Carbon loss through respiratory processes in terrestrial ecosystems: evidences from recent literature
Forest@ - Journal of Silviculture and Forest Ecology, Volume 3, Pages 317-319 (2006)
doi: https://doi.org/10.3832/efor0405-0003
Published: Sep 20, 2006 - Copyright © 2006 SISEF
Commentaries & Perspectives
Abstract
Papers in the recent literature document advances in respiration in both methods and results. A general discussion is proposed about the common features of the latest studies, the current state of the knowledge of the respiration processes as well as some need for future researches.
Keywords
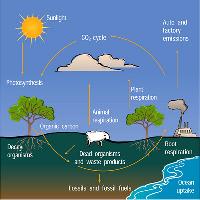
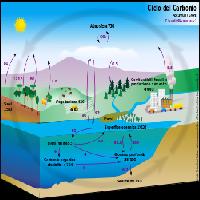
Il carbonio si trova in natura combinato in un numero straordinariamente grande di composti. Nelle sue forme organiche il carbonio rappresenta la fonte primaria di energia per tutte le forme di vita eterotrofiche le quali, utilizzatolo come substrato per le proprie esigenze metaboliche, lo restituiscono all’atmosfera attraverso un insieme di processi definiti nel complesso come respirazione. Fotosintesi e respirazione alimentano un ciclo che si rinnova nella biosfera da migliaia di anni e che negli ecosistemi terrestri si è mantenuto nel tempo in uno stato di equilibrio stazionario.
Sebbene la condizione di equilibrio sia attualmente soddisfatta a scala planetaria, a livello ecosistemico è ormai ampiamente dimostrato che perturbazioni quali l’aumento della CO2 atmosferica, le attività antropiche di uso del suolo, la variabilità climatica e fattori naturali di incidenza locale (soprattutto incendi) determinino significative fluttuazioni interannuali nel bilancio dei processi di assimilazione e respirazione. Ad oggi gli ecosistemi terrestri sono considerati sinks per il carbonio atmosferico ma è evidente che la variabilità dei processi di scambio del C tra ecosistemi terrestri ed atmosfera attribuisce a questa potenzialità ampi margini di incertezza complicando la possibilità di prevedere con sufficiente accuratezza la risposte degli ecosistemi terrestri ai cambiamenti del clima attuali e futuri.
Valentini et al. ([6]), ricostruendo il bilancio del carbonio di 15 foreste europee riportano che la respirazione rappresenta il fattore principale nel determinare lo scambio netto dell’ecosistema (NEE). Sulla scorta di questo ed altri contributi lo studio dei processi respiratori ha ricevuto negli ultimi anni un’attenzione crescente nell’ambito degli studi dedicati alle dinamiche di scambio del carbonio tra ecosistemi terrestri ed atmosfera ma nonostante ciò la respirazione continua ad essere ad oggi un processo per molti aspetti poco conosciuto.
Uno dei punti più critici fa riferimento al fatto che sebbene siano stati identificati con ragionevole confidenza i fattori ambientali che controllano la respirazione (temperatura, umidità, quantità e qualità del C-substrato) non si è attualmente in grado di definire una teoria organica che spieghi come questi fattori interagiscono tra loro nel determinare le perdite totali di C dell’ecosistema.
Il problema maggiore è legato alla eterogeneità biologica dei processi autotrofi ed eterotrofi che concorrono a generare le perdite di C dell’ecosistema, i quali, a parità di fattori, possono presentare risposte funzionali differenti. È noto infatti che la respirazione dell’ecosistema comprende sia la “respirazione delle piante” (autotrofa) che la “respirazione del suolo”, costituita a sua volta da processi autotrofi (respirazione radicale) ed eterotrofi (ossidazione dei substrati C che includono la lettiera superficiale, sotterranea e la sostanza organica del suolo ad opera della catena del detrito); in foresta la respirazione del suolo contribuisce fino al 65% dell’emissione totale del C.
Le difficoltà nel definire una base teorica di riferimento per la respirazione sono anche frutto di approcci sperimentali che hanno scontato nel passato un certo carattere “subalterno” dei processi respiratori negli studi finalizzati a valutare il bilancio del C degli ecosistemi. Gran parte di questi studi è stata effettuata infatti con l’obiettivo di contribuire a valutare la produttività netta dell’ecosistema e per questa finalità la respirazione, in particolare quella del suolo, veniva misurata complessivamente, senza l’esigenza di separare il flusso nelle componenti costitutive allo scopo di chiarire le dinamiche di dettaglio dei processi coinvolti. Questo ha fatto sì che la maggior parte dei modelli sviluppati a partire da quei dati sperimentali avesse un carattere empirico (il modello più diffuso prevede la dipendenza della respirazione dalla temperatura del suolo) e forti limitazioni nei tentativi di spazializzazione delle stime o di applicazione in ecosistemi diversi da quelli per i quali erano stato definiti.
Lo sforzo di separare le componenti autotrofe e eterotrofe della respirazione, in particolare del suolo, è relativamente recente e si auspica che possano essere presto disponibili maggiori dati per l’elaborazione e validazione di modelli su base funzionale che migliorino la nostra attuale capacità previsionale sulle perdite di C dal suolo.
A questo proposito, grazie soprattutto agli avanzamenti ottenuti nei metodi sperimentali sono stati ottenuti alcuni interessanti risultati che meritano una riflessione.
Un punto comune che caratterizza questi contributi fa riferimento al fatto che le proporzioni relative delle componenti autotrofe ed eterotrofe della respirazione del suolo possono essere molto variabili e non necessariamente costanti durante le stagioni.
Scott-Denton et al. ([3]) in un esperimento di cercinatura in Colorado riportano che la respirazione autotrofa costituisce dal 31% al 44% della respirazione del suolo ed un andamento stagionale della respirazione del suolo da cui emerge che la componente eterotrofa è più sensibile di quella autotrofa alle variazioni di umidità.
Cisneros-Dozal et al. ([1]) in un querceto misto del Tennessee, mediante l’analisi delle firme isotopiche del 14 C stabiliscono che la respirazione etrotrofica, in particolare la respirazione della lettiera, costituisce la maggior parte della respirazione del suolo durante la stagione vegetativa, con variazioni stagionali che riflettono la disponibilità di lettiera ed il suo contenuto idrico. Non osservano nessun pattern stagionale per la respirazione autotrofa che raggiunge il suo massimo contributo nelle condizioni minime di flusso.
Shuur & Trumbore ([4]) mediante la determinazione del decadimento degli isotopi radioattivi del carbonio in popolamenti di Picea mariana in Alaska osservano un contributo della respirazione eterotrofa pari al 47- 63% rispetto a quella autotrofa.
Sebbene i risultati siano stati ottenuti con tecniche diverse, la ricerca delle cause alla base dei contributi misurati ha evidenziato due argomenti comuni che possono essere così riassunti: (1) l’importanza della disponibilità di substrato per la respirazione autotrofa ed in genere dei pattern di allocazione del C prodotto nei processi di assimilazione tra le funzioni del metabolismo della pianta (crescita, mantenimento, respirazione, riserva); (2) l’esistenza di dinamiche di breve e lungo periodo nella respirazione eterotrofa.
Per quanto riguarda il primo punto, Davidson et al. ([2]) attraverso una riflessione teorica sulla sensibilità della respirazione alla variabilità della temperatura dimostrano l’importanza di distinguere un effetto diretto della temperatura, dell’umidità e della disponibilità di substrato sulla respirazione, dall’effetto che le stesse variabili climatiche esercitano indirettamente sulla disponibilità del substrato stesso. Questa riflessione per niente sofistica è un contributo al tentativo di sviluppare una base teorica per la definizione di modelli a base funzionale che riconcilia inoltre alcune evidenze sperimentali quali la difficoltà di correlare la respirazione all’umidità del suolo sotto certe soglie di contenuto idrico del suolo stesso.
Una particolare attenzione è riservata al ruolo dei carboidrati di riserva, una componente della produttività della pianta tradizionalmente poco considerata negli studi dedicati al bilancio del carbonio dell’ecosistema.
Cisneros-Dozal et al. ([1]) riporta che la firma isotopica del C respirato dalle radici presenta delle variazioni all’inizio ed alla fine del periodo vegetativo che possono essere spiegate ipotizzando uno switch tra le sorgenti di C utilizzate nella respirazione autotrofa da carboidrati di riserva a prodotti recenti della fotosintesi.
Shuur & Trumbore ([4]) dimostrano che la firma isotopica del C emesso dalla respirazione di radici escisse è più vecchio di quello recentemente fotosintetizzato il che porta a concludere che probabilmente il substrato respirato dalle radici è stato “riciclato” a partire dai carboidrati di riserva.
Per quanto riguarda la respirazione eterotrofa gli spunti più interessanti che forniscono gli studi più recenti fanno riferimento alla possibilità di chiarire la natura dei processi che controllano i tempi di turnover delle frazioni del C organico del suolo.
Trumbore et al. ([5]) confrontando in due siti sperimentali in Amazzonia Orientale la firma isotopica del C-CO2 presente nella porosità del suolo e dei tessuti delle radici fini suppone che una porzione delle radici fini morte e quindi in decomposizione rimanga nei pools della sostanza organica del suolo per un tempo maggiore di quello previsto e che possa quindi essere prevista la possibilità che le radici fini abbiano un tempo di turnover superiore a quello generalmente assunto.
La nostra attuale conoscenza dei processi di decomposizione è in genere legata all’assunzione semi-empirica che le frazioni del C organico del suolo possono essere ricondotte a pochi comparti che a seconda della cinetica di reazione possono essere distinti in “active, slow, passive, inerte” ma anche in questo caso non si dispone di un teoria organica sul fenomeno “decomposizione” in grado di cogliere gli effetti dei processi coinvolti.
Si è guardato con interesse ad alcuni studi che mostrano come le comunità microbiche siano in grado di utilizzare substrati di C considerati normalmente inerti e che la sensibilità alla temperatura delle componenti del C nel suolo considerate meno reattive possa essere superiore rispetto a quelle cui si attribuisce maggiore reattività, a dimostrazione che molti aspetti del fenomeno ci sono ancora poco chiari.
In definitiva dall’analisi dei contributi più recenti della letteratura si può concludere che siamo in una fase della ricerca sulla respirazione in cui i metodi sono maturi per superare l’approccio classico della misure del flusso di CO2 emesso “tal quale” dal suolo ed indirizzare gli sforzi verso studi che chiariscano gli aspetti funzionali dei processi coinvolti nella respirazione del suolo con l’obiettivo ultimo di superare il carattere empirico dei modelli attraverso per es. l’introduzione di nuovi fattori come i controlli enzimatici.
References
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar