secondo anno di vegetazione dopo il taglio, in ambedue le aree era vitale il 90% circa delle ceppaie.
Accrescimento dei polloni
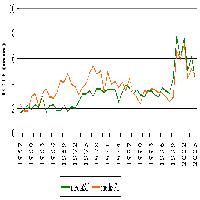
La Fig. 4 mostra l’accrescimento medio dei polloni dominanti nel corso dei due anni di rilievo.
Fig. 4 - Variazioni di Hd media nei due anni di rilievo. Pannello superiore: Area 1 (Settefonti); pannello inferiore: Area 2 (Marcignanella).
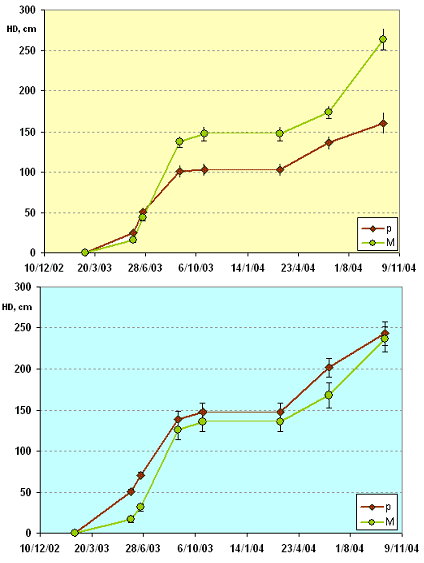
Dall’esame dei grafici si può identificare una tendenza comune alle due aree che vede le ceppaie p presentare gli incrementi medi maggiori nel periodo primaverile, mentre le ceppaie M crescono maggiormente nel periodo estivo-autunnale.
In Tab. 2 sono riassunti i risultati relativi al test dell’ANOVA applicati ai valori medi di altezza dominante alla fine di ciascun anno di rilievo. Nell’area 1 emergono differenze molto significative (p<0.01), con valori medi superiori per le ceppaie derivanti dal taglio dalle matricine, con differenze che si fanno più marcate alla fine del secondo anno. Nell’area 2 i valori medi di altezza dominante non differiscono in maniera significativa fra le categorie confrontate, con trascurabili differenze nelle due annate a favore delle ceppaie derivanti da polloni.
Tab. 2 - Test dell’ANOVA relativi ai valori medi di Hd 2003 e 2004; p ed M indicano le categorie di ceppaia (vedi Materiali e metodi).
| Aree | Anno | F di Fisher | gl | Significatività | Hd media, cm | |
|---|---|---|---|---|---|---|
| p | M | |||||
| A1 | 2003 | 10.76 | 1; 64 | p<0.01 | 108.1 | 147.4 |
| 2004 | 27.87 | 1; 64 | p<0.01 | 160.8 | 263.8 | |
| A2 | 2003 | 0.16 | 1; 56 | n.s. | 147.7 | 141.4 |
| 2004 | 0.74 | 1; 57 | n.s. | 243.5 | 236.4 | |
Il confronto tra le due aree (ANOVA, Tab. 3) evidenzia un comportamento diverso fra le categorie di ceppaia: a fronte di valori medi di Hd paragonabili per i ricacci delle ceppaie originate dalle matricine (ma con valori leggermente superiori per l’area 1), si osservano differenze molto significative per quelli delle ceppaie originate da polloni, con valori medi questa volta significativamente superiori per l’area 2.
Tab. 3 - Differenze tra le aree. Test dell’ANOVA relativi ai valori medi di Hd 2004.
| Categorie | F di Fisher | gl | Significatività | Hd media, cm | |
|---|---|---|---|---|---|
| Area 1 | Area 2 | ||||
| p | 18.419 | 1; 64 | p<0.01 | 160.8 | 243.5 |
| M | 1.762 | 1; 67 | n.s. | 263.8 | 236.4 |
Diametro del pollone dominante
In Tab. 4 sono riassunti i risultati relativi al test dell’ANOVA applicati ai valori medi del diametro del pollone dominante (d). I risultati sono in linea con quelli relativi all’altezza dominante: le differenze sono significative a favore delle matricine nell’area 1, non significative nell’area 2, con valori leggermente superiori per le ceppaie p.
Tab. 4 - Test dell’ANOVA relativi ai valori medi di d 2004; p ed M indicano le categorie di ceppaia (vedi Materiali e metodi).
| Aree | F | gl | Significatività | d Medio, mm | |
|---|---|---|---|---|---|
| p | M | ||||
| A1 | 28.99 | 1; 64 | p<0.01 | 14.7 | 27.2 |
| A2 | 0.54 | 1; 58 | n.s. | 27.6 | 25.9 |
Numero di polloni
Il numero totale medio di polloni per ceppaia, così come quello medio di polloni dominanti per ceppaia non presentano differenze significative tra le categorie esaminate (Tab. 5)
Tab. 5 - Test dell’ANOVA relativi ai valori medi di valori medi di Nt e Nd 2004; p ed M indicano le categorie di ceppaia (vedi Materiali e metodi).
| Aree | Variabile | F di Fisher | gl | Significatività | Nt Medio | |
|---|---|---|---|---|---|---|
| p | M | |||||
| A1 | Nt 2004 | 0.50 | 1; 63 | n.s. | 11.6 | 12.7 |
| A2 | 0.56 | 1; 49 | n.s. | 13.9 | 12.3 | |
| A1 | Nd 2004 | 1.04 | 1; 63 | n.s. | 3.2 | 3.6 |
| A2 | 0.16 | 1; 49 | n.s. | 4.3 | 4.2 | |
Discussione
Nel definire l’età delle matricine si è fatto riferimento al numero di turni, come riportato nella legislazione forestale e adottato nella consuetudine. In realtà occorre considerare che questo sistema aveva una sua giustificazione ed una maggiore rispondenza con l’età cronologica delle matricine quando i turni erano cadenzati in maniera più regolare di quanto avvenga oggi. Con la sospensione delle utilizzazioni nella seconda metà del XX secolo e con la tendenza attuale all’allungamento del turno, ogni volta che si parla di numero di turni è necessario considerare che ci si può riferire anche a situazioni molto diverse tra loro. Quindi è possibile che, oggi, matricine “di 3 turni” abbiano un’età uguale a quella che, fino a qualche decennio fa (o in cedui trattati regolarmente) avrebbero avuto matricine di 4 o più turni.
Le osservazioni sono state effettuate su due aree di saggio che, pur presentando minime variazioni a livello di clima e substrato, differiscono per le caratteristiche strutturali dei popolamenti, effetto sia della loro storia sia della modalità dell’ultima utilizzazione. Queste differenze non consentono la generalizzazione dei risultati, tuttavia i due casi di studio sono entrambi espressivi di condizioni molto frequenti nella zona oggetto d’indagine.
Nell’area 1 la presenza del bosco, e la forma di governo a ceduo composto, risalgono a tempi lontani, tanto da poter affermare che anche le matricine più vecchie siano passate attraverso le varie fasi descritte in letteratura (allievi, matricine di un turno, etc.).
L’area 2, secondo testimonianze fornite dai proprietari, fino nei primi decenni del ’900, era destinata a pascolo arborato. In quest’area è probabile che le matricine più vecchie abbiano avuto, nel periodo giovanile, condizioni di crescita diverse da quelle dell’altra area, benché si siano potuti documentare, attraverso l’analisi della sezione di taglio, almeno tre turni di ceduazione.
La vitalità delle ceppaie originate dal taglio delle matricine è abbastanza alta, anche se inferiore a quella delle ceppaie di polloni. Tra le matricine, inoltre, si osserva la mortalità più alta per le ceppaie più vecchie (3 turni e più), mentre le matricine di 2 turni, in particolare nell’area 1 (dove l’ampiezza del campione permette una valutazione migliore), si sono dimostrate più vitali. Questa tendenza era già stata segnalata da La Marca et al. ([16]) in boschi cedui della stessa specie.
Per quanto riguarda l’entità del fenomeno, il tasso di mortalità rientra nei limiti osservati nella normale gestione di un ceduo composto; i risultati, ed in particolare le differenze tra le diverse categorie, sono paragonabili a quelli riportati da La Marca et al. ([15]).
È importante evidenziare che alcune matricine hanno ricacciato solo nella primavera/estate del secondo anno, anche se la maggioranza di esse, così come la totalità di quelle originate da polloni, ha ricacciato durante il primo anno. Dal punto di vista applicativo queste osservazioni suggeriscono che un esame della vitalità delle ceppaie effettuato pochi mesi dopo il taglio potrebbe portare ad errori di valutazione.
Per quanto riguarda la vigoria dei polloni, nelle due aree è stato osservato un comportamento differente tra le ceppaie delle diverse categorie.
Nell’area 1 le ceppaie originate dal taglio delle matricine presentano le migliori performances, con valori molto più elevati rispetto alle altre per quanto riguarda l’altezza del pollone dominante.
Nell’area 2, invece, questa tendenza non trova conferma; ma il comportamento delle ceppaie più vecchie è comunque paragonabile a quello delle più giovani, con analoghi accrescimenti longitudinali.
Numerosi studi hanno evidenziato che la capacità di ricaccio è strettamente correlata alla vigoria dell’individuo prima del taglio ([13], [24], [12], [1], [19], [14]). Nel nostro studio sono soprattutto le ceppaie delle matricine più giovani ad essere più vigorose; probabilmente l’essere cresciute in condizione radiative ottimali durante l’ultimo turno (posizione dominante) può aver influito positivamente su questo fenomeno. Questo effetto, tuttavia, non si verifica per le matricine più vecchie, confermando che al crescere dell’età tende a diminuire la vitalità delle ceppaie (nessuna emissione di polloni, minore vigoria degli stessi), trovando ampia conferma in letteratura (tra gli altri: [8], [22], [5], [14]).
Le differenze osservate fra le due aree sembrano imputabili soprattutto alla diversa intensità dell’intervento e quindi al diverso grado di copertura dopo l’intervento: nell’area 2 sono state rilasciate meno matricine (42% di copertura dell’area 2 contro il 58% dell’area 1), con relativo minore ombreggiamento delle ceppaie. Sembra che l’aduggiamento limiti maggiormente l’accrescimento dei ricacci delle ceppaie più giovani, come suggerisce il confronto tra le due aree.
Per quanto riguarda i ritmi di accrescimento, quasi tutte le ceppaie hanno presentato più flussi di accrescimento nella stessa stagione. Questa caratteristica è tipica delle querce, in particolare nel periodo giovanile ([17], [13], [23], [10], [12], [7]).
In ambedue le aree si può notare che le p mostrano gli incrementi maggiori nel periodo primaverile, a differenza di delle ceppaie M che crescono di più nella seconda metà del periodo vegetativo. Il maggiore accrescimento primaverile sembra imputabile ad un diverso comportamento fenologico delle due categorie di ceppaia. I ricacci delle ceppaie dei polloni entrano prima in vegetazione ed usufruiscono di un periodo vegetativo più lungo rispetto a quelli delle ceppaie M, dato che l’inizio della stasi vegetativa estiva avviene nello stesso periodo per entrambe le categorie. La ripresa vegetativa tardo-estiva avviene quasi simultaneamente e i ricacci delle ceppaie delle matricine si accrescono vigorosamente. Un simile comportamento è stato osservato in cedui di leccio da Giovannini et al. ([12]) e in un ceduo di cerro nei pressi di Firenze (Giovannini, in preparazione).
Il diametro del pollone dominante ed il numero di polloni per ceppaia sono dati che, a pochi anni dal ricaccio, sono di minore utilità ai fini del confronto.
I dati del diametro seguono lo stesso andamento osservato per l’accrescimento longitudinale, cosa per altro abbastanza logica vista la nota correlazione tra i due parametri.
Il numero di polloni, infine, non ha dato luogo in nessuna delle due aree a differenze statisticamente significative. Anche in questo caso il risultato è prevedibile dato che nelle prime fasi di sviluppo non si sono ancora instaurati fenomeni di concorrenza tra i polloni tali da determinare una selezione tra gli stessi, né tra i polloni dominanti, né tra tutti gli altri.
Conclusioni
Il ceduo composto rappresenta in diverse zone dell’Italia Centrale, ancora oggi, una forma di governo attuata e mantenuta sia come conseguenza dell’uso tradizionale dei boschi, sia per l’alto valore paesaggistico che viene loro attribuito.
La tendenza attuale, nella gestione di questi soprassuoli, va verso l’aumento della densità della componente a fustaia, attuata rilasciando un numero crescente di matricine. Questo atteggiamento è dovuto, almeno in parte, al timore del mancato ricaccio delle matricine più vecchie.
Pur non potendo generalizzare i risultati, queste prime osservazioni non sembrano avvalorare questi timori: la mortalità delle ceppaie originate dal taglio delle matricine appare accettabile e in linea con quanto riportato in letteratura. Lo sviluppo dei polloni di queste ceppaie è superiore o paragonabile a quello delle ceppaie più giovani, tanto che si può affermare che possa compensare la perdita di alcune ceppaie per mancato ricaccio.
L’aumento dell’età delle matricine, al di là del numero di turni, sembra comunque avere una certa influenza negativa sulla capacità di ricaccio delle ceppaie e sul vigore dei nuovi polloni.
Per mantenere la fisionomia del ceduo composto, e soprattutto per garantire la produttività del ceduo, è quanto mai opportuna una attenta regolazione della copertura con il taglio delle matricine più vecchie, per evitare l’aduggiamento dei polloni (e anche della eventuale rinnovazione da seme).
Ringraziamenti
Un ringraziamento particolare a Stefano Ignesti e Gaetano Di Benedetto, del Corpo Forestale dello Stato per le informazioni e per l’aiuto nella scelta delle aree di studio. Lavoro eseguito con fondi di ateneo ex-60%.
References
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar