Forests, woods, forest plantations
Forest@ - Journal of Silviculture and Forest Ecology, Volume 3, Pages 464-487 (2006)
doi: https://doi.org/10.3832/efor0424-0030464
Published: Dec 18, 2006 - Copyright © 2006 SISEF
Review Papers
Abstract
In protected areas the forest ecosystem management is directed to define the best approaches with high protection levels from ecological, historical, anthropological and landscape point of view. The conservation purposes have to be taken in consideration to not disturb the natural and functional processes, and therefore any forest human activity has to be done. Through a detailed analysis of the relations among functionality, stability, productivity and genetic diversity, the statement of the reasons for application of close-to-nature silviculture is described and discussed. Some specific silvicultural systems are illustrated on the basis of very large quantity of data and information originated from researches carried out for long time. A major challenge facing modern silviculture is to reconcile the traditional objectives of timber production with the demand for multifunctional forest ecosystems which arises from the society. The preservation of the functionality is strictly related to the forest genetic pool which is the basis of biodiversity, as it represents the basis for adaptation and survival of species and individual.
Keywords
Protected areas, forest ecosystem functionality, genetic diversity, close to nature silviculture.
Premessa
Il 6 dicembre del 1991 il Parlamento italiano promulgava la Legge n. 394 “Legge quadro sulle aree protette”. L’importanza della legge risiede soprattutto nella sua valenza generale proponendosi come punto di riferimento per la gestione e uso del territorio di aree protette dettando una normativa, ma anche sanzioni per trasgressioni ovvero indennizzi ed incentivi. In questo senso la legge indica la via maestra per la tutela e l’attuazione di progetti di salvaguardia e di sviluppo. Difatti nel caso dei Parchi Nazionali gli obiettivi di tutela devono essere manifesti nel piano del Parco (art. 12 della suddetta legge): “la tutela dei valori naturali ed ambientali affidata all’Ente parco è perseguita attraverso lo strumento del piano del parco...”.[1]
Le risorse forestali sono soggette a tutela particolare attraverso l’individuazione di linee di gestione eco-sostenibile che devono scaturire dalle relazioni intercorrenti tra valori del bosco, oggetto ed obiettivi di tutela, nonché caratteristiche bio-ecologiche degli ecosistemi forestali.
L’attività antropica, nel corso dei millenni, percorrendo una strada diretta verso il traguardo del proprio benessere ed impostata su uno sfruttamento delle risorse per incrementare lo sviluppo economico, ha di fatto modificato, spesso in modo permanente, la struttura, la densità e il numero delle specie presenti negli ecosistemi.
Nel caso degli ecosistemi forestali[2], che molto probabilmente nelle aree geografiche a condizioni ambientali più favorevoli all’uomo erano i sistemi naturali più ampiamente diffusi, l’impatto di una civilizzazione non sempre ragionata ha determinato due situazioni prevalenti: a) la distruzione della foresta con la sostituzione i) temporanea di ecosistemi più semplificati (produzione agricola) e/o ii) permanente (urbanizzazione) e b) l’utilizzo attraverso il ricorso a modelli più o meno rispettosi della complessità e funzionalità dell’ecosistema bosco (utilizzazioni legnose, pascolo, etc.). In questi casi la distruzione o comunque un uso non corretto della foresta ha portato a situazioni inverosimili di degrado fino alla desertificazione mentre in altri, in cui le condizioni stazionali non rappresentavano, di per sé, limiti insormontabili (condizioni estreme di temperatura, disponibilità idriche e caratteristiche geo-pedologiche), il dinamismo successionale ha consentito la restaurazione di ecosistemi forestali.
D’altra parte è purtroppo noto il fatto che, a livello mondiale, la superficie boscata tende ancora oggi a diminuire e che comunque la produzione legnosa dei popolamenti artificiali attualmente in atto, non potrà coprire il fabbisogno dei paesi ad elevato tasso di industrializzazione che tenderanno comunque a rifornirsi dalle riserve naturali mondiali. Tra l’altro da più parti viene sollevato la preoccupante situazione a livello ambientale del diffuso degrado determinato dalla sistematica distruzione della superficie boscata ad opera di vari fattori tra cui, ad esempio, gli incendi.
Tutto ciò sebbene da oltre mezzo secolo venga ripetuto con insistenza il ruolo svolto dal bosco, l’importanza delle sue funzioni e la necessità di ricorrere ad una gestione sostenibile della risorsa rinnovabile foresta ([31]).
In un recente saggio ([18]) sulle linee guida per la gestione sostenibile delle risorse forestali e pastorali nei Parchi Nazionali, vengono illustrati con estrema chiarezza i fondamenti dottrinali su cui si basa la gestione sostenibile a confronto di una visione basata sullo sfruttamento delle risorse. La diversità di pensiero che porta a riconoscere posizioni differenziate di sostenibilità (sostenibilità molto debole, debole, forte, molto forte) attraverso un rapporto tecnocentrismo/ecocentrismo delimita “la diversa concezione dello sviluppo e del valore attribuito alla natura: strumentale illimitato, strumentale limitato, intrinseco”.
Negli anni 1992 - 1993 due incontri di importanza globale, il Convegno di Rio e la Conferenza di Helsinki, enunciano alcune regole che rappresentano le pietre miliari della definizione delle linee guida per la gestione sostenibile e di quelle per la conservazione della biodiversità.
Infatti è l’improrogabile necessità di individuare un percorso comune per costruire uno sviluppo sostenibile che conduce la Comunità mondiale a riunirsi nel 1992 a Rio de Janeiro. I Paesi aderenti riconoscono che le problematiche ambientali devono essere affrontate in maniera universale e che le soluzioni devono coinvolgere tutti gli Stati. Vengono negoziate e approvate tre dichiarazioni di principi, e firmate due convenzioni globali. Inoltre per sovrintendere all’applicazione degli accordi nasce la Commissione per lo Sviluppo Sostenibile delle Nazioni Unite con il mandato di elaborare indirizzi politici per le attività future e promuovere il dialogo e la costruzione di collaborazioni tra governi e gruppi sociali.
Alla Conferenza delle Nazioni Unite su Ambiente e Sviluppo Vertice della Terra h anno partecipato rappresentanti dei governi di 178 Paesi, più di 100 capi di Stato e oltre 1000 Organizzazioni Non Governative. Sono state sottoscritte 2 convenzioni e 3 dichiarazioni di principi. In particolare:
- l’Agenda 21: il Programma d’Azione per il XXI secolo, che pone lo sviluppo sostenibile come una prospettiva da perseguire per tutti i popoli del mondo;
- la Dichiarazione dei principi per la gestione sostenibile delle foreste sancisce il diritto degli Stati di utilizzare le foreste secondo le proprie necessità, senza ledere i principi di conservazione e sviluppo delle stesse;
- la Convenzione quadro sui cambiamenti climatici cui seguirà la Convenzione sulla Desertificazione, pone obblighi di carattere generale miranti a contenere e stabilizzare la produzione di gas che contribuiscono all’effetto serra;
- la Convenzione quadro sulla biodiversità, che tutela le specie nei loro habitat naturali e riabilitare quelle in via di estinzione;
- la Dichiarazione di Rio su Ambiente e Sviluppo, che definisce, in 27 principi, diritti e responsabilità delle nazioni nei riguardi dello sviluppo sostenibile.
I principi di sviluppo sostenibile sono ripresi successivamente nella Seconda Conferenza Ministeriale, tenutasi ad Helsinki nel 1993. I ministri responsabili per il settore forestale in Europa hanno sottoscritto i Principi Forestali UNCED, accettati a livello internazionale, facendo un passo avanti nella storia del concetto della gestione forestale sostenibile adottando, tra l’altro, la risoluzione H1 “Linee guida generali per la gestione sostenibile delle foreste europee” e la risoluzione H2 “Linee guida generali per la conservazione della biodiversità delle foreste europee”. Queste linee guida generali rappresentano l’impegno politico dei Paesi firmatari della risoluzione e forniscono un indirizzo di politica generale e un obiettivo a lungo termine per soddisfare le richieste europee relative alla multifunzionalità delle foreste in modo coerente con la loro gestione sostenibile e con la conservazione e l’incremento della loro biodiversità.
Nella risoluzione H1 viene formulata una nuova e comune definizione di “gestione forestale sostenibile” la quale indica che “la gestione corretta e l’uso delle foreste e dei terreni forestali deve avvenire nelle forme e a un tasso di utilizzo tali da mantenere la loro biodiversità, produttività, capacità di rinnovazione, vitalità e potenzialità che assicuri ora ed in futuro, rilevanti funzioni ecologiche, economiche e sociali a livello locale, nazionale e globale tali da non comportare danni ad altri ecosistemi”.
Gli ecosistemi forestali
Da un punto di vista generale gli ecosistemi forestali forse più di altri, possono essere considerati “capitali” ad alto tasso di investimento, o “capitali vantaggiosi” in quanto attraverso una gestione appropriata sono capaci di fornire flussi diversificati di servizi che comprendono: produzione di beni (produzione legnosa), processi di supporto alla vita (effetti protettivi), fattori estetici che condizionano la vita (bellezza, serenità ). Per maggiori dettagli vedi Appendice 1.
Nel passato in molte parti del nostro pianeta, ma ancora oggi in altrettante, la produzione primaria di livelli trofici successivi (che si estrinsecano in catene alimentari), rappresenta il principale servizio vitale. Altrove ed in situazioni socio-economiche diverse, la foresta è vista come bene di supporto alla vita (aspetti produttivi) ovvero elemento estetico ed etico che può condizionare il modo di vivere (Box 1, Box 2).
Box 1 - Definizione di Selvicoltura.
La Selvicoltura può essere definita come la Scienza applicata alla coltivazione degli ecosistemi forestali, che, basandosi su informazioni di natura eco-biologica, punta al miglior compromesso tra produzione di beni e servizi e stabilità funzionale. Nella pratica corrente l’azione di maggiore valenza è la regolazione nello spazio e nel tempo della densità degli alberi che comporta l’utilizzazione di massa legnosa.
Box 2 - Razionale.
In accordo a questa scelta operativa, in modo provocatorio, ma sotto certi aspetti molto realistico, potrebbe essere di interesse conoscere dove effettivamente si dispone della libertà eco-biologica di azione che ci garantisca di pervenire con successo a quanto indicato. In primo luogo si dovrebbe disporre di un’ampia messe di dati relativa alla casistica di tipologie eco-stazionali e per ciascuna i limiti che le caratterizzano. Attraverso un’analisi congiunta multivariata per fattori a dominanza crescente (o decrescente) si potrebbe pervenire ad individuare alcune caratteristiche che per valori limite escludono la possibilità di interventi selvicolturali altrove di per sé biologicamente efficaci, per modificare l’ecosistema verso stadi più evoluti e funzionali. Questa analisi potrebbe portare alla individuazione di aree e/o di soprassuoli di non intervento e quindi restringere enormemente la superficie di applicabilità dei modelli colturali.
Non può essere poi sottovalutato il ruolo svolto oggi dall’ecosistema foresta[3] in termini di conservazione della biodiversità e della variabilità genetica. Si intuisce che l’insieme delle attese che la foresta può esplicare sono numerosissime. Da più parti a queste attese sono stati assegnati valori che in parte si collocano in una concezione strumentale ovvero in un valore intrinseco ([15], [61]). In effetti è possibile fare riferimento a beni e servizi, ma anche agli aspetti informativi ed ai valori spirituali ([56]).
Sono evidenti le ricadute che le attese riflettono sulle attività che di fatto possono essere svolte all’interno di aree protette dove comunque dominante se non imperativo, dovrebbe essere l’individuazione della migliore gestione ecosostenibile.
Perseguire una politica di conservazione diretta per entità naturali di forte attrattiva ecologico-naturalistica, scenica e panoramica, significa individuare e perseguire metodi di gestione che di volta in volta possono essere indirizzati verso una salvaguardia estrema (nessun intervento) da contrapporre ad interventi di corretto uso, difesa e ripristino, individuati da modelli operativi organizzati e definiti attraverso parametri scientifici.
Occorre comunque considerare che una gestione ecosostenibile deve valutare in primo luogo i fattori di disturbo e il loro allontanamento, almeno nei limiti del possibile.
Nel caso della deforestazione è emblematico il fatto che la presenza dell’uomo sia condizionata, per la sopravvivenza, alla disponibilità di nuove aree da destinare alla produzione agricola. Da un punto di vista generale la trasformazione nella destinazione d’uso del territorio può essere giustificata di per sé se guardata dall’interno delle popolazioni in pericolo di sopravvivenza e se questa ultima è legata ad un aumento della superficie coltivabile.
È molto difficile in simili situazioni, che appena un secolo fa erano assai meno estese a livello mondiale, convergere, verso gestioni solo rispettose dell’ambiente naturale.
L’estensione e le caratteristiche strutturali di molti (forse di tutti) dei boschi presenti oggi in Italia, derivano dagli effetti dell’uso del passato anche recente, ovvero dalle necessità da parte delle popolazioni le quali, ad esempio, dovevano lottare per riuscire ad individuare il miglior compromesso tra il rispetto dell’unica fonte energetica facilmente accessibile, quella fornita dalla materia prima legno, e la sostituzione del bosco con colture agrarie, che, in condizioni ottimali, erano capaci di fornire rese produttive cerealicole inferiori da 4 a 7 volte quelle odierne[4].
Si è fatto riferimento alla necessità di fare ricorso a modelli operativi applicabili al mondo forestale, basati su dati oggettivi derivanti da indagini scientifiche. In effetti non si dispone di ampie conoscenze sugli ecosistemi forestali anche perché sono poco monitorati, per cui non si può escludere ad esempio, la possibilità di arrecare, anche in tempi brevi, azioni depauperative e di degrado. Il rischio può essere grave in aree ecologicamente difficili dove le azioni di disturbo hanno valenza più elevata.
Del resto le aree protette in generale ed i Parchi in particolare vengono istituiti in diverse realtà ambientali comprendendo un’ampia diversità di risorse vegetali ed animali che sono state modellate dall’uomo con criteri e metodi diversi in relazione alle diverse esigenze e alla differente cultura delle popolazioni interessate. Ad esempio, i beni forestali hanno rappresentato e rappresentano tutt’oggi per le Comunità Montane un fondamentale mezzo economico per la capacità di fornire la materia legno, ma nello stesso tempo rappresentano integrazione territoriale per l’allevamento ed il pascolo nonché per la fauna selvatica.
Si deve sottolineare allora la necessità di caratterizzare in modo più puntuale le funzioni delle aree forestali descrivendo come siano capaci di fornire servizi dei quali occorre quantificare le interazioni anche perché sarebbe utopistico pensare che il bosco possa sempre e comunque esprimere beni congiunti di intensità massima.
Un approccio da seguire è quello che prevede la catalogazione delle risorse a livello di area territoriale e di valutarne i consumi nel tempo considerando nel contempo il fatto della loro presenza in un’ area protetta.
In un recente lavoro Ciancio et al. ([19]) indicano come la definizione di “linee guida per una gestione ecosostenibile delle risorse forestali all’interno dei Parchi Nazionali, derivi dall’interazione fra obiettivi di tutela e caratteristiche degli ecosistemi forestali”.
In altri termini viene posto al centro dell’attenzione l’intervallo ideale compreso tra una gestione finalizzata alla preservazione e quella che prevede comunque l’uso delle risorse ([61]).
Nel caso della preservazione in senso stretto non viene proposto alcun intervento da parte dell’uomo perché è prioritaria la tutela e quindi la necessità di non alterare i processi in atto la cui osservazione ed analisi scientifica può essere momento importante per l’aumento delle conoscenze.
Diversamente viene considerato il concetto di conservazione che si riferisce a valori concreti e che fa riferimento esplicito alle possibilità di uso ed interventi purché venga rispettata la capacità funzionale dell’ecosistema foresta. In questo caso le conoscenze fornite dall’ecosistema sotto un regime di protezione integrale sono fondamentali.
Da un punto di vista gestionale è importante distinguere se la conservazione viene perseguita per il mantenimento della diversità o se è diretta alla massimizzazione di questa. Nel primo caso si adotteranno strategie per il mantenimento di situazioni e varietà di condizioni. Nel secondo caso l’obiettivo è favorire la presenza di tutti gli organismi caratteristici di una determinata area. Un terzo caso potrebbe riguardare sempre, in modo indiretto, il concetto di conservazione: è il caso di situazioni alterate per le quali si prevede il ripristino di uno stato antecedente il disturbo.
Per aree assegnate a forme differenziate di uso è prevedibile la conservazione delle attività tradizionali (agro-silvo-pastorali) in quanto in esse si identifica il riconoscimento di un valore di protezione dovuto alla presenza di emergenze antropologiche, storico-tradizionali e paesaggistiche[5].
Questo pensiero, sviluppato anche in seguito all’importanza ad esso assegnato da Berks et al. ([7]), ipotizza in particolare gli aspetti positivi di tradizioni e usi stabiliti in base ad esperienze di gestioni locali (da parte di utenti) ed ai quali ancora oggi viene riconosciuto un alto valore positivo gestionale.
In generale appare evidente come in situazioni di protezione dominante, l’obiettivo è quello di preservare la più ampia naturalità all’ecosistema forestale ([1]). Questo fatto implica l’accettazione, ma anche la ricerca del migliore compromesso nel riconoscimento odierno (ma anche individuazione) di uno stato di naturalità che è rappresentato da situazioni comunque influenzate dall’attività antropica del passato ([88]), ma che presentano comunque caratteri residuali ancora ad alta valenza quali stabilità, resistenza, resilienza, costanza dei caratteri e della biodiversità ([44], [1], [86]).
L’allontanamento sempre più spinto da stati di naturalità comporta a livello di ecosistema una semplificazione dei processi ed una riduzione della funzionalità, la quale, per mantenere potenzialità produttiva, richiede aumenti di input energetici. Questo avviene nel caso delle produzioni agricole, ma anche in quello di piantagioni da legno (ad esempio, pioppicoltura).
Funzionalità e processi a livelli di sistema
Il punto focale del problema connesso con la gestione sostenibile finalizzata alla conservazione della biodiversità in generale e della diversità genetica in particolare, è che alla base di tutto vi è un allarme mondiale che fa riferimento alla valutazione del danno che sarebbe arrecato al sistema di vita umano odierno per una perdita di diversità biologica. In altri termini alla radice di questa preoccupazione sta la sopravvivenza della popolazione umana che è in forte aumento e che utilizza una produzione primaria fornita da un basso numero di specie coltivate. A questo si associa il problema della conservazione del germoplasma e quello di come individuarne le migliori strategie di gestione.
Riconoscendo a tutte le forme di vita un valore intrinseco, che può essere economico o ecologico, realizzato o potenziale, la gestione della diversità si riflette sulla gestione di tutte le forme di vita e quindi per tutte ne viene sottolineata l’importanza di conservazione ([93], [90], [50]).
Le interazioni tra ambiente e specie a livello di comunità nonché la presenza dell’attività umana, coinvolgono vari aspetti correlati che sollevano considerazioni di natura economica, ecologica, estetica ed etica e che si compenetrano nella protezione delle piante coltivate e di quelle delle specie minacciate nonché nella preservazione di pool genici per l’uso delle generazioni future.
Per il settore degli alberi forestali alta valenza di considerazione è certamente l’aspetto ecologico. Difatti l’estinzione di singole specie o di parte della loro variabilità genetica intraspecifica, che può essere causa di minore sopravvivenza, può comportare la perdita di interi ecosistemi attraverso un effetto a cascata.
L’azione antropica influenza la biodiversità a diversi livelli. In California, lo sviluppo urbanistico incontrollato è ora riportato come la causa principale di pericolo di estinzione di varie specie. Ci sono anche vari esempi relativamente recenti, di estinzione specie-specifica, dovute all’uomo, come il Grizzly della California (Ursus horribilis - [74]).
Poiché l’azione antropica agisce a diversi livelli sugli ecosistemi, è necessario, per le azioni di conservazione, rispondere con azioni diversificate mitigando le cause generali del depauperamento della biodiversità (conservazione degli ecosistemi) e preservando non solo quelle specie sulla soglia dell’estinzione (conservazione monospecifica), ma anche la conservazione della variabilità genetica intraspecifica (conservazione a livello di popolazione). L’attenzione internazionale in questi ultimi anni si è rivolta con particolare interesse alla conservazione ecosistemica sia su scala nazionale che regionale ([5], [74]).
La diversità a livello di ecosistema è dovuta a differenti fattori anche se un’analisi di sintesi ne individua tre di particolare importanza ([73], [39]). Il primo riguarda l’abbondanza di risorse a livello di ecosistema e la concorrenza fra le specie per la fruizione di queste risorse (un ecosistema avrà bassa diversità se poche sono le specie che possono usufruirne). Questo fatto comporta l’esclusione di altre specie in modo anche accentuato se le risorse sono limitate ([14]). Ecosistemi caratterizzati da eterogeneità ambientale consentono una più ampia coabitazione di specie ([34]).
Il secondo riguarda la dinamica intrinseca dell’ecosistema. La comunità ecosistemica è soggetta a disturbi che danno avvio a fasi successionali diversificate che, per la loro presenza, determinano un aumento della diversità e della variabilità ambientale. In questo modo si riducono le probabilità che poche specie possano esprimere una dominanza per lunghi periodi di tempo ([24]).
In questo caso la massima diversità a livello di bioma (distribuzione spaziale di stadi o fasi cronologiche di sviluppo) è quella che si può riscontrare in un soprassuolo forestale che, sull’unità di superficie, abbia subito il più alto numero di disturbi in quanto su quella superficie ci si può attendere la presenza ripetuta di più fasi cronologiche.
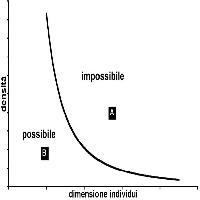
Se i disturbi sono piccoli, come ad esempio la morte e/o la caduta di una o poche piante, nonché ripetuti nel tempo anche in successione frequente, una grande diversità può essere raggiunta anche su di un ettaro (Fig. 1). Se il disturbo è esteso e massiccio (valanga, tromba d’aria, eruzione vulcanica) con bassa frequenza temporale gli stessi valori di diversità potranno ricercarsi su superfici molto più vaste ([80]).
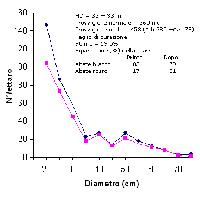
Fig. 1 - Bosco Giovald (Lavarone, TN). Diversità della distribuzione dei diametri (bosco misto di Abete bianco e Abete rosso) determinata dai disturbi del taglio di curazione (prima e dopo il taglio).
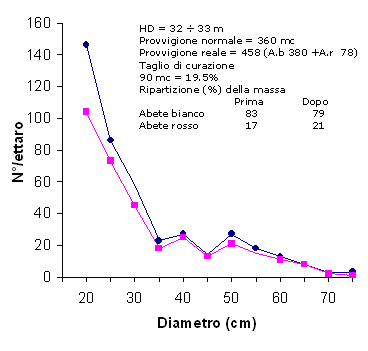
Un terzo caso coinvolge il concetto di dominanza specifica. L’abilità di alcune specie di dominare può essere regolata attraverso l’azione di predatori, parassiti, patogeni ([64], [42]).
L’azione congiunta di questi fattori può spiegare come la diversità risulti influenzata dagli effetti dell’interazione tra l’ambiente e le specie. Ma per addivenire alla comprensione più profonda del perché una foresta è più diversa di un’altra, è necessario considerare fattori che agiscono a livello storico e geografico regionale e che incidono sulla distribuzione delle specie ed all’azione dei fattori evolutivi su di queste.
Più in generale la diversità dell’ecosistema naturale foresta si rileva come un mosaico di microambienti nel quale possono vivere e coesistere, anche in stretta vicinanza, specie con autoecologia anche solo leggermente diversa rispettando il principio dell’esclusione competitiva. Tutto ciò crea ricchezza e diversità di aree. Uguale valenza ecologica crea competizione reale nell’ambito della stessa nicchia ecologica. La presenza di una specie è presumibilmente spiegabile perché particolari interazioni tra caratteristiche genotipiche e condizioni ambientali (o casualità ) hanno favorito le progenie di quella specie rispetto alle concorrenti (alla specie favorita può essere riconosciuto un maggior valore di fitness) [6].
La diversitàè però, come già indicato, originata e controllata anche dalla frequenza dei fenomeni di disturbo che in relazione alla loro periodicità ed intensità possono alterare la funzione competitiva e quindi l’organizzazione della struttura e della dinamica degli ecosistemi.
Nei boschi naturali le aperture (cadute di alberi) consentono l’introduzione e l’affermarsi della rinnovazione naturale. Il selvicoltore aveva percepito questa dinamica. Desiderando il corretto uso e la perpetuità del bosco attraverso la rinnovazione, codificava il trattamento, dedicando attenzione a singole piante o a gruppi di queste la cui utilizzazione, ad intervalli di tempo diversificati, doveva soddisfare in modo positivo la preoccupazione di mantenere e usare in modo corretto la potenzialità produttiva della stazione, compresa quella (in prevalenza erbacea) da destinarsi alla componente animale di un’attività pascoliva e zootecnica altrettanto essenziale per la sopravvivenza delle comunità umane.
Le foreste sono caratterizzate da cicli molto lunghi per cui un osservatore poco attento potrebbe in una primo momento, considerarle comunità statiche. Il dinamismo è invero assai intenso e descrivibile in fasi di successione le quali possono essere caratterizzate da ricchezza di specie e diversità differenziate ([21], [62], [11], [40]).
Per comprendere i meccanismi che determinano la diversità di specie, il primo step è chiarire se esistono delle regole che determinano o influenzano l’assemblaggio delle specie. Una delle domande che deve avere ancora una risposta è quale fattore influisce sull’assemblaggio delle specie: sono i (i) fattori deterministici, così come la separazione in nicchie lungo un gradiente ambientale, o i (ii) fattori stocastici così come i processi di colonizzazione?
I materiali, i siti ed il target di scala variano secondo le situazioni. Quindi occorre un gran numero di informazioni sulle diverse tipologie[7].
Non si dispone di dati oggettivi, statisticamente validati, per dare una risposta puntuale alle domande poste. Può essere di aiuto comunque fare alcune considerazioni di carattere generale in quanto efficaci a trovare la via che potrebbe fornire un alto contributo all’individuazione delle strategie da adottare, a livello di ecosistema. Occorre sottolineare che, in considerazione delle complessità del problema, soprattutto nel caso di scelte gestionali, è importante possedere le basi scientifiche per una continua riflessione sui processi che la natura mette in opera nei suoi vari scenari.
Nella maggior parte degli ecosistemi studiati è stata dimostrata una relazione positiva tra diversità e produttività ([86]), mentre pochi sono gli ecosistemi in cui è stata trovata una relazione non significativa; invero sono noti casi in cui aumentando la diversità di specie diminuisce la produttività ([38]). Questi risultati, non sempre concordanti, hanno fatto ritenere che molto probabilmente la relazione positiva deve essere vista come dipendente da una relazione positiva tra diversità ed abilità competitiva ([17]) delle specie. Se una specie ha l’abilità di massimizzare il guadagno di carbonio in un particolare ambiente questa capacità o superiorità le conferirà anche un vantaggio competitivo in quel dato ambiente. Di conseguenza la competizione renderà massima la produttività favorendo la specie più produttiva[8].
Nelle foreste le specie successionali pioniere a crescita veloce sono rimpiazzate dalle specie successionali definitive caratterizzate da uno sviluppo più lento. In questo caso la produttività non è massimizzata dalla competizione poiché i boschi dominati da specie successionali definitive a crescita lenta, fissano meno carbonio dei boschi dominati da specie successionali pioniere a crescita più rapida. D’altra parte la produttività non aumenta con la diversità successionale: soprassuoli forestali misti di specie successionali precoci e tardive possono fissare meno carbonio di boschi dominati da una singola specie pioniera a crescita veloce.
Le foreste sono state scarsamente menzionate nel dibattito della diversità a causa della difficoltà di condurre esperimenti in ecosistemi con una dinamica lenta. I dati disponibili possono essere utilizzati comunque per esaminare se l’ecosistema foresta differisce dagli altri ecosistemi ([17]).
La maggior parte della ricerca sulla biodiversità si è concentrata sulla perdita della diversità vegetale e sugli impatti che si ripercuotono sui processi ecosistemici ([60], [86], [89]). Non c’è nessuna ragione di credere che una minore biodiversità di consumatori e decompositori avrà un impatto minore ([43], [57]). Le comunità microbiche del suolo sono tra i componenti che presentano la maggior ricchezza in specie dell’ecosistema terrestre ([92]). La maggior parte del flusso annuale di carbonio e nutrienti avviene nei primi 10 cm del profilo del suolo ([91]). La dimensione e la natura chimica di questi flussi sono controllati dagli organismi del suolo, per cui questi sono componenti integranti di un grande range di processi ecosistemici che includono i cicli dei nutrienti, lo stoccaggio del carbonio, la degradazione di inquinanti, il mantenimento della struttura.
Una prima conclusione per gli obiettivi di questo lavoro appare evidente e riguarda alcune caratteristiche degli ecosistemi forestali. Nelle scelte per una gestione eco-sostenibile risulta prioritario:
- salvaguardare la complessità funzionale dell’ecosistema;
- considerare che, essendo nell’ambito forestale i cicli temporali molto lunghi (l’unità di misura potrebbe essere il secolo), la valutazione dell’azione di interventi anche di ripristino, può essere giudicata dopo diversi anni.
A favore di una gestione che ponga la diversità come punto di riferimento centrale, sta la motivazione che vede coinvolta la stabilità. In effetti il principio di Elton ([26]) che indica come gli ecosistemi ad alta diversità siano più stabili rispetto a quelli meno diversi non è accettato da tutti gli ecologi o per lo meno alcuni di questi fanno notare l’assenza di un’evidenza generale e prevedibile per la relazione diversità /stabilità ([54], [46]) anche se un sistema stabile è probabilmente più diverso ([33]). Una relazione positiva diretta e di lunga durata temporale tra diversità e produttivitàè stata trovata di recente studiando l’ecosistema marino del reef (barriera corallina). In questi casi l’analisi ha evidenziato come la stabilità ecologica sia in relazione con la diversità tassonomica ([45]). Secondo l’ecologo Pimm ([67]) la natura della stabilitàè comunque individuabile attraverso un’analisi tra resistenza e resilienza.
Gli aspetti delle relazioni intercorrenti tra perdita di diversità e funzionalità devono essere valutati attraverso un approccio che inquadri le relazioni tra disturbo e risposta a questo da parte dell’ecosistema. Questo può esprimere la resistenza nei confronti di cambiamenti che incidono sulla propria funzionalità ovvero possono possedere una minore o maggiore valenza di risposta (resilienza al fattore perturbatore) per tornare ai livelli funzionali precedenti.
È stato già accennato che se si considera l’ipotesi di un effetto positivo della biodiversità, la resistenza e la resilienza dovrebbero aumentare (a livello di sistema) con la ricchezza in specie in quanto un maggiore numero di queste può permettere un più elevato range di risposte alle perturbazioni. Tra l’altro, una maggiore ricchezza aumenta la probabilità che alcune specie a basso tasso di performance (è il caso delle specie altamente specializzate) aumentino la propria presenza essendo compensate da altre. Le risposte di un ecosistema ad una perturbazione dipendono comunque dalla forma e dalle relazioni esistenti tra variabilità della risposta e biodiversità.
Per alcuni ecosistemi naturali (ad es., le praterie) la produzione di biomassa è di fatto un’importante risposta variabile e associata con la ricchezza di specie vegetali; non si può escludere la possibilità di pervenire a conclusioni simili anche per le foreste per le quali sarà necessario considerare il completo ciclo successionale delle fasi di sviluppo e le loro durate temporali.
Per queste ultime è stato evidenziato sperimentalmente che in condizioni prive di stress ambientali, le specie caratterizzate da bassi valori del sistema resistenza/resilienza hanno capacità di produrre livelli minori di biomassa rispetto a quelle caratterizzate da un sistema più efficiente. Le prime saranno più resistenti alle perturbazioni rispetto alle seconde ([66]).
Più in generale la biodiversità favorisce la produzione di biomassa, ma nello stesso tempo differenti associazioni diversità /produttività possono portare al verificarsi di relazioni di segno opposto tra biodiversità e stabilità di un ecosistema. Seguendo questo pensiero ci dobbiamo attendere situazioni di elevata diversità, nelle quali perturbazioni di una certa intensità possono essere non sufficientemente efficaci da rimuovere la dipendenza della funzionalità dell’ecosistema dall’effetto positivo della diversità per cui è evidente che vi siano molti argomenti a favore di mantenere un’elevata biodiversità per ottenere un’altrettanta elevata produttività, anche se sistemi con poche specie sono più resistenti e/o resilienti. D’altra parte in sistemi in cui la nicchia di complementarità non rappresenta motivo di dipendenza dalla diversità, potranno verificarsi situazioni perturbative multiple (siccità, incendi, utilizzazioni) tali che la produttività risulterà influenzata in modo più efficace dagli eventuali alti valori dei livelli trofici piuttosto che da elevati tassi di diversità.
Più in generale si può aumentare uno scambio potenziale tra produzione e stabilità in sistemi dove la produzione è dipendente dalla diversità. La valutazione di questo scambio può risultare utile per comprendere le conseguenze, a livello di ecosistema, dei cambiamenti di biodiversità ([66]).
Recenti lavori dimostrano che la produzione primaria entro singoli livelli trofici, spesso, ma non sempre, è saturata attraverso la ricchezza di specie ([25]). Tuttavia si deve considerare che la variazione nella diversitàè necessariamente associata con i cambiamenti della configurazione in specie (identity) e che i cambiamenti nella biodiversità spesso avvengono attraverso livelli trofici multipli[9].
Old-forest growth: modello di riferimento di naturalità
L’imperativo che la diversità genetica soprassieda alla funzionalità e alla potenzialità evoluzionistica è valido nel caso di un coinvolgimento della sostenibilità dell’efficienza biologica delle foreste, ma anche in un contesto di cambiamenti ambientali.
Queste considerazioni inducono a ritenere, in un primo momento, che la conservazione della diversità genetica degli alberi, che rappresentano negli ecosistemi forestali la componente dominante di forme di vita, rivesta uno speciale significato. In un secondo momento è necessario prendere in esame, seguendo un approccio scientifico, strategie informative che siano capaci di fornire le conoscenze di base per una comprensione delle interazioni intercorrenti tra funzionalità dell’ecosistema e diversità genetica. Ovvero è necessario individuare dove sia possibile reperire le informazioni di riferimento per poi saggiare nelle foreste coltivate[10] la validità dei modelli colturali e/o gestionali ad esse applicati.
Dalla letteratura ed in particolare da quella sviluppatasi nelle Scuole di ecologia forestale degli Stati Uniti e del Canada, emerge che, punto di riferimento conoscitivo in questa specifica tematica è espresso dalle così dette Old-growth forests in quanto considerate gli ecosistemi forestali caratterizzati dal più alto valore conservativo di specie ([2]). Infatti queste foreste possono fornire la più alta variabilità di habitat capaci di sostenere le più ampie catene trofiche di vita naturale.
Ad esse è riconosciuto sia un alto valore biologico che un valore sociale. Questo ultimo perché è assegnato un valore economico alla presenza di piante che si distinguono per le grandi dimensioni (diametro, altezza), ma soprattutto perché suscitano forte attrattiva, come parte di un retaggio naturale del passato o dell’interpretazione di questi. La letteratura specialistica indica diversi criteri che sono stati considerati per assegnare una definizione delle Old-growth forests[11]. È condivisibile quanto indicato da Hunter ([9]) che supera le diversità di pensiero individuando le Old-growth forests in quei soprassuoli forestali in cui la composizione specifica si è stabilizzata, l’incremento medio annuo di massa legnosa è prossimo allo zero (biostasi), l’età media è significativamente superiore al tempo medio intercorrente tra la fase di disturbo/innovazione e la fase di degradazione, gli alberi del piano dominante hanno un’età equivalente a quella della vita media attesa per una specie in una particolare stazione, l’azione di utilizzo è quasi assente o molto limitata, nessuna azione è intervenuta circa eventuali cambiamenti di e da altri tipi di ecosistemi.
È evidente che al parametro età dell’albero o del soprassuolo, viene dato un valore dominante anche se definire quando, un albero o una comunità di alberi, siano biologicamente maturi non è certamente facile. La maturità biologica è comunque più vecchia di un “climax economico”.
Ma il ruolo svolto da questo tipo di foreste è anche di riserva di diversità genetica e di adattamento riproduttivo. Alcuni studi recenti ([59]) condotti in Canada in boschi di abete rosso americano (Picea rubens) hanno evidenziato relazioni statisticamente positive tra altezza degli alberi, carattere valutato come espressione della fitness, e la percentuale di loci polimorfici, mentre relazioni negative appaiono qualora venga considerato il potenziale genetico latente ([6]) quale misura della ricchezza genetica espressa dalla bassa frequenza di alleli rari. La superiorità delle piante in fase di biostasi avanzata emerge anche nel caso delle performance delle generazioni parentali (boschi da seme di qualità intrinseche superiori). Difatti in generale le popolazioni di età più avanzata mostravano caratteristiche specifiche di natura genetica di mantenere o di promuovere un potenziale genetico superiore nelle progenie in termini di sviluppo e crescita in altezza. Tali condizioni erano esclusive di boschi estesi in superficie e caratterizzati da valori di densità sufficienti ad impedire effetti negativi di consanguineità e deriva genetica.
In Italia situazioni di riferimento così descritte praticamente oggi non esistono. Ciò a causa dei rapporti che da oltre 5000 anni si sono instaurati tra risorse del territorio e attività antropica. La necessità di disporre di una produzione agricola sempre maggiore ha determinato la contrazione della superficie a bosco che pur essendo sorgente principale di fonte di energia veniva sacrificato a vantaggio delle produzioni alimentari. Alcuni esempi di Old-growth forest sono stati descritti: a) per la lecceta del Supramonte di Orgosolo (NU), località Mereu e Nuraghe Mereu, dove fasi disetanee adulte-giovani si compenetrano a fasi senili di sviluppo caratterizzate da piante in età molto avanzata e grossi diametri ([80]); b) per le faggete del Vallone del Venaquaro (Gran Sasso) in provincia di Teramo dove in un’area in contestazione tra i Comuni di Pietracamela e di Fano Adriano, le utilizzazioni forestali sono state sospese da oltre 400 anni. Sono dominanti le fasi adulta e senile con piante di 300-350 anni e diametri di oltre 100 cm (Giannini et al., dati non pubblicati).
Nel primo caso ([80]), viene indicato come l’aspetto della foresta fosse caratterizzato dall’accumulo al suolo di notevoli quantità di necromassa, per lo più fusti e rami di grandi dimensioni in diversi stadi di decomposizione. La struttura somatica di complesso del popolamento arboreo (disetanea), l’esistenza di alberi con diametri fino al oltre 150 cm e con età superiori a 500 anni (e in qualche caso fino a quasi 300 cm con più di 1000 anni d’età ), la tipicità di decorso degli incrementi legnosi erano altri non meno importanti attributi comuni al soprassuolo, i quali, essendo col primo propri delle foreste vergini, facevano conferire a più tratti della lecceta del Supramonte caratteri di naturalità. Nel soprassuolo arboreo erano inoltre individuabili le fasi morfologico-strutturali che contrassegnano le tappe evolutive dei popolamenti forestali ed equilibrio naturale. In particolare venivano ad essere riconosciute distribuzioni aggregate di alberi isodiametrici appartenenti allo stesso stadio cronologico in numero ed ampiezza tale da conferire al popolamento una struttura articolata la cui norma veniva espressa da una funzione di tipo esponenziale. Riguardo alle funzionalità dell’ecosistema nello studio tale norma viene indicata come modello di stato di equilibrio.
Le ricadute pratiche di tali osservazioni sono importanti perché una gestione di tipo conservazionistico può essere correttamente impostata secondo criteri naturalistici avendo come modello le strutture descritte.
Per meglio comprendere cosa significhi avere come modello di riferimento le Old-growth forests, occorre accettare ancora una volta il ruolo svolto dal fattore tempo nel controllare la dinamica dei singoli alberi e dell’intero ecosistema foresta. Gli alberi più vecchi, secondo la specie, possono raggiungere e superare anche i 1000 anni, per cui potranno essere diversi o più di uno i cicli successionali a cui occorre fare riferimento ([22], [58]). La prima implicazione che ne deriva è che difficilmente in aree soggette ad una gestione di conservazione, potrà essere accettata l’applicazione di una selvicoltura intensiva basata su ecosistemi fortemente antropizzati, semplificati ed estesi su vaste superfici ed in particolare assoggettati a cicli molto brevi.
Conservazione e selvicoltura eco-compatibile
Le foreste rappresentano una delle componenti di maggior importanza tra gli ecosistemi terrestri. Sotto l’aspetto funzionale sono ricche di caratteristiche proprie tra cui elevata complessità. Merita ricordare che anche nei confronti di una conservazione ambientale, i boschi hanno svolto e svolgono ruolo determinante nel rappresentare oggetto di riferimento[12].
Anche se la pressione antropica delle attività forestali è stata ed in parte lo è ancora, assai alta (ciò ha portato sicuramente ad una manomissione più o meno accentuata della naturalità ), è doveroso riconoscere che è sempre più diffusa la convinzione che vi siano dei limiti che non possano essere superati anche nell’uso delle foreste ([55]).
Lo sviluppo di tale pensiero non è certamente di oggi: il riconoscimento al bosco di risorsa rinnovabile, caratterizzata da limiti di sopravvivenza, nasce alla fine del diciannovesimo secolo quando prende origine e si sviluppa il concetto di selvicoltura naturalistica (close to nature silviculture) in un momento in cui erano emergenti nuovi paradigmi sotto l’influenza dei Fisiocratici ed il loro motto “Return to nature” ([76], [4], [3]).
Gli insegnamenti di Gayer ([30], [29]), ripresi successivamente dalle Scuole forestali di Svizzera, Francia ed Italia, rappresentavano riferimento dottrinale per una selvicoltura diretta alla realizzazione di soprassuoli forestali misti e caratterizzati da irregolarità strutturale.
La filosofia profusa dalla Scuola forestale di Monaco individuava una metodologia da applicare alla pratica selvicolturale, quindi di coltivazione, e non poteva avere ricadute diverse essendosi basata e sviluppata in seguito ad esperienze di utilizzazione del bosco.
In Italia l’inizio operativo di una Selvicoltura, dalla locuzione collettiva denominata naturalistica, si può datare al 1951, anno in cui entrò in vigore il primo di una ventina di piani di assestamento che in un decennio sottoposero a un inedito regime, la maggior parte dei boschi del Cadore orientale e di alcuni del Cadore occidentale ([78]).
Da allora l’idea di una selvicoltura diversa da quella in auge, ma comunque vicina a quella praticata dalla Serenissima Repubblica di Venezia dal 1500 al 1700, non è stata più abbandonata (anche se la Repubblica focalizzava al massimo la garanzia di un riferimento continuo nel tempo di “assortimenti strategici”. È anzi via via penetrata estendendosi, pur con varietà di interpretazioni, fin alla sua consacrazione avvenuta nel 1980 con l’emanazione di una normativa regionale ([78]).
In sintesi venivano dettate alcune prescrizioni, convalidate nel tempo che prevedevano:
- mantenere, normalizzare e ripristinare lo stato disetaneo con tagli saltuari in bosco misto, con tagli “da dirado a gruppi” o marginali in bosco puro, limitare al minimo l’ampiezza delle tagliate; favorire il faggio ed allevarlo ad altofusto;
- curare attentamente il governo della densità (copertura, rinnovazione) attenendosi alle indicazioni dei piani, ma, contravvenendo ai sacri canoni, con facoltà di rilasciare gli alberi più grossi e vigorosi (fino a 60-65 centimetri) privilegiando così la scelta qualitativa (età, vitalità ) poiché la martellata doveva essere colturale, cioè badare innanzitutto alle esigenze del bosco.
Successivamente la dottrina ecologica veniva applicata a diversi tipi di bosco e concorreva a svincolare la selvicoltura dagli empirismi dei tagli mercantili. Il fattore humus-suolo acquistò allora e per la prima volta un ruolo funzionale integrativo del soprassuolo. In esso anche i consumatori, ad esempio ungulati, assumevano un ruolo parimenti funzionale come regolatori, come feedback, della vegetazione e della rinnovazione. Si chiariva e si rafforzava così il significato della complessità e della disetaneità di struttura del soprassuolo nonché della imprescindibilità della sua composizione polifita. In sede di trattamento si doveva, fra l’altro compiere la scelta degli alberi da eliminare “molto più guardando in basso... che solamente guardando in alto”, facendosi guidare dalla vegetazione del sottobosco per giudicare le condizioni di suolo circa la fertilità e le attitudini alla rinnovazione ([78]).
Nel modello scompariva il classico diametro di recidibilità come rigido limite superiore per l’ammissione invece di alberi fino a 75 cm ed oltre. Compariva il parametro della statura del bosco, definita dall’altezza media degli alberi dominanti, quale espressione della fertilità della stazione, ma anche dello spazio epigeo ed ipogeo protetto nel quale il sistema viveva compiendovi tutti i processi biologici dalla genesi dello humus e dalla nascita delle plantule al loro accrescimento fino alla caduta per decrepitezza. La foresta provvista di tali caratteri - mescolanza, più elevata consistenza, struttura stratificata - appariva in grado di assicurare meglio la propria continuità nel tempo mediante la costanza della rinnovazione e la libertà rispettata di competizione ed evoluzione dei suoi componenti (compresi il dominio alterno nel tempo delle specie edificatrici), mantenendo un incremento sostenuto, ma prodotto da masse relativamente elevate con minori saggi di volume, come è nell’ordine naturale delle cose (Tab. 1 - [78]).
Tab. 1 - Caratterizzazione delle strutture disetanee del Comelico. In prevalenza boschi misti di abete rosso, abete bianco e faggio. Dati riguardanti una superficie totale di circa 12000 ha e circa 800 particelle (da [78], modificata). Per maggiori dettagli, vedi Appendice 1.
| Caratteri del soprassuolo | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Anni | S. | Alb. | area b. | massa | increm. | diam. | Composizione | ||
| m. | n ha-1 | m2 | m3 | m3 ha-1 | cm. | Picea | Abies | altri | |
| 1950-51 | 27 | 300 | 18 | 220 | 3.6 | 50-55 | 75 | 23 | 2 |
| 1996 | 29 | 290 (350) | 33 | 290 | 5 | 70-80 | 77 | 17 | 5 |
| differenza | 2 | -10 | 15 | 70 | +1.4 | 5 | 2 | -6 | 3 |
| Norma | |||||||||
| S = 29 | 320 | 28 | 280 | 6 | 75 | 40 | 40 | 20 | - |
| Caratteri auxometrici | |||||||||
| Anni | Incremento | Ripresa unitaria m3 ha-1 | |||||||
| corrente (m3 ha-1) | percent.% | saggio utilizz. % | ogni 9 anni 8-16 | Media annua | |||||
| 1950-51 | 3.6 | 1.7 | 4.0 8.0 | 6.0% di 225 m3 ha-1 | 1.34 37% di 3.6 m3 | ||||
| 1996 | 5.0 | 1.7 | 6.6 | 6.6% di 290 m3 ha-1 | 2.14 43% di 5.0 m3 | ||||
| differenza | +1.4 | 0 | -0.6 | 7 | +1.2 | ||||
Schütz (1999) riporta come l’applicazione[13] per oltre un secolo in Svizzera del pensiero di Gayer, che il selvicoltore concretizzava nel taglio saltuario, abbia consentito di verificare la concretezza di molte delle idee da questi espresse congiuntamente ad altre disattese perché difficili da realizzare. Importante è il fatto che per queste ultime siano stati guardati con senso critico i punti più difficili. D’altra parte il fine ultimo operativo era quello che prevedeva di raggiungere soluzioni di gestione attraverso un affinamento del sistema anche se ciò richiedeva comunque tempi lunghi a causa della lentezza dei processi a livello di ecosistema foresta. In effetti è molto difficile, nella pratica, realizzare sempre ed ovunque quanto indicato dalla teoria anche se maturata in base ed esperienze di lavoro. Un primo fattore di disturbo ad esempio, è quello relativo alla variabilità stazionale che può coinvolgere la singola pianta o gruppi di queste.
Per valutare l’efficienza funzionale di questo modello selvicolturale è necessario considerare il bosco come edificato di unità o microcosmi eco-gestionali (eco-unità ) distribuiti in modo non ordinato nello spazio e nel tempo nell’ambito di singoli complessi boscati. In questo caso il selvicoltore opera in armonia a tale filosofia attraverso interventi coscienti tesi al mantenimento della struttura presente perché ritenuta concorde con quella ideale, ovvero operando in modo diversificato per una sua realizzazione.
In questo contesto si deve accettare il fatto che mantenere e/o realizzare una ricchezza (mescolanza) di specie possa richiedere interventi anche poco rispettosi della naturalità dei processi successionali, ma, nello stesso tempo, questo potrebbe essere richiesto per preservare le specie meno competitive.
Le specie arboree presentano differenti stadi di sviluppo[14] che si organizzano in ecostrutture (eco-unità : superficie sulla quale una parte di foresta si insedia e inizia a vivere secondo una propria dinamica - [35], [63]), che possono avere esigenze autoecologiche proprie e durata temporale diversificata dipendendo queste dalle caratteristiche intrinseche delle singole specie e dalle interrelazioni che si instaurano con le condizioni ecologico-stazionali del bosco ([79]). La fase di maturità o biostasi della biocenosi ([63], [28]) che in prima approssimazione, è espressa a partire dal momento della culminazione dell’incremento medio annuo in altezza degli alberi, riguarda spesso un periodo temporale molto lungo anche perché i singoli alberi possono raggiungere in momenti diversi il profilo più alto delle chiome occupando lo spazio disponibile e realizzando una stratificazione compatta, relativamente uniforme e duratura nel tempo[15].
In natura le varie fasi successionali possono di per sé essere costituite e realizzarsi attraverso anche una mescolanza di specie. Invero nelle foreste dei climi temperati le fasi di sviluppo finali del ciclo successionale tendono ad essere dominate da una singola specie. Si fa riferimento, come esempio alle fasi climax di maturità dei boschi di leccio, di faggio, ma anche di abete rosso, nelle quali è presente una uniformità diffusa nella specie e nella struttura soprattutto se riferita alla posizione spaziale delle chiome. In questi casi molte specie tendono a scomparire a causa di una loro scarsa competitività intrinseca e/o per insufficienza di radiazione luminosa nei piani inferiori. Ma allo stesso tempo non mancano esempi in cui la fase di maturitàè dominata da mescolanza quali Abete bianco - Faggio, Abete rosso - Abete bianco - Faggio, Rovere - Farnia.
In generale la più alta ricchezza specifica è tipica della fase così detta rigenerativa (dalla fase di inizio della decrepitezza e senescenza alla fase affermativa di innovazione) che quasi sempre interessa periodi temporali più brevi della precedente.
Le osservazioni di Korpel ([48], [49]) in boschi di abete rosso, sui Monti Tatra in Slovacchia indicano come questa specie, caratterizzata da una elevatissima valenza autoecologica, realizzi in condizioni stazionali ottimali, soprassuoli densi, chiusi e con struttura molto regolare se si considera la posizione della chioma nello spazio piuttosto che la distribuzione dei diametri dei fusti a petto d’uomo o dell’età dei singoli alberi[16]. A quota più elevata e quindi in condizioni ambientali di maggiore stress, i soprassuoli di foreste vergini della stessa specie si presentano con struttura irregolare e più “aperta” che consente anche la presenza di altre specie arboree.
Le osservazioni nelle foreste naturali sono molto istruttive, anche se non risolvono certamente in modo esauriente l’esigenza di perseguire gli obiettivi di diversità genetica e irregolarità strutturale delle componenti dominanti dell’ecosistema foresta. In situazioni naturali acquistano, però importanza perché recano sicuramente un aumento delle conoscenze sulla dinamicità temporale del popolamento forestale e rappresentano punto di riferimento per una valutazione di applicabilità di modelli colturali rispettosi di una selvicoltura naturalistica. In altri termini la disponibilità di dati scientifici di riferimento consente di individuare ed applicare interventi di affinamento e/o di correzione finalizzati ad arrecare un effettivo beneficio biologico all’ecosistema bosco e raggiungere il migliore compromesso tra uso eco-compatibile e condizioni di massima naturalità [17].
Si deve sottolineare che comunque l’indicazione di una regola di carattere generale finalizzata ad esempio al raggiungimento di irregolarità del soprassuolo, è accettabile anche nei casi in cui si debba operare per un ripristino di situazioni alterate, ma non appare estensibile ovunque ed in tutte le situazioni.
L’idea di una differenziazione strutturale a livello microstazionale ed incentrata sulla posizione del singolo albero ha inizio in Svizzera per opera di Biolley ([8]) e trova identità nel taglio saltuario (single-tree selection system).
Tanto nelle strutture disetanee naturali, quanto in quelle di soprassuoli coltivati la struttura di equilibrio è caratterizzata da rapporti degli alberi adulti (da 15 - 20 cm di diametro in su) che decrescono dalla fase giovane a quella senile.
Nelle foreste coltivate o più generalmente antropizzate le aree occupate dalle singole fasi sono di estensione variabile, ma comunque ridotta, intercalate e compenetrate fra loro; già su piccole superficie (1 - 2 ha) è possibile trovarle rappresentate tutte. Sul terreno, in senso orizzontale, la struttura di complesso che ne deriva è generalmente aggregata, con distribuzione delle aggregazioni (gruppi) di tipo casuale ([86]); mentre in senso verticale le aggregazioni di età, diametri ed altezze differenti si stratificano scalarmente su piani diversi, tendendo a saturare il biospazio epigeo.
Sul terreno un’aggregazione per alberi isodiametrici non appare rilevabile o lo è solo in modo incerto e frammentario. Applicando invece il metro dell’età al posto di quello del diametro, se si formano classi cronologiche di una certa ampiezza (20-30 anni) e se si considera come aggregazione elementare (minima) la serie unitaria di alberi che può condurre in media ad un albero “maturo” (per esempio 8 alberi di 30 anni, 6 di 50 anni, ecc., fino a 1 albero di 150 anni), la struttura di complesso risulta da un insieme di popolamenti elementari in forma di gruppi di piccola estensione, costituiti da alberi della stessa classe di età e da alberi isolati di età differenti, che vivono, dominanti o dominati, all’interno del gruppo e fra gruppo e gruppo. In altri termini nelle strutture disetanee gli alberi si distribuiscono quindi non in aggregazioni isodiametriche, bensì in aggregazioni di individui tendenzialmente coetanee ([81], [82], [83]) [18].
In realtà il concetto di struttura disetanea non è di immediata comprensione, né sempre di facile individuazione sul terreno. Nulla vieta di concepire tra la compresa coetanea teorica ed il teorico soprassuolo disetaneo “per piede d’albero”, una successione di forme strutturali intermedie, quali nella realtà, in termini estremamente variabili, è dato di regola di riscontrare. La maggiore o minore distanza dai due estremi teorici sembra potersi essenzialmente correlare alla estensione delle aggregazioni che costituiscono il “complesso”. C’è tuttavia un’estensione limite delle aggregazioni al di là della quale la struttura coetanea di un complesso trapassa in quella disetanea e viceversa. Un limite evidentemente non troppo rigido, né immutabile, poiché lo si può supporre funzione simultanea del clima regionale, dell’ecologia della stazione, del temperamento degli alberi che formano il complesso e di molti altri fattori ([81]).
Nelle radure e nelle lacune più vaste che si producono in seguito alla caduta degli alberi decrepiti (fase di rinnovazione), si instaurano condizioni ecologiche simili a quelle delle tagliate a raso di ampie dimensioni. Il popolamento arboreo che vi risorge assume una struttura coetanea o paracoetanea, con densità che permangono elevate a tutte le età. Radure e lacune di ampiezza inferiore a 1000 m2, ma più sicuramente fra 200 e 600 m2, conservano invece condizioni di bioclima e di suolo meno diverse rispetto a quelle del bosco chiuso. Con novelleti già in partenza meno densi che qui si insediano, l’evoluzione segue tendenze diverse determinando tuttavia una più marcata riduzione numerica e ad una più intensa selezione, promossa dal “contorno” (competizione, protezione laterale, illuminazione diretta discontinua, maggiore umidità, microclima marginale, ecc.), ovvero dal competitivo ambiente in cui il novelleto deve svilupparsi. Tale processo è tanto più attivo quanto minori e più frazionate sono nelle strutture disetanee le are del novelleto. La più forte concorrenza permette agli individui meglio dotati di emergere precocemente e di dominare gli altri circostanti, raggiungendo dimensioni che non si osservano nelle ordinarie strutture coetanee, salvo in quelle fortemente diradate.
Con senso critico anche Schütz ([76]) indica questo modello selvicolturale, anche se maturato nel tempo da esperienze e pratiche locali e quindi influenzato da fattori socio-economici[19], molto vicino alla filosofia della selvicoltura naturalistica. Il paradosso sta forse nel fatto che lo stesso Autore commenti come, per un’applicazione corretta nello stesso vi sia la necessità di un intervento del selvicoltore puntuale e continuo. Difatti ad esempio, per i boschi misti del consorzio Picea, Abies, Fagus Susmel ([85], [77], [84], [82]) indica, sotto l’aspetto teorico, parametri di riferimento relazionati alla fertilità della stazione che impongono, per il mantenimento di strutture fortemente articolate e per l’affermazione della rinnovazione naturale, di non superare certi valori limiti di biomassa legnosa ad ettaro. Questo fatto determina tra l’altro l’intervallo temporale tra un intervento di utilizzazione e l’altro (periodo di curazione).
I limiti più evidenti riguardano il fatto che il modello consente una reale applicabilità solo in alcuni casi tra i quali quelli in cui sia presente irregolarità permanente della struttura e/o mescolanza di alcune specie. Devono altresì essere soddisfatte alcune condizioni. Queste riguardano in primo luogo l’assenza di fattori che possano condizionare l’insediamento e lo sviluppo della rinnovazione (frequenza di annate di fruttificazione abbondante, componente faunistica in equilibrio, etc.). Inoltre difficoltà possono emergere in presenza di specie ad elevata esigenza di radiazione luminosa (ad es., molte specie del genere Pinus), ma anche nel caso di quelle caratterizzate da fasi successionali che si protraggono per periodi di tempo molto lunghi e che tendono a regolarità strutturale. Una estensione dei limiti di applicabilità porterebbe al ricorso di interventi ripetitivi, intensivi ed ad alta artificialità.
Il sistema colturale saltuario per piede d’albero confluisce e si associa ai modelli che incentrano l’attenzione per la sostituzione del soprassuolo maturo, su interventi in prevalenza per gruppi di varia estensione e forma che prendono avvio spesso dai margini del soprassuolo[20] stesso (irregular shelterwood system - [75], [51], [23], [9]).
Tutti questi modelli basandosi sull’analisi della dinamica successionale dell’ecosistema foresta, rientrano a pieno titolo in una visione di selvicoltura naturalistica. In altri termini la selvicoltura in tutti questi casi prevede modelli di un bosco colturale che perseguono la rinnovazione naturale attraverso il controllo di una maggiore o minore disponibilità di intensità di radiazione luminosa fatta pervenire al suolo attraverso una modifica cosciente della diversità nello spazio della chioma degli alberi. In questo caso l’aggettivo naturalistica, che accompagna il sostantivo selvicoltura,sta ad indicare ed a comprendere il riferimento a tutti i modi di realizzazione della rinnovazione naturale ([76]).
Nelle foreste naturali i pattern di rinnovazione sono in prevalenza basati su di un insediamento sottocopertura degli alberi più longevi e quindi sulla presenza contemporanea e per periodi temporali più o meno lunghi, di generazioni sovrapposte. Se consideriamo l’intero ciclo vitale del popolamento le strutture così dette saltuarie ed in modo particolare quelle che coinvolgono singoli alberi, sono molto rare in natura o quasi inesistenti per alcune specie, quali ad esempio il faggio ([71]).
Per questa specie sono state descritte in Italia strutture molto mosse in boschi antropizzati, assimilabili a soprassuoli disetanei a gruppi. L’origine di queste tipologie strutturali è imputabile al fatto che in passato le utilizzazioni seguivano criteri tesi a soddisfare richieste differenziate quali ad esempio assortimenti legnosi di valore il che comportava il taglio a scelta di alberi con determinate caratteristiche (spesso gli alberi di maggiori dimensioni). Successivamente il ripetersi sulla stessa superficie della “consuetudine di utilizzazione”, ha permesso di mantenere nel tempo quelle “irregolarità di struttura” sufficiente per una autoregolazione ed autoconservazione[21].
Nel nostro contesto, preme sottolinearlo ancora, se si escludono i casi di totale conservazione, si deve affrontare la scelta e l’applicabilità di modelli definiti in considerazione anche della reale situazione in cui si opera.
Ove sia possibile si dovrebbe puntare alla creazione di quella irregolarità di struttura purché questo non comporti forzature eccessive alla tendenza evolutiva del popolamento.
L’esperienza del modello colturale delle peccete disetanee del Comelico (Veneto - [3]) merita essere citato anche perché dimostra in modo evidente la validità del modello individuato nel 1951 e da Susmel codificato nel 1980 a livello regionale, così come si è accennato in precedenza. I risultati di studi condotti per oltre un decennio hanno evidenziato come lo sviluppo di modelli di gestione a flessibilità diversificata, possono conciliare nello spazio e nel tempo emergenze diverse in relazione alle funzioni che potrebbero essere richieste anche in futuro alla foresta stessa. Ad esempio è emerso che una valutazione della biodiversità trova grande rilevanza se non altro perché la perdita anche parziale di una parte di questa potrebbe avere effetti negativi sul funzionamento dell’ecosistema. Il trattamento disetaneo trova i fondamenti della sua applicabilità in questi boschi perché di fatto vi è stata una applicazione continuata nel tempo delle utilizzazioni che ha sempre considerato la salvaguardia della potenzialità produttiva del bosco allo stesso livello dell’interesse del prodotto ricavabile (anche economico). D’altra parte il selvicoltore, per le condizioni favorevoli stazionali, in questo caso poteva disporre di un’ampia probabilità di riuscita della rinnovazione naturale in tempi relativamente brevi. Più in generale i sistemi forestali produttivi dell’area alpina del Cadore interessati dal trattamento disetaneo, manifestano elevata polifunzionalità, stabilità stazionale e biologica tanto che possono essere considerati modelli colturali di riferimento.
Seguendo il filo conduttore di pensiero che considera come soggetto di riferimento la ricerca del migliore compromesso tra massima naturalità e selvicoltura, fondamento essenziale di una corretta gestione sostenibile delle risorse forestali, non possiamo escludere, anzi dobbiamo considerare, altri modelli colturali che a tutti gli effetti possono essere inclusi in un contesto di selvicoltura naturalistica. Ciò perché la selvicoltura che sebbene si identifichi nel modo più palese nell’operazione guidata dell’intervento di utilizzazione, resta comunque strumento di gestione insostituibile anche in un contesto eco-compatibile. Difatti in situazioni opposte, quali ad esempio quelle che prevedono conservazione assoluta, la selvicoltura non trova spazi applicativi[22].
Poiché l’intervento selvicolturale finalizzato all’ottenimento della rinnovazione naturale per lo meno in una prima fase di un percorso gestionale diretto verso la realizzazione dei presupposti (ad es., struttura del bosco) per l’applicazione di forme d’uso più vicini alla naturalità, può essere compreso in un insieme ecosostenibile, i modelli che fanno riferimento ai sistemi a tagli successivi possono risultare facenti parte di una selvicoltura naturalistica.
Può essere criticato il fatto come questi sistemi selvicolturali, in alcuni casi, dovuti spesso a fattori economici e gestionali, prevedano una uniformità del soprassuolo anche su vaste superfici; d’altra parte nelle foreste naturali, come già indicato, le fasi successionali di sviluppo di biostasi o bosco maturo possono permanere anche per diversi secoli. È la fase di decrepitezza o stadio cronologico tardo-senile che nel bosco antropizzato, spesso viene ad essere fortemente accorciato con i tagli di rinnovazione, mentre nei boschi naturali questo stesso può durare molto a lungo.
Non è quindi la presenza di per sé di un soprassuolo con uniformità nei confronti della posizione spaziale delle chiome ad inficiare a priori l’applicabilità di un modello colturale: hanno certamente più valore le modalità specifiche di come si opera o come si applica il modello.
I tagli successivi prevedono particolari modalità di applicazione altamente flessibili tanto che per alcune specie si assiste alla convergenza tra questi e i modelli saltuari irregolari o a gruppi; i primi tra l’altro vengono indicati strumento efficace per un cambiamento (trasformazione) del trattamento dal bosco coetaneo a quello disetaneo.
Trova conforto il fatto che il raggiungimento dell’obiettivo della rinnovazione naturale, tende a unire più che a separare, pur fornendo strade diverse, in una selvicoltura naturalistica eco-compatibile, una libertà di scelta di utilizzazione (free choice of felling) che rappresenta principio innovativo moderno in una visione di conservazione della biodiversità, funzionalità e stabilità degli ecosistemi forestali.
Resta aperto il problema relativo al fattore tempo. Nel caso in cui la ricerca mettesse in chiara evidenza la superiorità di un modello colturale rispetto ad altri, saremmo portati ad adottarlo ovvero a realizzare la sostituzione di quelli applicati.
In questo caso il riferimento al fattore tempo acquista un grande significato in quanto nel settore forestale i tempi di realizzazione sono sempre molto lunghi non esistendo procedure e metodi capaci di abbreviare i cicli biologici; la selvicoltura sotto questo punto di vista può arrecare solo un piccolo contributo nel favorire le fasi successionali.
Ad esempio la trasformazione di un bosco coetaneo di faggio in fase di biostasi ed in condizioni favorevoli alla rinnovazione ([52]), verso un bosco disetaneo, può essere presa in considerazione e si può configurare con un successo.
Altri fattori entrano però in gioco. Nel caso siano prevalenti le posizioni conservative (aree protette, riserve integrali) l’intervento selvicolturale di trasformazione non avrebbe senso; tra l’altro la fase di biostasi è propria della dinamica successionale della faggeta naturale e non vi sono motivazioni ne convenienza effettiva (anche sotto gli aspetti dei costi degli interventi) perché questa non possa compiersi nella più completa naturalità. Diverso potrebbe essere il caso di pervenire ad una maggiore stratificazione (o diversità ) delle chiome del popolamento coetaneo per boschi gestiti dall’uomo ovvero dove è atteso il prelievo della produzione legnosa.
Più in generale è possibile indicare che attraverso l’applicazione di tagli successivi non uniformi (per esempio a gruppi) e non prefissando un periodo di rinnovazione dell’intero bosco (particella), realizzare la trasformazione verso il bosco disetaneo. Il tempo richiesto è difficile da valutare, molto probabilmente superiore al secolo e per periodi così lunghi può essere molto aleatoria qualsiasi previsione.
L’insegnamento che emerge da questa casistica riguarda il fatto che anche nel convincimento della superiorità di certi trattamenti rispetto al altri, se si opera in un contesto in cui la finalità prioritaria riguarda la rinnovazione naturale, gli sforzi del selvicoltore devono concentrarsi sull’affinamento e sul miglioramento del trattamento in atto cercando di interpretare e seguire le indicazioni della natura. Spesso la pazienza dell’attesa assume saggia importanza. Le modalità che sono state seguite nel nostro Paese, ad esempio nel caso della conversione di molti boschi cedui rappresenta un esempio negativo da tenere presente. Difatti la volontà di operare comunque con interventi di avviamento senza conoscerne (o non avendo i dati di riferimento) gli “effetti biologici” del così detto taglio di avviamento, ha determinato in molti casi, un diffuso peggioramento della struttura e della composizione dei soprassuoli rispetto a quanto invece si sarebbe voluto ottenere.
Vincoli e libertà nelle aree protette
Nei paragrafi precedenti, anche se in forma condensata, sono state illustrate alcune delle conoscenze eco-biologiche di base che rappresentano punto di riferimento per la ricerca e per l’applicazione di linee di gestione eco-compatibili per aree forestali soggette a particolare attenzione di conservazione.
Nella consapevolezza di una reciproca interdipendenza fra conservazione dell’ecosistema bosco ed attività antropica e considerando anche quanto indicato in un precedente lavoro ([31]) si sono individuati alcuni scenari per i quali sono prevedibili interventi specifici e puntuali.
Foreste di conservazione
In questo primo caso gli aspetti conservativi sono prioritari ed interessano il paesaggio, l’ecosistema, le specie. Gli ecosistemi forestali di riferimento svolgono funzioni incentrate sulla massimizzazione della stabilità funzionale e quindi della biodiversità biologica.
È coinvolta comunque anche una conservazione ambientale che si riflette nell’azione di difesa esercitata dalla presenza del bosco stesso. L’orografia del nostro paese caratterizzata da accidentalità, ma anche da precarietà e instabilità dei versanti, impone una valutazione anche sotto questo aspetto che travalica il fatto che la foresta sia inclusa o meno in un territorio protetto[23].
Nel caso della conservazione della variabilità del germoplasma, questa potrà essere gestita in modo differenziale in riferimento alle informazioni fornite da un’analisi genetica puntuale ed accurata. Ovvero attraverso la conoscenza della distribuzione nello spazio e nel tempo delle frequenze geniche e genotipiche, sarà possibile indicare quali siano le popolazioni a più alta variabilità genetica e che richiedono quindi una maggior attenzione per la conservazione.
Nella stessa categoria rientreranno anche quei soprassuoli caratterizzati dalla presenza di specie relitte o minacciate per le quali la protezione dovrà essere estesa anche alla difesa da fattori inquinanti da parte di gene flow non desiderato. In questi casi l’approccio selvicolturale dovrà prevedere anche interventi di ripristino artificiale finalizzato.
Rientra a pieno titolo in questo scenario la tipologia dei soprassuoli descritti come Old-growth forests alle quali è assegnata l’identità più ampia di naturalità per la presenza di cicli successionali di maturità biologica e come tali svolgono il ruolo di vere risorse genetiche ([59]).
È utile rimarcare il fatto che l’età di un ecosistema foresta rappresenta un indice del suo stato successionale al cui variare corrispondono variazioni nel numero e nella frequenza degli organismi viventi che sono presenti e/o coinvolti in esso.
La maturità biologica differisce dalla maturità economica alla quale afferiscono anche i modelli dettati da una selvicoltura naturalistica che prevede comunque una utilizzazione.
Foreste polifunzionali
Sono le foreste di specie autoctone coltivate per le quali sono prevedibili diversità d’uso.
La coltivazione si attua con l’individuazione e l’applicazione di modelli colturali riferibili ad una selvicoltura incentrata all’ottenimento della rinnovazione naturale del soprassuolo.
Anche se prevalente resta la pressione conservativa, proprio perché si fa riferimento ad aree protette, la coltivazione persegue l’obiettivo di raggiungere il migliore equilibrio dinamico fra le funzioni produttiva, protettiva ed estetico-ricreativa della foresta.
In generale è dominante l’esigenza e quindi l’impegno, di mantenere un flusso continuo di prodotti e servizi il quale nello stesso tempo non può arrecare riduzione nei valori intrinseci o effetti indesiderati.
Per le specie che edificano i popolamenti della fase climax e per condizioni stazionali in cui le osservazioni in natura indicano buone possibilità di rinnovazione naturale, i cicli di maturità possono essere abbreviati (150-200 anni) rispetto a quelli valutati in un contesto di maturità biologica.
I dati sperimentali sono in questo specifico settore, relativamente scarsi, ma di estremo interesse. Ad esempio in boschi di faggio dell’Appennino Tosco-Emiliano (Foresta di Abetone, PT) è stato studiato l’effetto dei tagli successivi sulla struttura genetica del popolamento. Asportando il 25-30% della massa legnosa con il taglio di sementazione realizzabile con le piante di diametro più piccolo, i soprassuoli (in fase di biostasi) prima e dopo il taglio risultano geneticamente simili con valori di distanze genetiche inferiori al 4% ([70]).
Come in precedenza indicato valori di stabilità genetica sono stati trovati ([32]) in peccete miste del Cadore applicando il taglio di curazione in popolamenti articolati in eco-unità contigue ed interconnesse e rispettando la norma basata sulla statura delle piante più alte del piano dominante ([85], [80]).
Più in generale per alcuni tipi di soprassuoli in regimi di stabilità, si possono indicare modelli colturali che permettono una efficace polifunzionalità, ecosostenibile e che nello stesso tempo utilizzano a pieno la fertilità stazionale.
Nel bosco colturale così strutturato resta da verificare quanto la differenza tra età dei cicli di maturità naturale e quelli di maturità economica possa incidere rispetto ai limiti sopra indicati.
Per meglio chiarire i concetti appena enunciati si riassume come esempio quanto veniva riportato in un lavoro precedente sull’argomento ([31]), le cui conclusioni sono ancora pienamente attuali.
Il piano di assestamento della Foresta di Abetone - PT ([16]) prevedeva, per i boschi coetanei puri di faggio trattati a tagli successivi uniformi, un turno di maturità di 85 anni ed un periodo di rinnovazione di 15 anni. Questi dati, che emergevano da accurate indagini incrementali in bosco, evidenziavano le favorevoli condizioni in cui questa specie si veniva a trovare in quell’area dell’Appennino tosco-emilano. L’adozione di un turno di maturità economica così basso, determinava l’utilizzazione di piante con dimensioni diametriche dei tronchi molto ridotte per cui la percentuale degli assortimenti “da sega o da trancia”, quelli di maggior valore commerciale, risultava insignificante al taglio di sementazione, così anche con i tagli secondari e leggermente più alta con il taglio di sgombero (i fusti degli alberi eliminati con il taglio finale avevano usufruito di un più lungo periodo di isolamento ed il faggio è specie che reagisce a quest’effetto anche in età avanzata). Dopo alcune verifiche (Giannini, dati non pubblicati) eseguite in test-site permanenti per circa un ventennio si conferma che:
- per raggiungere una “sostenibilità economica” nei boschi di faggio dell’Appennino Tosco-Emiliano, è necessario disporre di almeno di una percentuale del 30-35% della massa cormometrica di “assortimenti nobili” (ad es., toppi da trancia per compensati);
- per raggiungere questo minimo di “sostenibilità economica” occorre allungare i) il turno di maturità economica, ii) il periodo di rinnovazione a 20-25 anni[24].
Per quanto riguarda il turno non sussistono problemi: i boschi di faggio, come la maggior parte delle latifoglie caratteristiche delle fasi finali del dinamismo successionale naturale, mantengono una efficacissima “stabilità” per tempi molto lunghi (200-300 anni) anche in popolamenti densi e monostratificati (fasi di maturità ) e su vaste superfici. Per il periodo di rinnovazione, il quale richiede più attenzione, occorre considerare il fatto che la sostituzione di un soprassuolo attraverso la rinnovazione naturale deve avvenire esclusivamente con novellame morfologicamente equilibrato e non sofferente[25]. Nello stesso tempo la regolazione della densità del popolamento durante tutto il suo ciclo di maturità deve essere prevista in modo tale da “sollecitare” le singole piante a crescere, senza però creare, in un momento non desiderato, condizioni favorevoli all’insediamento di una pre-rinnovazione che sarà inaffidabile per una sostituzione del soprassuolo e di ostacolo a quella guidata dal selvicoltore.
L’insegnamento ultimo di questo esempio riguarda il fatto che la ricerca di una “stabilità economica” specifica per le faggete dell’Abetone - Pian di Novello (PT) determina contemporaneamente migliori situazioni di stabilità polifunzionale e stabilità genetica.
Foreste di produzione, di specie esotiche a rapida crescita ed Arboricoltura da legno
Occorre subito considerare che, nell’ambito di aree protette, questo scenario può verificarsi per situazioni già esistenti, mentre è da ritenersi improponibile come possibilità progettuale.
D’altra parte la casistica del man-made forest non può essere oggetto totalmente discriminante.
L’incremento del consumo di legno a livello mondiale impone di considerare con sempre maggiore interesse nuove strategie per incrementare la produzione di biomassa. Tra l’altro risulta necessario trovare in tempi brevi una destinazione produttiva alle aree rese disponibili dall’abbandono delle attività agricole e zootecniche. Infine incrementare la produttività forestale con particolari sistemi o modelli colturali e concentrare su questi i prelievi per i consumi, significa alleggerire dalle utilizzazioni i boschi esistenti molti dei quali necessitano di “riposo” per ripristinare in sé quella soglia di efficienza e funzionalità capace di utilizzare in pieno la fertilità stazionale.
Sono noti dalla letteratura gli ottimi risultati ottenuti da piantagioni di conifere e latifoglie che sottoposte a regimi intensivi di coltivazione hanno consentito produzioni di biomassa da 3 a 7 volte superiori a quella ottenibile in boschi naturali.
Una selvicoltura intensiva richiede di accettare alcuni compromessi: una riduzione della ricchezza di specie ed una semplificazione dell’ecosistema. Le foreste edificate per via artificiale con una unica specie sono le più vulnerabili e spesso sotto questa ottica le giustificazioni economiche diminuiscono quando si analizzano i probabili costi di lotta e di restauro per situazioni compromesse ([47]).
È anche vero che le motivazioni sulle esigenze produttive hanno un peso di non poco conto. Da oltre 5000 anni l’uomo ha trasferito da una parte all’altra della terra piante di interesse e di valore commerciale. Così in Nepal si coltivano le patate, in Italia diventa nazionale, il pomodoro, un frutto sud-americano! Questo approccio è di interesse anche per gli alberi forestali.
Il caso più recente è quello della Leucaena sp., specie nativa dell’America latina, considerata un “miracolo vivente” dai suoi sostenitori, in quanto può crescere fino a 15 metri in 5 anni, produrre legno per fabbricati e quale fonte di energia, produrre foraggio per gli animali e cibo per l’uomo, migliorare le caratteristiche chimiche del suolo attraverso l’azoto fissazione oltre a possedere elevata resistenza a stress biotici ed abiotici ([12]).
Certamente l’introduzione di nuove specie determina considerevoli benefici, ma a loro volta impone anche notevoli costi.
Non possiamo tralasciare di citare il fatto che quando una specie viene introdotta questa può eliminare una o più specie locali determinando cambiamenti anche di vasta portata (l’introduzione della robinia o della douglasia, ha, ad esempio, modificato la tipologia dei funghi saprofiti tipici del consorzio naturale dell’area di distribuzione favorendo la diffusione dei “chiodino”nel caso della prima specie, impedendo la presenza del “porcino”, nel secondo).
Dobbiamo riconoscere comunque il successo proprio in Italia di alcune specie introdotte il cui impiego nasceva dallo schema propositivo di Aldo Pavari che considerava l’introduzione una integrazione per superare l’impossibilità o le carenze di specie indigene a soddisfare pienamente l’aumento in tempi relativamente brevi della produzione legnosa.
Ma in alcuni casi il fine ultimo può essere quello di massimizzare le produzioni di qualità nelle condizioni migliori stazionali adottando metodi di coltivazione intensiva.
Sotto l’aspetto economico-finanziario nell’arboricoltura da legno si ricerca il massimo utile. La produzione legnosa si inserisce in un contesto sempre più specifico del mondo agrario in cui l’ecosistema si semplifica e si caratterizza per incrementi congiunti produttività /input energetici e decrementi proporzionali dell’integrità ecosistemica/diversità biologica.
L’ecosistema così semplificato, può avere una durata anche molto breve (6-12 anni con ibridi di pioppo) e può non essere ripetibile essendo legato a cicli di maturità economica di “natura tecnica” riguardando assortimenti ritraibili da specie ad elevata produttività a richiesta differenziata sul mercato.
Restano invero la perplessità circa la vulnerabilità degli ecosistemi edificati da pochi genotipi ed il fatto che pochi sono oggi i tipi che possono essere disponibili sul mercato.
Un analisi appropriata degli scenari descritti, porta a considerare l’importanza di una definizione cosciente di linee multi funzionali di uso del territorio in particolare nel caso che questi ricada in aree soggette a protezione. D’aiuto a questo è certamente l’analisi accurata delle possibilità applicative in relazione anche a fattori tempo e spazio nonché alla puntuale situazione nei confronti dei diversi gradi di tutela dell’area oggetto di protezione.
Considerazioni conclusive
Il raggiungimento di situazioni prossime alla foresta naturale (near-nature forest) dovrebbe essere favorito nelle aree protette. Per molti tipi di boschi presenti in queste aree dominate da climi temperati e caratterizzati da mescolanza di specie (in particolare latifoglie), il traguardo da raggiungere dovrebbe essere: i) pervenire nell’ambito di superfici più o meno ampie, a variazioni nella struttura del soprassuolo arboreo e ii) favorire la presenza, in porzioni diffuse del popolamento, di fasi di sviluppo dominate da alberi biostatici (stramaturi) e di grandi dimensioni associati a elevate quantità di legno in decomposizione (tronchi morti, sul suolo, etc. - [27]). Questo traguardo si può raggiungere attraverso i) l’istituzione di risorse forestali integrali e/o orientate, ii) una gestione a favore della biodiversità ed infine iii) una conservazione della stabilità della foresta.
A livello europeo il primo punto è stato soddisfatto in diversi paesi ([13]) reclutando tali aree nell’ambito di boschi polifunzionali. Il valore biologico di queste iniziative può essere incrementato attraverso oculati e puntuali interventi colturali tesi a favorire nel tempo strutture spaziali a mosaico ([72]) nonché dare avvio alla fase dinamica iniziale creando aperture ed interruzioni nella copertura (gap-phase) della chioma del soprassuolo (ad es., taglio di sementazione su piccole superficie).
Tutto ciò deve rappresentare solo un aiuto alla dinamica evolutiva fino al raggiungimento di una stabilità strutturale, di processi e di habitat tipici delle foreste naturali ([27]).
Siamo consapevoli che tali indirizzi possono scontrarsi con il pensiero di alcuni che sono favorevoli comunque a non prevedere alcun intervento.
D’altra parte abbiamo già fatto presente che la diversità di pensiero non ha motivi per essere origine di contese né tanto meno di pragmatismi.
Si intuisce che coloro che sono favorevoli all’intervento (oculato e puntuale) vedano in questo lo strumento per generare valori biologici e processi in relazione al fattore tempo (sempre molto lungo): sotto certi aspetti indicano che prima si raggiunge l’obiettivo meglio è.
È necessario, tra l’altro, fare altre considerazioni. La prima riguarda la necessità di realizzare delle riserve forestali (foreste di conservazione). Poiché gli obiettivi potranno essere raggiunti solo dopo un periodo di tempo anche molto lungo, è necessario che il soprassuolo primitivo abbia capacità di resistenza e resilienza perché almeno alcuni alberi raggiungano quella età e quelle dimensioni che consentano ad essi stessi, di divenire i capisaldi della foresta naturale (Old-growth forests).
L’esperienza dimostra ad esempio che i boschi coetanei di conifere (boschi coltivati) sono generalmente boschi più “fragili” a parità di altre condizioni, dei boschi di latifoglie per cui già questa caratteristica differenziale può essere motivo nel propendere verso una soluzione rispetto ad un’altra.
L’esperienza documentata attraverso i modelli sviluppati da Emborg et al. ([27]) per la foresta semi vergine di Suserup Skov in Danimarca, indica con chiarezza la possibilità di intervento intesa come guida per una trasformazione dei boschi coetanei in disetanei tra i quali possono essere incluse le faggete.
Merita anche ricordare quanto indicato da Susmel ([78]) circa i modelli di coltivazione elaborati per i boschi del Cadore orientale ed occidentale che individuavano il più corretto compromesso tra i ripristino di una funzionalità ecosistemica del bosco e le possibilità di prelievo legnoso quale fonte economica delle popolazioni locali.
Ecco che nel caso di aree protette, anche dove i soprassuoli sono fonte economica importante, è da suggerire l’adozione di interventi di coltivazione che seguano il più possibile la tendenza naturale di sviluppo del bosco e che prevedano procedure guidate perché lo stesso ecosistema subisca disturbi su piccola scala[26].
In altri termini l’implementazione di modelli selvicolturali deve basarsi sul principio della automazione naturale (principle of nature automation) che prevede, per le attività forestali, la massima utilizzazione dei pattern dei processi naturali spontanei ([23], [27]).
In questo caso potremmo individuare una fitness selvicolturale sintesi di due strategie operative prevalenti la prima della quali, ecologica, in riferimento alle condizioni stazionali e la seconda, genetica, risultante dagli effetti congiunti dei fattori evolutivi.
Perseguendo un pensiero strettamente legato alla conservazione dell’ecosistema foresta emerge con chiarezza che lo sforzo dominante è la protezione della biodiversità.
In essa è espressa la conservazione ed il mantenimento della variabilità genetica che rappresenta strumento di preservazione del tasso di adattabilità delle specie e punto di partenza per una continuità evoluzionistica che è necessario garantire.
In riferimento a ciò alcuni problemi potrebbero sorgere quando nell’ambito di un principio generale di conservazione si passa a considerare il metodo per realizzarla.
Ad esempio dovendo ricorrere alla rinnovazione artificiale potremmo domandarci quante informazioni disponiamo per valutare se le operazioni standardizzate dalla selvicoltura rispondano veramente ai principi fondamentali della conservazione stessa e chiederci ancora quale potrebbe essere il danno di un eventuale inquinamento genetico o quello arrecato dalla riduzione della variabilità.
Molte delle specie che edificano i principali ecosistemi forestali, a causa della loro distribuzione naturale, non rispettano i confini amministrativi di Stati o Nazioni. Sotto questo aspetto la conservazione assume carattere internazionale e quindi ai singoli Stati potrebbero essere anche imputate responsabilità molto forti.
Se spostiamo la discussione a scala di area geografica (regione, provincia) un primo quesito riguarda ad esempio quale livello di rarità può essere assegnata la presenza di una specie o di una popolazione.
Se la specie è frequente può essere oggetto di attenzioni differenziate o divenire di scarso interesse. Molte delle aree protette in Italia (1160 secondo il T.C.I. nel Volume informativo del 1999) riguardano la presenza di una particolare specie o endemismo in un determinato contesto territoriale.
Ma l’interesse può riguardare anche la forma e la continuità dell’areale ed allora, ad esempio, le popolazioni marginali, quelle disgiunte o isolate, dovrebbero ricevere più attenzione conservativa? È probabile di si, in quanto non si può escludere che esse rappresentino il risultato di azioni diversificate dei fattori evolutivi, rispetto a quanto questi hanno operato nelle aree centrali dell’areale stesso.
Sottolineando quanto è stato indicato in precedenza una prima conclusione di carattere generale, ma che assume particolare significato nel caso di aree protette, riguarda il fatto che mantenendo ecosistemi forestali ad elevata diversitàè possibile sperare di conservare nel tempo livelli soddisfacenti di produttività.
Gli alberi forestali, organismi molto longevi, ad alta eterozigosità nell’ambito di un genoma molto ampio, si caratterizzano per una elevata plasticità; la marcata dispersione dei flussi pollinici e del seme nonché la presenza di particolari sistemi di riproduzione, controllano un continuo ed intenso rimaneggiamento genico.
È questo forse il segreto rivelato del perché tali particolarità hanno come conseguenza la realizzazione nella foresta di grandi serbatoi di variabilità genetica e di diversità.
In effetti, se consideriamo l’intero ecosistema, ancora oggi molte porzioni di questo restano sconosciute: basti pensare alla componente microbica nei primi 10 cm di suolo ([85], [84]). Anche dove l’azione di conservazione è intensa e determinata, sappiamo, sotto l’aspetto genetico, cosa effettivamente si conserva?
L’attività antropica è stata causa per lungo tempo - e lo è tuttora - di modifiche degli ecosistemi naturali creando, come prima conseguenza, catene trofiche semplificate, ed in generale stadi di degrado che in parte derivano da azioni dirette, ma anche indirette attraverso alterazioni ambientali quali l’inquinamento.
L’interrogativo è se esistono, nell’ambito delle popolazioni (nel nostro caso quelle degli alberi forestali), le condizioni di fondo di adattamento, ovvero se sono presenti individui dotati di fitness per cui le generazioni successive possono garantire la presenza di comunità adattate a scenari mutevoli. Questo è importante proprio perché gli alberi, organismi di lunga vita, sono coinvolti in modo più spinto alla dinamicità di questo adattamento evolutivo il quale è regolato dalla velocità del cambiamento ambientale e dall’intervallo di tempo che intercorre fra generazioni successive.
Cambiamenti a livello di fitness per presenza/assenza o per dimensione dei discendenti possono portare a perdita di alleli. La conservazione di stazioni di rifugio e la realizzazione di corridoi di gene flow rientrano tra le strategie operative di corretto interesse per una conservazione in-situ delle risorse del germoplasma.
Più in generale appare evidente come le risorse genetiche ed i processi responsabili della loro evoluzione nello spazio e nel tempo, rappresentino componente vitale di una selvicoltura sostenibile close to nature.
Note al Lavoro
L’ultima fatica del Prof. Lucio Susmel, ha riguardato questo lavoro congiunto con il Prof. Raffaello Giannini sulle problematiche della gestione delle foreste nelle aree protette. Il lavoro si inseriva in una tematica di una più vasta ricerca proposta dal Prof. Umberto Bagnaresi approvata dall’Accademia Nazionale di Agricoltura e finanziata dal Ministero delle Politiche Agricole e Forestali, che aveva come obiettivo quello di fornire un contributo propositivo all’evoluzione della concettualità ispiratrice della “politica agricola” delle aree protette ed alle metodologie per qualificare gli interventi. Una prima stesura del lavoro è stata resa nota in un volume specifico dell’Accademia. La redazione ha ritenuto importante che il contributo scientifico dei due Autori avesse una più puntuale diffusione nel mondo forestale decidendone la pubblicazione.
References
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar