Effects of soil characteristics on functionality and growth of forest stands: a sensitivity analysis of the model 3-PG
Forest@ - Journal of Silviculture and Forest Ecology, Volume 4, Pages 298-309 (2007)
doi: https://doi.org/10.3832/efor0473-0040298
Published: Sep 20, 2007 - Copyright © 2007 SISEF
Research Articles
Abstract
Al fine di predire la produttività forestale a larga scala occorre disporre di un modello affidabile che richieda in ingresso pochi parametri di facile misurabilità. Nel presente lavoro è stata condotta un’analisi di sensitività del modello a base funzionale 3-PG ([18]) relativamente alla fertilità, ritenzione idrica e tessitura del suolo, parametri di difficile misura. Lo studio è stato suddiviso in due parti. Nella prima è stata analizzata la risposta del modello, per un terreno sabbioso-limoso, a differenti profondità del suolo e concentrazioni di nutrienti. Mentre per l’evapotraspirazione non si evidenziano differenze in risposta alla quantità di nutrienti, la produttività primaria lorda, il volume e il LAI mostrano invece una risposta positiva all’aumentare sia della fertilità che del contenuto idrico, più marcata nel primo caso. Nella seconda parte del lavoro sono stati analizzati i meccanismi di azione della tessitura del suolo attraverso i suoi effetti sulla capacità di campo e per le differenze nella risposta degli scambi gassosi al potenziale idrico, per quattro differenti tipologie di suoli (sabbia, sabbioso-limoso, argilloso-limoso e argilla). Da questa seconda analisi è emerso come la funzionalità del bosco sembri favorita da tessiture fini, con i risultati peggiori costantemente osservati nel caso di suoli sabbiosi. L’effetto della tessitura appare legato in larga misura alla diversa capacità di campo dei suoli a differente granulometria. Una volta compensato tale effetto, infatti, i valori di crescita e funzionalità non sembrano differire significativamente fra i diversi suoli. In fase predittiva è possibile quindi, accorpare i due parametri legati alla tessitura del suolo riferendosi solo al quantitativo massimo di acqua senza commettere errori significativi. Trascurare sia tale parametro sia l’indice di fertilità comporta un errore nella stima della GPP del 30-32%.
Keywords
Modelli funzionali, carbonio, fertilità, profondità suolo, contenuto idrico, tessitura, produttività.
Introduzione
La possibilità di predire il tasso di crescita e quindi la produttività di un bosco a partire da dati iniziali facilmente reperibili è uno dei principali problemi per la gestione forestale, sia nel caso di boschi già esistenti sia per la programmazione di nuovi impianti ([2]).
Modelli empirici basati interamente su misure effettuate in campo, sebbene descrivano accuratamente il sistema bosco nell’intervallo di condizioni per cui sono stati costruiti e calibrati, non sono però estrapolabili al di fuori di tale intervallo e quindi non sono applicabili ad altri contesti con scopo predittivo ([4]).
Per superare tale limite risulta utile l’impiego di modelli che siano in grado di descrivere le dinamiche della pianta e del bosco basandosi sulla conoscenza delle condizioni ambientali e dei processi fisiologici che ne governano la crescita. Nel corso degli ultimi anni sono stati sviluppati molti di questi modelli, che riflettono le nuove conoscenze di fisiologia e le sempre migliori capacità di descrivere quantitativamente tali processi e le loro interazioni. Questi modelli a base funzionale, però, se da un lato descrivono fedelmente la realtà, dall’altro richiedono un numero molto elevato di parametri d’ingresso, molti dei quali sono di difficile, se non impossibile, misurazione.
Nasce, quindi, l’esigenza di avere a disposizione uno strumento semplice e affidabile, con pochi parametri d’ingresso facilmente reperibili, utilizzabile per la gestione forestale e applicabile a larga scala.
Il modello 3-PG ([18]), analizzato in questo lavoro, è un modello generale a base funzionale facilmente adattabile a qualsiasi sito e specie di piante, previa opportuna parametrizzazione.
Allo scopo di facilitare l’applicazione del modello ad un ampio numero di situazioni occorre ridurre lo spettro di parametri di ingresso ovvero analizzare quali fra essi siano realmente indispensabili e quali al contrario possano essere trascurati o accorpati. A tal fine nel presente studio è stata condotta un’analisi di sensitività della risposta del modello a differenti livelli di fertilità e alle caratteristiche idriche e di tessitura del suolo: parametri, questi, di non facile misura che hanno ricevuto una limitata attenzione da parte della ricerca ecofisiologica.
Tale ricerca si è infatti principalmente focalizzata sull’effetto dei parametri meteorologici di più comune misura (radiazione, temperatura, precipitazioni,...) o sugli effetti di fattori modificabili dall’uomo nei sistemi colturali intensivi, quale ad esempio la fertilità chimica. Temperature troppo alte o troppo basse, valori elevati di deficit di pressione di vapore, la mancanza di acqua o un suolo poco fertile sono causa di una diminuzione della capacità della pianta di fotosintetizzare, inibendone così la crescita; il verificarsi anche di una sola di queste condizioni può avere conseguenze estreme ([15]).
Una minore attenzione è stata invece prestata agli effetti di altri fattori stazionali, di non immediata misura e non facilmente manipolabili, ma che l’esperienza selvicolturale ci dice giocare un ruolo centrale nella crescita del bosco, come la profondità o la tessitura del suolo.
È nota ad esempio l’importanza della profondità dei suoli su funzionalità e crescita del bosco, soprattutto in ambienti mediterranei ([29]), a causa della maggiore quantità di acqua immagazzinabile e quindi, in linea di principio, sfruttabile dall’apparato radicale.
Molto meno si sa invece degli effetti sulla crescita delle piante della tessitura del suolo. Mentre è noto che nei climi boreali la crescita può essere limitata dalle condizioni di saturazione e anossia spesso associate a suoli a tessitura fine ([36], [25]), poco si sa della rilevanza della tessitura in ambienti mediterranei, dove l’acqua costituisce il principale fattore limitante.
Nel presente studio, gli effetti delle caratteristiche fisiche e chimiche del suolo sono state esplorate attraverso l’applicazione del modello ecologico 3-PG, con l’obiettivo di meglio comprendere i meccanismi alla base della interazione pianta-suolo, e di poter valutare quanto questi meccanismi incidano sulla produttività del bosco.
Materiali e metodi
I modelli a base funzionale cercano di saldare le conoscenze di dettaglio disponibili sui singoli processi biologici e sulle loro mutue interrelazioni per spiegare e predire quantitativamente le proprietà emergenti di sistemi complessi quali le piante e gli ecosistemi forestali. Il modello 3-PG, in particolare, cerca di catturare la funzionalità del bosco concentrando l’attenzione su un numero molto limitato di processi fondamentali, a loro volta rappresentati con la massima semplicità. Se da una parte questa rappresentazione schematica implica un minor realismo e una ridotta capacità predittiva, dall’altra essa permette di meglio individuare e comprendere le mutue interrelazioni delle parti e le dinamiche alla base delle proprietà emergenti del sistema. Tale semplicità si traduce inoltre in una più semplice parametrizzazione, rendendone possibile l’applicazione ad un ampio spettro di situazioni ([1], [10], [34], [19], [30], [8], [7]).
Descrizione del modello
Il modello 3-PG (Physiological Principles in Predicting Growth) è un modello a base funzionale di crescita del bosco ([18], [38], [30]). A partire dalla radiazione solare incidente mensilmente sulle foglie, considerando i vari fattori ambientali (temperatura, pressione di vapore, umidità, fertilità, deficit idrico del suolo, ecc.) e l’età del bosco stesso, è possibile ottenere una stima della produttività lorda del soprassuolo e della crescita delle varie parti delle piante (radici, fusto, chioma), assumendo che:
- la fotosintesi del bosco sia proporzionale alla quantità di energia luminosa assorbita dalla copertura ([24], [17]);
- a causa delle limitazioni ambientali (legate a contenuto idrico del suolo, deficit di saturazione, temperatura atmosferica, giorni di gelo) e dell’età solo una parte della radiazione fotosinteticamente attiva assorbita possa essere effettivamente utilizzata nei processi fotosintetici;
- il rapporto tra la produttività primaria netta (P N) e la produttività primaria lorda (GPP) sia in buona approssimazione costante (NPP/GPP = 0.47; [37]);
- l’allocazione delle risorse disponibili per la crescita all’apparato radicale aumenti in relazione ai fattori di stress e all’età del bosco ([31], [6], [22]);
- le risorse non utilizzate per la crescita radicale vengano ripartite tra fusto e foglie secondo equazioni allometriche appropriate per la specie;
- la conduttanza di copertura (somma della conduttanza stomatica delle foglie nell’intera chioma) e quindi la traspirazione del bosco raggiungano un valore massimo quando l’indice di area fogliare (LAI) raggiunge un valore di 3.0 m2/m2 ([21], [14]) e vengano modulate in funzione delle stesse limitazioni ambientali ed endogene sopra menzionate;
- la densità delle piante nel popolamento venga ridotta dai processi di competizione secondo la legge dell’auto-diradamento ([39]).
Per una descrizione di maggior dettaglio del modello si rimanda a Landsberg & Waring ([18]) e Waring & McDowell ([38]).
Risposta alla fertilità fisica e chimica del suolo
Poiché la risposta del bosco alle caratteristiche fisiche del suolo ed alla fertilità stazionale costituisce l’obiettivo della presente analisi, la trattazione di questi processi nel modello 3-PG viene descritta in maggiore dettaglio.
La fertilità chimica del suolo viene catturata empiricamente nel modello da un semplice parametro sintetico (FR, fertility ratio). Si assume che variazioni di fertilità stazionale influenzino tanto l’efficienza di uso della luce quanto la ripartizione della crescita fra radici ed apparati epigei. In particolare si assume che l’efficienza massima di uso della luce (a c; rapporto fra luce intercettata e fotosintesi in condizioni ottimali) aumenti linearmente all’aumentare della fertilità del suolo, a causa della maggiore concentrazione fogliare di azoto e proteine ([32]), mentre non si ha alcun effetto diretto di tale parametro su conduttanza stomatica e traspirazione del soprassuolo. All’aumentare della fertilità, diminuisce il coefficiente di allocazione agli apparati radicali (h r, frazione della produttività primaria netta utilizzata per la crescita delle alle radici fini preposte all’assorbimento dei nutrienti; [6]), aumentando la frazione disponibile per la crescita epigea (eqn. 1):
dove fage, fθ, fD e FR rappresentano, rispettivamente, i fattori di riduzione sugli scambi gassosi dovuti all’età, al contenuto idrico, al deficit di pressione e alla fertilità e possono assumere valori compresi tra zero e uno (assenza di stress).
Funzionalità e crescita delle piante sono inoltre influenzate dallo stato idrico del suolo e quindi dalle sue caratteristiche fisiche ([5]). Il modello 3-PG comprende un sottomodello per la stima del bilancio idrico stazionale. Dopo un’inizializzazione al massimo valore di acqua immagazzinabile (θ max), uguale al prodotto tra la profondità del terreno e la capacità di campo associata ad una particolare tessitura, il bilancio idrico viene aggiornato giornalmente, sommando le precipitazioni e sottraendo le perdite per evapotraspirazione. Qualora si superi tale valore, l’acqua in eccesso viene perduta (per percolazione o scorrimento superficiale).
Si assume che la profondità del suolo agisca sulla pianta solo attraverso i suoi effetti sulla capacità totale di ritenzione idrica, la quale permette alla pianta di affrontare periodi secchi facendo affidamento sulle riserve accumulate nella stagione piovosa. Si trascurano invece eventuali effetti legati all’aerazione del suolo, alla resistenza all’accrescimento delle radici o alla loro mortalità.
Valori di evapotraspirazione superiori alle precipitazioni determineranno deficit idrici del suolo crescenti, deprimendo l’evapotraspirazione stessa in un ciclo di retroazione negativa, così come l’efficienza di uso della luce e l’allocazione dei fotosintetati alla parte epigea. La risposta dei tre processi al deficit idrico è influenzata inoltre dalla tessitura del suolo, con una risposta più precoce e graduale nel caso di terreni argillosi e più brusca e tardiva nel caso di tessiture grossolane. L’effetto della tessitura riflette in maniera implicita la risposta funzionale della pianta al potenziale idrico piuttosto che al contenuto idrico del suolo ([32]) e la diversa ritenzione idrica di suoli di diversa granulometria ([5]). La relazione fra il contenuto idrico relativo del suolo (r θ, rapporto fra contenuto idrico del suolo e capacità di campo) ed il fattore di riduzione degli scambi gassosi (f θ, con valore compreso fra 0 e 1) è catturata dai due parametri c θ, il deficit idrico relativo cui corrisponde una riduzione del 50% degli scambi gassosi, ed n θ, un parametro empirico, secondo la relazione (eqn. 2):
I valori dei parametri cθ, nθ e della capacità di campo relativi alle diverse tessiture del suolo impiegate nelle simulazioni sono riportati in Tab. 1.
Tab. 1 - Valori dei parametri di risposta c θ, n θ e della capacità di campo per i quattro tipi di suolo considerati ([38], [5])
| Tipo di suolo | c θ | n θ | Capacità dicampo (m3 /m3) |
|---|---|---|---|
| Argilloso | 0.4 | 3 | 0.443 |
| Argilloso - limoso | 0.5 | 5 | 0.357 |
| Sabbioso - limoso | 0.6 | 7 | 0.213 |
| Sabbioso | 0.7 | 9 | 0.148 |
A parità di altri fattori, in condizioni di stress idrico tanto la produttività primaria lorda quanto l’evapotraspirazione variano in proporzione diretta al fattore di riduzione per l’umidità del suolo f θ. L’allocazione di biomassa agli apparati radicali diminuisce invece in modo curvilineo all’aumentare di tale fattore, ovvero al diminuire dello stress idrico (eqn. 3):
dove il coefficiente ftot rappresenta l’effetto cumulato sugli scambi gassosi di tutti gli altri fattori esogeni ed endogeni.
Dati di ingresso, parametri e variabili di risposta
Il modello 3-PG richiede come variabili di ingresso dati mensili di radiazione globale incidente, deficit di saturazione e temperatura media dell’aria, precipitazioni e frazione mensile di giorni di gelo. Nel presente studio sono stati utilizzati in particolare i valori riportati da Waring e McDowell ([38]), relativi al sito di Wind River, Washington, USA, caratterizzato da una copertura coetanea di Douglasia (Pseudotsuga menziesii).
La produzione primaria lorda, i flussi di allocazione e crescita, la mortalità e la caduta delle foglie, il valore dell’indice fogliare (LAI, area fogliare per unità di superficie), l’evapotraspirazione e il bilancio idrico sono stati simulati per un periodo di 160 anni con un passo giornaliero, ad eccezione della GPP per cui si è utilizzato un passo mensile.
Un elenco dei valori numerici e di alcune espressioni analitiche utilizzate nel modello viene riportato in Tab. 6. Nelle Tab. 1 e Tab. 2 sono riportati i valori di tessitura e di fertilità del terreno utilizzati nelle simulazioni.
Tab. 6 - Valore dei parametri utilizzati per le simulazioni con il modello 3-PG, relativi ad un soprassuolo di Douglasia sulla costa ovest del Nord America ([38]). (1): Landsberg & Waring ([18]); (2): Waring & McDowell ([38])
| Simbolo | Descrizione | Valore e funzioni | Unità di misura | Fonte |
|---|---|---|---|---|
| cpp | Rapporto PN /PG | 0.47 | - | 1 |
| n age | Esponente in f age | 4 | - | 1 |
| A 0.5 | Età per cui f age =0.5 | 190 | anni | |
| T min | Temperatura minima in f T | -2 | °C | 2 |
| T max | Temperatura massima in f T | 40 | °C | 2 |
| T opt | Temperatura ottimale in f T | 20 | °C | 2 |
| k g | Coefficiente in f D | 0.5 | kPa-1 | 2 |
| SLA | Area fogliare specifica | 12 | m2 kgC-1 | 2 |
| g s max | Massima conduttanza stomatica | 0.006 | m s-1 | 2 |
| g cbound | Massima conduttanza di copertura | 0.018 | m s-1 | 2 |
| γ fmax | Massimo coefficiente di mortalità fogliare tasso di caduta delle foglie |
0.2 | annomth-1 | 2 |
| c γ | Costante empirica per γ f | 15 | - | 2 |
| k γ | Esponente per γf | 0.12 | - | 2 |
| W s max | Valore inizio auto-diradamento | 11 | kgC m-2 | 2 |
| - | Densità del legno | 200 | kgC m-3 | 2 |
| - | Numero iniziale di alberi | 1500 | alberi ha-1 | 2 |
| W s | Biomassa legnosa iniziale | 0.3 | kgC m-2 | 2 |
| W f | Biomassa fogliare iniziale | 0.05 | kgC m-2 | 2 |
| W r | Biomassa radicale iniziale | 0.15 | kgC m-2 | 2 |
| - | Equazione allometrica per massa legnosa | Ws=0.0843x(diametro(cm)) 2.436 | kg | 2 |
| - | Equazione allometrica per massa fogliare | Wf=0.1484x(diametro(cm)) 2.167 | kg | 2 |
| - | Equazione allo metrica per massa radicale | Wr=0.0161x(diametro(cm)) 2.533 | kg | 2 |
Tab. 2 - Valori del coefficiente di fertilità e di efficienza di uso della luce corrispondente utilizzati nelle simulazioni ([38])
| FR | a c (kgC MJ-1 APAR) |
|---|---|
| 0.42 | 0.00237 |
| 0.45 | 0.00248 |
| 0.50 | 0.00276 |
Per caratterizzare la risposta del bosco si è focalizzata l’attenzione in particolare su quattro variabili di risposta: il valore massimo di LAI alla chiusura delle chiome (corrispondente alla fase di spessina), il volume dendrometrico di massa principale a 100 anni ed i valori medi mensili di evapotraspirazione (ET) e produttività primaria lorda (GPP) alla stessa età.
Risultati
Al fine di meglio comprendere le predizioni del modello, l’analisi degli effetti del suolo su funzionalità e crescita è stata suddivisa in due fasi, analizzando dapprima (1) la risposta a fertilità e profondità per un suolo di medio impasto, e quindi (2) la risposta alla profondità del suolo in condizioni di diversa granulometria per un dato valore di fertilità.
Effetto della fertilità e della profondità del suolo sull’accrescimento forestale
Per queste simulazioni si è considerato un suolo sabbioso limoso (c θ= 0.6, n θ= 7). Sono stati presi in esame tre livelli crescenti di fertilità del terreno (FR= 0.42/0.45/0.50) a cui corrispondono tre distinti valori del coefficiente di efficienza di uso della luce a c (Tab. 2).
Il contenuto idrico massimo del suolo q max è stato calcolato come prodotto fra capacità di campo (assunta pari a 0.213 m3/m3 per questo suolo; [5]) e la profondità del suolo, la quale è stata fatta variare da un minimo di 25 cm ad un massimo di 1.5 m.
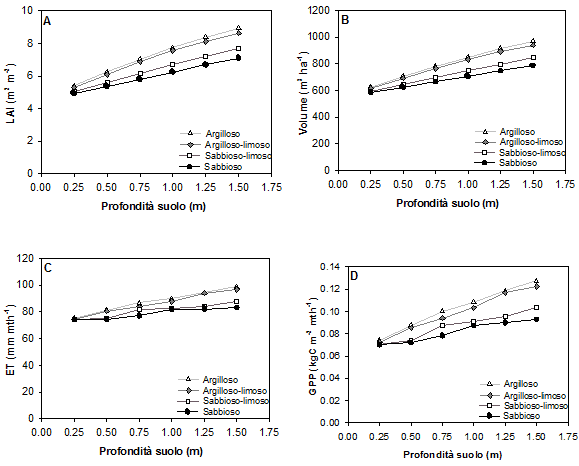
Come si osserva nei grafici di Fig. 1, la risposta di tutte le variabili funzionali considerate all’aumentare della profondità del suolo, e quindi del contenuto idrico massimo, è positiva. Passando da un suolo di profondità pari a 1 m ad un suolo di 0.5 m (ovvero per una riduzione del 50 % del contenuto idrico massimo), a parità di indice di fertilità (FR= 0.42), si osserva un decremento percentuale di GPP del 18.6 %, del 16.8 % per il LAI, del 13.7 % per il volume e dell’8.8 % per ET; in condizioni di suolo moderatamente superficiale (25 cm) la produttività primaria si riduce del 32 % rispetto a condizioni di suolo profondo. Al diminuire della profondità del suolo si osserva sempre un aumento dell’efficienza di uso dell’acqua (rapporto fra produttività primaria e quantità di acqua traspirata). Il decremento percentuale di produttività al diminuire della profondità del suolo risulta meno marcato all’aumentare della fertilità (Tab. 3).
Fig. 1 - Valore massimo del LAI (A), valore medio a 100 anni del volume (B) e valori medi mensili a 100 anni dell’evapotraspirazione (C) e della produzione primaria netta (D), in funzione della profondità del suolo per quattro differenti tipi di suolo (argilloso, argilloso limoso, sabbioso limoso e sabbioso).
Tab. 3 - Variazione percentuale di GPP, del LAI e del volume a 100 anni, passando da una profondità del suolo di 1m a quella di 0.5 m, al variare dell’indice di fertilità del terreno.
| Variabile | Indice di fertilità FR | Diminuzione percentuale (%) |
|---|---|---|
| GPP | 0.42 | 18.58 |
| 0.45 | 18.01 | |
| 0.50 | 17.08 | |
| LAI | 0.42 | 16.79 |
| 0.45 | 16.33 | |
| 0.50 | 15.57 | |
| Volume | 0.42 | 13.74 |
| 0.45 | 13.67 | |
| 0.50 | 13.56 |
Non si osserva alcun effetto della fertilità stazionale sull’evapotraspirazione dell’ecosistema (Fig. 1C) mentre a parità di profondità del suolo condizioni di maggiore fertilità hanno come conseguenza un aumento di LAI, di produttività e, quindi, di provvigione a 100 anni di età. Le variazioni percentuali dovute a variazioni nella disponibilità di nutrienti sono superiori in queste condizioni stazionali a quelle dovute alla presenza di acqua: per una profondità del suolo pari ad 1 m, passando da FR= 0.50 ad FR= 0.42 (ovvero per una riduzione del 16 % dell’indice di fertilità, variabile fra 0 e 1), si osserva un decremento del 17.7 % per GPP, del 20.7 % per il LAI e del 18.7 % per il volume (Tab. 4).
Tab. 4 - Variazione percentuale di GPP, del LAI e del volume, passando da un indice di fertilità del terreno di 0.50 ad uno di 0.42, al variare della profondità del terreno.
| Variabile | Profondità del terreno (m) | Diminuzione percentuale (%) |
|---|---|---|
| GPP | 0.50 | 19.20 |
| 1 | 17.71 | |
| LAI | 0.50 | 21.81 |
| 1 | 20.67 | |
| Volume | 0.50 | 18.87 |
| 1 | 18.71 |
Confrontando la risposta del modello ad una variazione del 25% sia del contenuto idrico massimo sia della fertilità, si può osservare come il modello sia molto più sensibile a questo secondo parametro date le condizioni ambientali riportate in Tab. 5.
Tab. 5 - Sensitività del modello a variazioni del 25% della profondità del suolo (valore di riferimento 1m) e della fertilità (valore di riferimento FR = 0.45) per: GPP, LAI, ET e volume.
| Variabile | Risposta alla variazione della profondità suolo (%) | Risposta alla variazione della fertilità (%) |
|---|---|---|
| GPP | 4.43 | 29.51 |
| LAI | 7.95 | 34.37 |
| Volume | 6.63 | 30.87 |
| ET | 1.54 | - |
Effetto delle caratteristiche fisiche del terreno e della profondità del suolo sull’accrescimento forestale
Avendo analizzato in dettaglio gli effetti della fertilità, si è deciso di assumere nel resto dell’analisi un valore costante di fertilità (FR= 0.45; corrispondente ad un valore di efficienza di uso della luce a c= 0.00248 kgC MJ-1 PAR).
Due sono i meccanismi di azione della tessitura del suolo presi in considerazione nel modello: (i) attraverso i suoi effetti sulla capacità di campo e quindi sulla quantità di acqua disponibile per le piante nel corso dell’anno e (ii) per le differenze nella risposta degli scambi gassosi al contenuto idrico del suolo (eqn. 2). Al fine di discriminare fra i due meccanismi, la capacità di ritenzione idrica totale (data dal prodotto della capacità di campo per la profondità del suolo) è stata modulata nelle simulazioni sia assumendo diverse tessiture del suolo (e quindi diversi valori di capacità di campo; Tab. 1), sia variando la profondità del suolo stesso. Sono stati a tal fine considerati quattro tipi di suolo (argilloso, argilloso limoso, sabbioso limoso, sabbioso; Tab. 1) e una profondità del suolo variabile fra 0.25 e 1.5 m.
I risultati delle simulazioni sono stati espressi in relazione sia alla profondità (Fig. 2) che al contenuto idrico massimo del suolo (Fig. 3).
Fig. 2 - Valore massimo del LAI (A), valore medio a 100 anni del volume (B) e valori medi mensili a 100 anni dell’evapotraspirazione (C) e della produzione primaria netta (D), in funzione della profondità del suolo e della fertilità, per un suolo sabbioso-limoso.
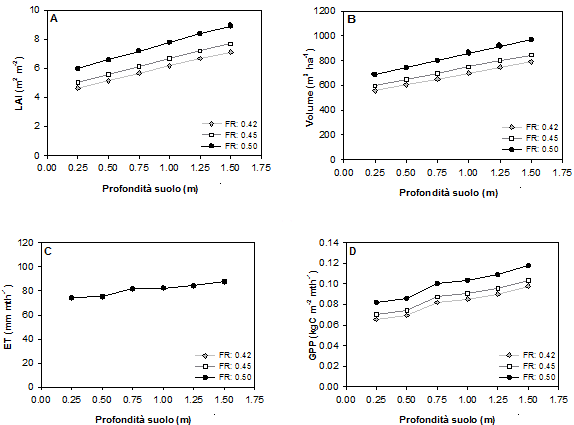
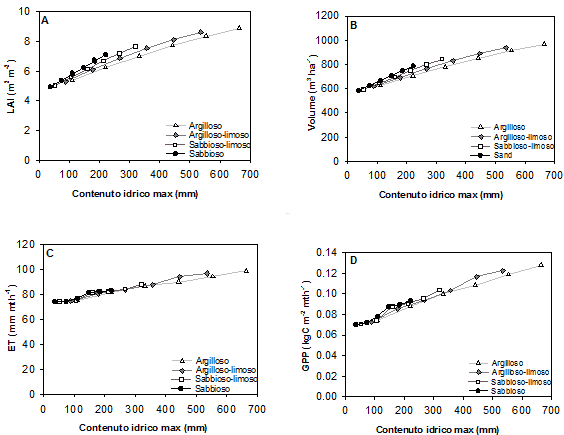
Fig. 3 - Valore massimo del LAI (A), valore medio a 100 anni del volume (B) e valori medi mensili a 100 anni dell’evapotraspirazione (C) e della produzione primaria lorda (D), in funzione del contenuto idrico massimo nel suolo per quattro differenti tipi di suolo (argilloso, argilloso limoso, sabbioso limoso e sabbioso).
Per tutte le variabili di risposta considerate la funzionalità del bosco appare essere favorita da tessiture fini, mentre i risultati peggiori sono costantemente osservati nel caso di suoli sabbiosi per ogni valore di profondità (Fig. 2). Le differenze legate alla tessitura si riducono per suoli poco profondi. Assumendo una profondità del suolo di 1 m si osserva, ad esempio, che il valore del LAI per le sabbie è inferiore del 19.11 % rispetto a quello nel caso dell’argilla; per il volume tale differenza è del 17%, per ET del 9.06% e 19.2% per GPP. Questa differenza si riduce, alla profondità di 25 cm, all’8.43% per il LAI, al 6.57% per il volume, al 1.56% per ET ed infine al 5.6% per GPP.
L’effetto della tessitura appare però legato in larga misura alla diversa capacità di ritenzione idrica dei suoli di differente granulometria, massima nei suoli argillosi e minima nei suoli sabbiosi. Una volta compensato l’effetto dovuto alla capacità di campo, esprimendo cioè i risultati in termini non di profondità del suolo ma di contenuto idrico massimo, si osserva infatti che a parità di quest’ultimo i valori di crescita e funzionalità non differiscono significativamente fra i diversi suoli (Fig. 3). Si osservano anzi valori di crescita maggiori per i terreni sabbiosi rispetto a quelli argillosi, con una differenza dell’11 % in corrispondenza di un contenuto idrico massimo di 200 mm circa. La differenza è invece quasi trascurabile per quanto riguarda evapotraspirazione e produttività primaria lorda.
Discussione
L’esigenza di stimare su larga scala la produttività e la capacità di fissazione ed immagazzinamento del carbonio atmosferico da parte degli ecosistemi forestali ci pone di fronte al problema di disporre di un modello affidabile che richieda al contempo un basso numero di parametri di facile misurabilità o reperibilità. Il modello 3-PG, sviluppato da Landsberg & Waring ([18]), è un modello a base funzionale che sembra rispondere a questi requisiti, grazie alla sua semplice struttura generale, focalizzata sui soli processi fondamentali, ed al riconoscimento di importanti convergenze funzionali fra differenti tipi vegetali. Il modello è stato ampiamente testato e sembra ben catturare la risposta ai principali fattori ambientali di funzionalità e crescita del bosco ([19], [34], [20]).
I modelli sono per definizione delle rappresentazioni semplificate e schematiche della realtà. Per tale motivo rischiano ovviamente di non cogliere le proprietà essenziali dell’ecosistema. Questo rischio è tanto più concreto nel caso di un semplice modello quale 3-PG. Anche considerando solo la risposta alla disponibilità idrica del suolo, infatti, il modello trascura numerosi dettagli evidenziati dalla ricerca idrologica. Nella realtà tessitura, ritenzione e capacità idrica, contenuto in nutrienti e profondità del suolo non sono fattori scollegati tra di loro, così come invece vengono trattati dal modello. Relativamente alla profondità del suolo, ad esempio, è noto come il parametro di reale interesse non sia tanto la profondità del suolo minerale, più facilmente determinabile, quanto piuttosto la profondità del suolo esplorato dalle radici, la quale varia in funzione della specie, dello stato di sviluppo delle piante e della tessitura del suolo stesso. In suoli con maggiore porosità si creano infatti condizioni di aerazione favorevoli ad un maggiore sviluppo in profondità dell’apparato radicale, che può così sfruttare meglio l’acqua e i nutrienti presenti nel suolo ([12], [36]). Inoltre le caratteristiche idriche del suolo dipendono non solo dalla tessitura, ma anche dalla quantità di sostanza organica ivi presente, un aspetto particolarmente importante nei suoli forestali ([36]). Nessuno di questi aspetti viene considerato nel modello 3-PG.
Anche l’applicazione di un modello semplificato come 3-PG, d’altra parte, rischia di risultare limitata, più che dalla conoscenza dei parametri funzionali della specie, dalla disponibilità di informazioni precise sulle caratteristiche stazionali. Se infatti variabili d’ingresso quali radiazione, temperatura e precipitazioni sono disponibili in forma spazializzata per tutto il territorio nazionale, altre informazioni, relative in particolare alla natura dei suoli ed alla loro profondità, sono di più difficile reperimento. Da qui la ricerca di ulteriori elementi di semplificazione attraverso un’analisi di sensitività ai fattori legati al suolo.
Da quanto detto, è quindi evidente la necessità di meglio comprendere quale sia l’effetto delle semplificazioni introdotte e in che misura sia possibile trattare i diversi processi in forma aggregata.
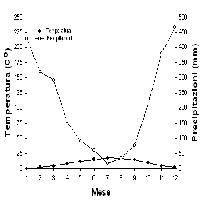
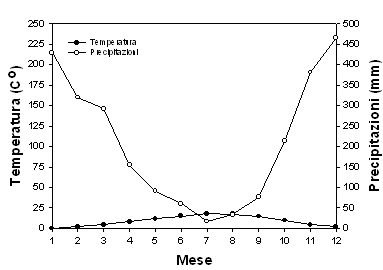
Dall’analisi di sensitività alle variazioni di fertilità e contenuto idrico del suolo è emersa una risposta positiva del modello a questi parametri, più marcata nel primo caso che nel secondo: per differenze del 25 % della fertilità del terreno la variazione di produttività primaria lorda si mantiene intorno al 30 %; in condizioni di suolo moderatamente superficiale (25 cm) tale variazione raggiunge il 32 % rispetto a condizioni di suolo profondo. È nota l’importanza della disponibilità di nutrienti per la crescita dei soprassuoli forestali ([27]), mentre è spesso trascurata l’influenza della profondità dei suoli, anche a causa della sua difficile definizione. Occorre sottolineare come la presente analisi abbia fatto riferimento a condizioni di buona disponibilità idrica, con due soli mesi di aridità estiva e temperature piuttosto miti (Fig. 4), tali da permettere a questa specie sempreverde di fotosintetizzare attivamente nei mesi autunnali e primaverili ([13]). È verosimile che la sensitività alla profondità del suolo risulti maggiore in presenza di un periodo secco più prolungato, quale spesso si riscontra nelle Regioni a clima mediterraneo del Centro e Sud Italia.
Fig. 4 - Distribuzione stagionale di temperature e precipitazioni dal 1900-1977 a Wind River, Washington ([38]).
Ciò sembrerebbe essere in accordo con l’osservazione ([9]) di un più stretto rapporto della produttività della specie nell’Italia centrale con i fattori climatici direttamente correlati al bilancio idrico che con i fattori chimici relativi al suolo o alla topografia. Se possiamo però dire che in questi climi (verosimilmente rappresentativi di gran parte dell’Appennino italiano) la presenza di suoli superficiali non compromette drasticamente la crescita delle specie, gli effetti della fertilità chimica appaiono invece di tutto rilievo, imponendo di disporre di stime attente di questo parametro o, in mancanza di queste, di stime dello stato nutrizionale della pianta da dati tele-rilevati ([23], [16]).
Molto limitata è invece la risposta della produttività forestale alla tessitura del suolo, a parità di grado di fertilità e di disponibilità idrica totale.
Nella presente analisi è stato messo in evidenza come due siano in realtà i meccanismi di azione della tessitura del suolo: (i) attraverso i suoi effetti su capacità di ritenzione idrica totale dell’intero profilo del suolo e quindi sulla quantità di acqua disponibile per le piante nel corso dell’anno, avendo i suoli sabbiosi tipicamente una capacità di campo più bassa dei suoli a porosità fine, e (ii) per le differenze nella risposta degli scambi gassosi al contenuto idrico del suolo, legate al diverso rapporto fra contenuto idrico e potenziale idrico del suolo. Vale la pena sottolineare come solo questo secondo aspetto fosse stato esplicitamente menzionato nella descrizione del modello 3-PG. I risultati dell’analisi dimostrano come funzionalità e produttività del bosco paiano essere favorite da tessiture fini, mentre i risultati peggiori sono costantemente osservati nel caso di suoli sabbiosi per tutti i valori di profondità del suolo (Fig. 2). Questa predizione del modello è in ottimo accordo sia con i risultati della sperimentazione agricola e forestale ([35]) sia con le più recenti analisi ecofisiologiche di dettaglio ([12]). Tali differenze legate alla tessitura si riducono peraltro nel caso di suoli poco profondi.
L’effetto della tessitura sembra legato però in larga misura alla diversa capacità di campo dei suoli a differente granulometria: massima nei suoli argillosi e minima nei suoli sabbiosi. Una volta infatti compensato l’effetto dovuto alla capacità di campo, esprimendo cioè i risultati in termini non di profondità del suolo ma di contenuto idrico massimo, i valori di crescita e funzionalità non differiscono sostanzialmente fra i diversi suoli (Fig. 3).
In fase predittiva sembrerebbe quindi possibile trascurare i parametri legati alla tessitura del suolo e riferirsi al solo quantitativo massimo di acqua disponibile nell’intero profilo a capacità di campo senza commettere errori significativi.
Le simulazioni condotte con il modello hanno anche sottolineato l’assenza di risposta dell’evapotraspirazione alla variazione del grado di fertilità del terreno. Questo risultato sembra confermato dalle osservazioni di Ripullone et al. ([28]), che non hanno riscontrato alcun effetto della fertilizzazione sulla traspirazione o sulla conduttanza stomatica. Una maggiore disponibilità di nutrienti, stimolando la produttività primaria senza determinare un parallelo aumento della traspirazione, parrebbe pertanto indurre una maggiore efficienza di uso dell’acqua, come già riportato da Guehl et al. ([11]) e da Ripullone et al. ([28]).
È questo un risultato di notevole importanza per la selvicoltura in area mediterranea, che dovrebbe pertanto mirare ad aumentare la fertilità chimica dei suoli attraverso un’attenta gestione della sostanza organica.
Numerosi studi evidenziano una correlazione tra quantitativo idrico disponibile e concentrazione di nutrienti nel suolo, sottolineando come la produttività aumenti in risposta alla fertilizzazione solo se l’apparato radicale dispone del quantitativo di acqua necessario per poterne trarre giovamento ([33], [26], [3]). Un aumento della concentrazione di nutrienti nel suolo causa, infatti, un aumento di produttività primaria solo nel primo anno (o al più nei primi anni) di trattamento, per poi tornare ai tassi di crescita simili a quelli del controllo. Un’adeguata irrigazione prolunga questo effetto nel tempo. Questo comportamento del sistema non viene evidenziato dall’analisi eseguita con il modello 3-PG in quanto, ancora una volta, il modello descrive i fattori influenzanti la crescita arborea come separati. Se da un lato è facile operare considerando separatamente i vari fattori ambientali, dall’altro è però molto difficile valutare e predire l’effetto delle loro combinazioni sulla crescita della pianta: combinazioni differenti possono avere effetti simili sulle piante a causa di meccanismi di compensazione o altri processi ancora non pienamente compresi.
Concludendo è possibile affermare che il modello 3-PG risulta sensibile sia a variazioni dell’indice di fertilità che al contenuto idrico massimo. Trascurando questi due fattori si ottengono errori nella stima della produttività primaria dell’ordine del 30-32%.
References
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar