
Temporal scale of the action of temperature and photoperiod on the xylogenesis at the treeline
Forest@ - Journal of Silviculture and Forest Ecology, Volume 4, Pages 6-10 (2007)
doi: https://doi.org/10.3832/efor0444-0040006
Published: Mar 21, 2007 - Copyright © 2007 SISEF
Short Communications
Abstract
The possible role and interaction of two ecological factors, photoperiod and temperature, on xylogenesis of conifers growing at treeline are review. In cold climates, temperature is one of the major factors regulating metabolic activities of plants and influencing onset and rate of cell production during the growing period. However, photoperiod was found to act as a signal regulating the timing of maximum growth rate and synchronizing radial growth at annual level. During tree-ring formation, both factors are crucial but with different levels of interaction. Some recent findings about xylogenesis in cold environments are described and discussed
Keywords
Alps, Xylogenesis, Cellular differentiation, Thermic thresholds, Treeline
La treeline è una linea immaginaria che connette i gruppi isolati di alberi (con fusti eretti di almeno tre metri di altezza) situati nel limite altitudinale o latitudinale della loro distribuzione naturale. Nei massicci montuosi italiani, questa linea delimita una fascia ecotonale compresa fra il bosco denso subalpino e le praterie di alta quota (Fig. 1A-C). È ormai largamente accettata l’ipotesi che questi limiti altitudinali e latitudinali del bosco siano strettamente collegati alla temperatura. A livello planetario infatti, la posizione delle treeline decorre parallelamente al limite termico individuato dall’isoterma di 0°C del mese più caldo dell’anno ([6]) e i limiti superiori del bosco vengono osservati dove la temperatura media stagionale del suolo si assesta intorno ai 6.7°C ([7]).
Fig. 1 - Ambienti forestali di alta quota in Italia. A: limite superiore del bosco sulla Tofana di Rozes (Cortina d’Ampezzo - BL); B: larici-cembreto con abete rosso al passo Giau (Cortina d’Ampezzo - BL); C: alpeggio in ambiente di treeline a Croda da Lago (Cortina d’Ampezzo - BL); D: prateria alpina a nardeto ai margini delle ultime avanguardie arboree nei pressi delle Cinque Torri (Cortina d’Ampezzo - BL); E: popolamento subalpino misto di larice e pino cembro con individui di Pinus uncinata a Mont Fort du Beauf (Clavière - TO); F: popolamento rado di pino loricato sovrastanti la faggeta pura a Serra del Crispo nel Parco Nazionale del Pollino (San Severino Lucano - PZ).

Non sorprende che la temperatura sia uno dei fattori ecologici cruciali per la vita in ambienti freddi. Tuttavia, pur essendo state sviluppate e discusse diverse ipotesi in merito ([18], [8], [19]), rimane ancora da dimostrare in che modo questo fattore influenzi i processi di accrescimento delle piante. Non è stata infatti ancora fornita una spiegazione definitiva sui meccanismi cellulari in cui agisce la temperatura, anche se alcuni indizi sembrerebbero condurre verso l’impossibilità da parte dei tessuti vegetali di investire il carbonio prodotto dalla fotosintesi nella produzione e differenziazione di nuove cellule (growth limitation hypotesis, [6]). Fino a qualche anno fa non si era in grado di fornire una spiegazione funzionale e determinare il ruolo e l’importanza intrinseca della temperatura e dell’eventuale associazione o interazione con altri fattori ecologici nell’accrescimento delle piante, in particolar modo a scala cellulare, alla scala cioè a cui avviene la formazione dei nuovi tessuti.
Recentemente, diversi progetti di ricerca canadesi ed europei (fra cui alcuni italiani[1]) hanno permesso di confermare il ruolo della temperatura nella formazione del legno anche a livello cellulare. In Finlandia, l’avvio della xilogenesi in Pinus sylvestris e Betula pendula è fortemente controllato dalla temperatura ([17]). Deslauriers & Morin ([2]) hanno dimostrato che, nel legno primaticcio di Abies balsamea in Canada, a temperature più elevate corrispondevano produzioni cellulari maggiori. Rossi et al. ([14]) hanno confrontato la fenologia delle cellule xilematiche delle principali conifere della treeline alpina in Italia con le temperature dell’aria, del fusto e del suolo per definire i valori delle soglie termiche necessarie per lo svolgimento dei processi di crescita nel tronco. In Larix decidua, Pinus cembra e Picea abies, la xilogenesi è attiva in primavera quando la temperatura minima giornaliera dell’aria raggiunge 2-4°C e quella del fusto 4°C. La convergenza delle temperature soglia fra i diversi siti analizzati, le specie e gli anni di studio, ha dimostrato l’esistenza di precisi limiti termici nell’accrescimento radiale e nella formazione dell’anello legnoso. Simili risultati sono stati confermati anche da analisi istologiche su Pinus leucodermis del massiccio del Pollino ([1]).
Il cambio è un importante utilizzatore di carboidrati non strutturali e la stessa attività cambiale richiede un flusso continuo di energia sotto forma di zuccheri, estratti dai tessuti di riserva o prodotti dalla fotosintesi ([10]). Durante la maturazione cellulare inoltre, gli alberi investono una notevole quantità di carbonio nella sintesi e nella deposizione delle microfibrille di cellulosa per la costituzione della parete cellulare ([4]). Secondo Rossi et al. ([14]) la domanda di fotoassimilati da parte dei processi metabolici coinvolti nella crescita cellulare xilematica verrebbe limitata con temperature medie giornaliere inferiori a 6-8°C. Temperature simili sono state stimate anche per altri organi della pianta: l’allungamento dei getti di pino silvestre veniva inibito a temperature di 6-8°C ([5]) e l’accrescimento delle radici si riduceva drasticamente al di sotto dei 6°C ([20]). Sembrerebbe quindi che i medesimi valori di temperatura (collocati intorno a 6-8°C) influenzino i processi di crescita delle diverse parti dell’albero (getti, fusto e radici). La convergenza delle temperature soglia stimate sui diversi meristemi apicali e laterali farebbe quindi supporre l’esistenza anche di meccanismi simili in tutti i tessuti di una pianta in grado di regolare la crescita a livello cellulare.
Nell’ipotesi di un processo di accrescimento regolato unicamente dalla temperatura, la produzione cellulare seguirebbe strettamente l’andamento termico stagionale. Come è noto, l’andamento della temperatura dell’aria in tutte le regioni temperate e boreali dell’emisfero nord è campaniforme, con un graduale aumento primaverile, un picco massimo intorno alla seconda metà del mese di luglio ed una successiva diminuzione durante l’estate e l’autunno (Fig. 2A-D). Ci si aspetterebbe quindi che tutti i processi di produzione cellulari che si verificano in piante di ambienti fortemente dipendenti dalla temperatura si verifichino in sincronia con questo andamento, cioè con una culminazione dell’accrescimento intorno alla seconda metà di luglio. Un’analisi comparata della xilogenesi su diverse specie e siti di studio in Italia (Fig. 1D-F) ed in Québec (Canada) ha rivelato un aspetto finora sconosciuto nella formazione del fusto a scala annuale ([15]). Il confronto delle curve di formazione cellulare nel tronco ha evidenziato come la produzione dei tessuti xilematici si concentri nella prima parte della stagione vegetativa e che tutte le specie raggiungano i valori massimi di accrescimento nel medesimo periodo dell’anno, in prossimità del solstizio d’estate, quando la lunghezza del giorno, e non la temperatura, è massima (Fig. 2A-D). Dopo il 21 giugno, la produzione di cellule è destinata gradualmente ad esaurirsi. In qualche modo quindi, gli effetti della temperatura sulla formazione dei tessuti nelle piante viene mediata dal fotoperiodo, un fattore astronomico certamente indipendente dalla temperatura e costante nel tempo.
Fig. 2 - Temperatura media giornaliera dell’aria (°C) e lunghezza del dì (minuti) misurata nel 2004 a diverse latitudini ed altitudini. A: treeline alpina in Italia (Cinque Torri: 46°27’N, 12°08’E, 2080 m s.l.m.; Cortina d’Ampezzo, BL, Italia); B: foresta boreale in nord America (Schefferville: 54°48’N, 66°48’W, 518 m s.l.m.; Québec, Canada); C: foresta boreale in nord Europa (Tromso: 69°39’N, 18°56’E, 114 m s.l.m.; Troms, Norvegia); D: limite latitudinale del bosco nell’Asia settentrionale (Chokurdakh: 70°37’N, 147°53’E, 61 m s.l.m.; Yakutia, Russia). Dati raccolti dal Dipartimento TeSAF, Università degli Studi di Padova (A) e dal Russia’s Weather server (B-D).
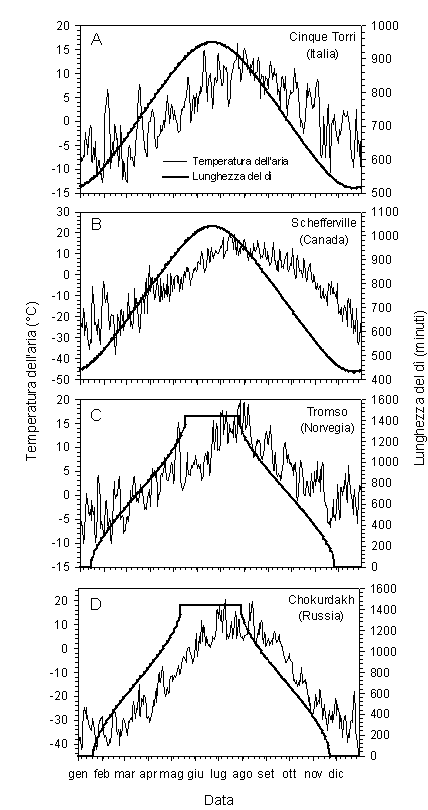
Le piante si sarebbero quindi evolute sincronizzando i ritmi di accrescimento con la lunghezza del giorno, che assume il ruolo di un vero e proprio calendario naturale. I vantaggi nella sincronizzazione dell’attività vegetativa con il fotoperiodo sono direttamente collegati alla necessità di evitare i periodi termicamente più sfavorevoli alla crescita. Un aumento della temperatura del fusto, infatti, induce la riattivazione delle divisioni cellulari nei meristemi cambiali solamente nei periodi tardo invernali e primaverili mentre non produce alcuna risposta in dicembre ([11]). Anche la data di schiusura delle gemme di specie di zone temperate e boreali è funzione della temperatura e della lunghezza del giorno ([9], [12]). Secondo Nizinski & Saugier ([9]), ad esempio, la soglia di temperatura cumulata che induce l’apertura delle gemme in Quercus petraea decresce durante la stagione con valori massimi in aprile e minimi in giugno.
Le piante devono assicurare il completamento della maturazione di tutte le cellule prodotte. Il fatto di completare le divisioni cellulari precocemente durante la stagione vegetativa consente alla pianta di destinare le risorse energetiche nella maturazione dei tessuti xilematici in formazione durante il periodo estivo. Nelle conifere di ambienti freddi, una cellula xilematica neoformata del legno tardivo richiede fino a 25-40 giorni per completare la differenziazione ([3], [16], [13]). Nell’ipotesi di divisioni cellulari protratte nella tarda estate, le ultime cellule prodotte non avrebbero il tempo sufficiente per completare la differenziazione (ad esempio l’ispessimento e la lignificazione delle pareti secondarie) prima dell’arrivo del periodo freddo, con una conseguente riduzione della funzionalità delle strutture xilematiche ed un aumento della sensibilità agli stress biotici ed abiotici.
Dai recenti progetti di ricerca svolti in ambiente di treeline, è emerso che, nei tessuti xilematici del fusto, la temperatura permetterebbe il mantenimento delle attività metaboliche come l’attività di produzione cellulare nel cambio e la maturazione delle cellule neoprodotte. Il fotoperiodo, invece, interverrebbe come segnale di regolazione del ritmo di accrescimento, e quindi di sincronizzazione della xilogenesi a scala annuale. Negli ecosistemi forestali di limite altitudinale e latitudinale, quindi, la temperatura ed il fotoperiodo rappresenterebbero due fattori ecologici cruciali per l’accrescimento radiale delle piante arboree, sebbene la loro influenza abbia effetti a scale temporali diverse.
References
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar













