Changes in rainfall patterns in Mediterranean ecosystems: the MIND project
Forest@ - Journal of Silviculture and Forest Ecology, Volume 4, Pages 460-468 (2007)
doi: https://doi.org/10.3832/efor0493-0040460
Published: Dec 20, 2007 - Copyright © 2007 SISEF
Research Articles
Guest Editors: Joint Session SITE-SISEF (Viterbo 2006)
« Forests and Climate: 10 years of Research in Italy »
Collection/Special Issue: Paolo De Angelis, Riccardo Valentini
Abstract
Will Mediterranean terrestrial ecosystems be affected by the expected changes in precipitation regimes? If so, by how much and in which direction? These questions are at the basis of the research performed in context of the EU MIND project, whose key objectives were: i) to investigate the potential effects of increasing drought on Mediterranean terrestrial ecosystems at the process, ecosystem and regional scales and ii) to assess ecosystem vulnerability to changes in rainfall patterns. A network of experimental study sites has been created in Portugal, Spain, France and Italy, where field manipulations alter the amount of water available to the ecosystem. The most up-to-date methods of ecophysiology, micrometeorology, soil ecology and remote sensing have been used to elucidate the mechanisms that regulate the response of vegetation and soil to changes in water availability. This information is providing the basis for the implementation and validation of simulation models capable of predicting the drought response of Mediterranean terrestrial ecosystems, and their vulnerability to future climate change, on a larger scale. The out-coming results are elucidating how water availability affects plant ecophysiological processes, the dynamics of soil carbon and the overall exchange of mass and energy between the land and the atmosphere. This paper focuses on some of the important, yet preliminary, results on C and energy fluxes that have been obtained at the large scale troughfall manipulation experiment (Tolfa, Italy), in a forest dominated by Arbutus unedo L.
Keywords
Climate change, Rainfall, Mediterranean ecosystems, Carbon stock, Eddy covariance, MIND project
Introduzione
Il clima nell’area mediterranea è caratterizzato da inverni freddi e piovosi ed estati calde e secche durante le quali si registrano numerose limitazioni nella disponibilità idrica ([14]). La vegetazione dominante in questa area è rappresentata da sclerofille sempreverdi che si sono adattate a sopravvivere durante il periodo di siccità estiva. Infatti, la struttura e la fisiologia fogliare di questo gruppo di specie consente loro di minimizzare le perdite d’acqua ed i danni termici. Gli apparati radicali in genere profondi consentono loro di accedere ad altrettanto profonde risorse idriche e di mantenersi fisiologicamente attive durante il periodo secco. Tuttavia, la maggior parte dell’accrescimento avviene nel corso del periodo primaverile ed autunnale quando le condizioni di temperatura e rifornimento idrico sono ottimali ([5]) e, nonostante si tratti di specie sempreverdi, la struttura e dimensione della canopy sono fortemente influenzate dalla disponibilità idrica estiva ([21]). Pertanto, la temperatura e le precipitazioni svolgono un ruolo importante nel determinare la produttività primaria e la struttura degli ecosistemi mediterranei. Non sorprende che i recenti cambiamenti climatici hanno avuto, e stanno avendo, un severo impatto su questo tipo di ecosistemi ([19]). Nell’ultimo secolo è stato registrato a livello globale un aumento della temperatura media dell’aria di circa 0.5°C dovuto ad un aumento dei gas serra tra cui la CO2 ([13]). I modelli di circolazione globale indicano che alcune regioni saranno soggette ad un aumento nelle precipitazioni, mentre altre subiranno periodi di siccità prolungati come conseguenza di una riduzione delle precipitazioni o di un aumento nella domanda evaporativa nei climi più caldi ([9], [18]); questi effetti saranno molto importanti soprattutto a livello di regione mediterranea ([13]). L’aumento della temperatura insieme alla diminuzione delle precipitazioni espongono la vegetazione ad un prolungato periodo di stress idrico che potenzialmente ha notevoli effetti sulle diverse componenti che costituiscono la produttività netta ecosistemica (NEP). Per esempio, un prolungato periodo di siccità può ridurre la produttività primaria netta (NPP), modificando l’efficienza traspirativa, ossia la quantità di carbonio prodotta per unità di volume d’acqua traspirato ([19]). Inoltre, la temperatura del suolo e l’umidità sono i principali fattori che influenzano la respirazione del suolo (SR): numerosi studi hanno verificato l’esistenza di una forte correlazione tra SR ed umidità del suolo in ambienti mediterranei ([28], [20], [22], [30]). In queste situazioni, a causa dello stress idrico, la SR mostra un andamento stagionale opposto a quello che sarebbe possibile prevedere facendo riferimento unicamente alla temperatura del suolo ([16], [31], [4]).
Il presente lavoro è stato condotto nell’ambito del progetto “Mediterranean Ecosystems under increasing Drought” (MIND, EU V Programma quadro) il cui scopo è quello di investigare gli effetti di lungo periodo derivanti da un aumento della siccità in ecosistemi mediterranei a diverse scale di misura (cioè processo, ecosistema e scala regionale) attraverso la quantificazione degli scambi di carbonio, acqua ed energia tra biosfera ed atmosfera a scale temporali e spaziali diverse. Il progetto ha previsto la messa a punto di esperimenti di manipolazione delle piogge in Spagna, Portogallo, Francia ed Italia. Nel presente lavoro, sono riportati alcuni dei risultati preliminari derivanti da due anni di misure condotte nel sito italiano di Tolfa Allumiere (Roma) in un popolamento dominato da corbezzolo (Arbutus unedo L.). In particolare è stata studiata la risposta in termini di scambio ecosistemico netto (NEE), respirazione del suolo (RS) ed evapotraspirazione (ET) a modificazioni nel regime pluviometrico.
Materiali e metodi
Sito di studio
Il sito di studio è situato a Tolfa Allumiere nel centro Italia (42°11’ N 11°56’E) ad una quota di 180 m s.l.m. L’area è caratterizzata da un clima mediterraneo con una precipitazione annua di 650 mm ed una temperatura media annua di 15°C. Il soprassuolo è un ceduo di circa 6 ha dominato da Arbutus unedo L. (65%) ed Erica arborea L. (13%) con sporadica presenza di Fraxinus ornus L., Quercus ilex L. e Quercus poubescens L. L’altezza media della canopy è 7 m e la densità del soprassuolo è di 4923 fusti ha-1. Una campagna di misura condotta prima dell’inizio dell’esperimento ha consentito di misurare un’area basimetrica totale di 19.57 m2 ha-1 ed una biomassa totale (fusto e rami) di 34.08 Mg ha-1. I suoli sono classificabili come Andosols derivati da depositi di origine vulcanica, sono poco profondi (mediamente 31 cm), franco-sabbiosi con un pH acido (pH=4).
Il sistema sperimentale
All’interno dell’area di studio, sono state identificate due parcelle sperimentali (DRY e WET) di ampie dimensioni (1 ha ciascuna; large plot) su cui è stato condotto l’esperimento di manipolazione della pioggia su larga scala attraverso la messa in opera di un sistema di intercettazione della precipitazione sottochioma costituito da dreni metallici coprenti il 20% della superficie del plot DRY e un sistema di irrigazione a goccia nel plot WET. In entrambe le aree, i flussi di carbonio, vapore acqueo ed energia sono stati misurati in maniera continua utilizzando due torri di Eddy Covariance (EC), posizionate ciascuna al centro di una delle due parcelle. Inoltre sono state individuate sei aree di monitoraggio intensivo (IMP) di 8 x 12 m accoppiate per trattamento (cioè DRY e WET) e replicate in tre blocchi. Nelle IMP DRY, la precipitazione sottochioma è stata intercettata attraverso dei dreni metallici sospesi a 1.8 m sopra il suolo nella parte centrale del plot e ad 1.6 m nella parte perimetrale e connessi a due grondaie per allontanare l’acqua intercettata al di fuori dell’IPM. Così come nelle aree di 1 ha, i dreni, che corrono a circa 10 cm dal suolo, coprono un’area pari al 20% della superficie di ciascun plot e sono pertanto in grado di intercettare il 20% della precipitazione sottochioma.
Il trattamento WET è stato realizzato mediante l’uso di irrigatori tipo sprinkler nelle IMP ed a pioggia nei large plot, distribuiti uniformemente al fine di simulare efficacemente gli eventi di pioggia. L’acqua necessaria per le irrigazioni è stata ottenuta da un pozzo artesiano ed ogni irrigazione è stata controllata manualmente da un operatore. Le irrigazioni sono state effettuate durante le estati del 2004 e del 2005, quando la vegetazione si trovava in condizioni di stress in modo da mantenere l’umidità del suolo al di sopra di una soglia del 10%, misurata quale soglia di stress per il corbezzolo nel sito di studio (Magnani, comm. pers.). In Tab. 1 sono riportate la distribuzione temporale delle irrigazioni effettuate.
Tab. 1 - Distribuzione delle irrigazioni nel trattamento WET durante il 2004 ed il 2005.
| Anno | Giorno | mm |
|---|---|---|
| 2004 | 209 215 237 245 254 |
9 18 18 8 20 |
| 2005 | 95 152 160 166 186 196 200 206 210 215 223 235 256 |
20 30 20 40 30 20 30 30 30 30 30 30 30 |
In ogni IMP, è stata misurata la temperatura del suolo utilizzando 3 termocoppie posizionate a 2 cm e 3 termocoppie posizionate a 12 cm in modo da misurare, rispettivamente, la temperatura dell’orizzonte organico e di quello minerale; l’umidità del suolo è stata misurata nel 2004 usando 6 sensori dielettrici di umidità (ECHO Mod. EC-20, Decagon Devices, Inc.) per ogni IMP che nel 2005, a causa di un guasto al sistema, sono stati sostituiti con un TDR per IMP (CS616, Campbell Scientific, Inc.). All’esterno del bosco è stata sistemata una stazione meteo per misurare la radiazione globale incidente, le precipitazioni, la temperatura e l’umidità dell’aria. Tutte le misure sono state acquisite usando un datalogger per ogni stazione (CR10X, Campbell Scientific, inc.) e mediate alla mezz’ora.
I sistemi di misura sono stati resi completamente operativi a partire da aprile 2004.
Evapotraspirazione
Il flusso istantaneo xilematico (Js) è stato misurato in maniera continua nell’estate 2004 e 2005 mediante sensori di dissipazione termica costruiti seguendo le indicazioni di Sakuratani ([26], [27]) e Baker & van Bavel ([2]). Sono stati selezionati 30 fusti di corbezzolo distribuiti nei sei IMP e le misure sono state mediate alla mezz’ora. I sensori sono stati installati su fusti con un diametro tra 17 e 30 mm ed al di sopra di qualunque ramo in maniera che la misura di flusso fosse rappresentativa del fusto. I valori misurati sono stati riportati a livello di soprassuolo usando valori di spessore del sap wood desunti dalla letteratura ([17]).
Respirazione del suolo
La respirazione del suolo è stata monitorata in continuo ogni quattro ore durante tutto il 2004 ed il 2005, attraverso due sistemi di misura con 6 camere ciascuno, operanti in contemporanea ciascuno in uno dei due trattamenti (cioè dry e wet), e che sono stati fatti ruotare da un blocco di IMP all’altro ogni due settimane. Ogni camera è costituita da un collare metallico (20 cm di diametro, 8 cm di altezza) aperto alle due estremità e dotato di un coperchio mobile automatizzato. Per minimizzare modificazioni dell’ambiente all’interno di ciascuna camera, il coperchio è rimasto aperto tra le sessioni di misura e le camere sono state posizionate in modo che il coperchio aperto fosse orientato a Nord. L’aria è prelevata dalla camera attraverso il coperchio ed è restituita alla camera da un anello posto al di sopra della superficie del suolo. Ogni camera è collegata al sistema di misura tramite tubi in PVC di 10 m di lunghezza e diametro interno di 4 mm. Il principio di misura è analogo a quello usato dal Li-Cor 6400 (LiCor Biosciences, Lincoln, Nebraska): il tasso di crescita della concentrazione di CO2 (dc/dt) all’interno della camera è misurato in un intervallo [CO2]ambiente Δ e il valore finale di flusso è calcolato utilizzando la pendenza della regressione lineare tra dc/dt e [CO2] all’interno della camera. Per una descrizione dettagliata del sistema si rimanda a Inglima ([11]). I dati misurati sono stati controllati utilizzando Stata 7.0 Intercooled Version: per ogni camera sono state plottate tutte le concentrazioni di CO2 misurate in un giorno rispetto al tempo. Quando anche una sola rampa sembrava non essere corretta a causa di un malfunzionamento del sistema, i dati di tutta la giornata della corrispondente camera sono stati eliminati. Le sessioni in cui le camere funzionanti erano meno di tre per plot sono state scartate. La media delle misure effettuate sulle camere funzionanti (sempre più di tre) è stata presa come valore della SR per il plot nella sessione considerata. I dati così ottenuti per i trattamenti DRY e WET sono stati confrontati e solo quando le misure effettuate in entrambi i trattamenti avevano superato il quality check sono state prese in considerazione per le successive analisi.
Scambio ecosistemico netto (NEE)
All’interno delle due parcelle di 1 ha sono state posizionate due torri di eddy covariance al fine di misurare i flussi di CO2 (NEE), calore sensibile (LE) e calore latente (H) tra l’atmosfera e l’ecosistema. Ciascuna torre è equipaggiata con un analizzatore ad infrarossi LI-7000 (Li-Cor Inc. Nebraska, USA) per misurare le concentrazioni di CO2 ed H2 O ed un anemometro Gill3 (Gill Instruments Ltd., Lymington, UK) per la determinazione delle tre componenti della velocità del vento. La frequenza di campionamento è stata di 20 Hz ed i flussi sono poi stati calcolati con passo alla mezz’ora. Gli algoritmi usati per la determinazione dei flussi e dei parametri di turbolenza sono stati forniti nell’ambito del progetto CarboEurope IP (⇒ http://www.carboeurope.org). In corrispondenza di ogni torre, sono anche stati misurati altri parametri quali la radiazione fotosinteticamente attiva (PAR), la radiazione netta (Rn) la temperatura del suolo, la temperatura dell’aria e l’umidità relativa. Sia il sistema di acquisizione eddy che quello meteo hanno lavorato in maniera continuativa dalla primavera del 2004 a tutto il 2005, ad eccezione di brevi periodi a causa di problemi tecnici ai sistemi. I flussi notturni sono stati corretti adottando una soglia di u star pari a 0.3 m s-1.
Risultati
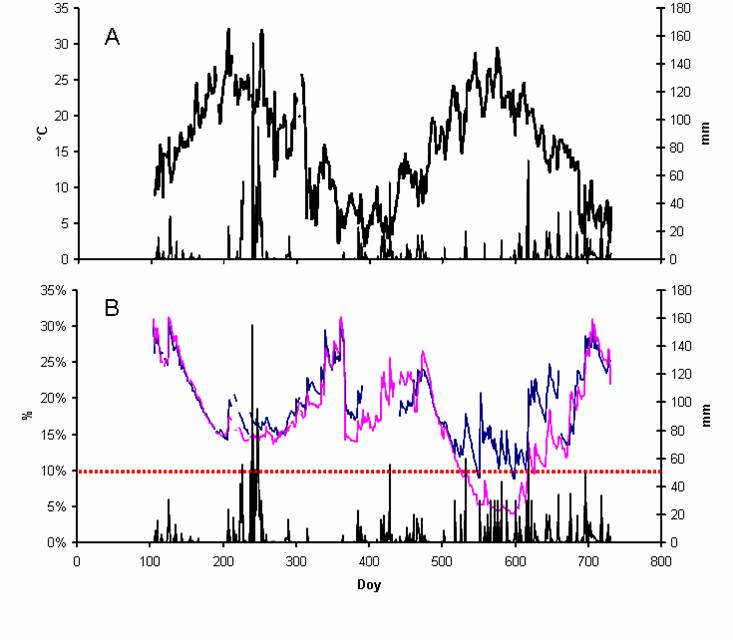
La temperatura media annua nel corso del 2004 è stata di 18°C con una minima assoluta di 2°C ed una massima assoluta di 32°C, nel 2005 la media è stata di 14°C con una minima di 2°C ed una massima di 29°C. Il 2004 è stato leggermente più piovoso del 2005 (978 contro 938 mm), ma si è registrata un’anomalia nella distribuzione delle precipitazioni. Infatti, nel primo anno le precipitazioni sono state concentrate nel periodo estivo (luglio-agosto), mentre nel 2005 sono state distribuite nel periodo primaverile ed autunnale come è tipico dell’ambiente mediterraneo (Fig. 1A). Pertanto, nel 2004 non si è registrata una differenza tra i due trattamenti in termini di umidità del suolo (SWC, Fig. 1B); nel 2005, invece, grazie alle irrigazioni (370 mm, Tab. 1) la SWC del WET non è mai scesa al di sotto della soglia prefissata del 10%, mentre il DRY ha subito una graduale riduzione della SWC fino ad un minimo estivo inferiore al 5%, per poi aumentare in seguito alle prime piogge autunnali.
Fig. 1 - Andamento meteo ed umidità del suolo nel 2004 e nel 2005: a) temperatura media dell’aria (linea continua) e precipitazioni (barre verticali); b) umidità media nel suolo nel trattamento WET (blu) e DRY (rosa) e somma delle precipitazioni ed irrigazioni (barre verticali) (B). La linea orizzontale tratteggiata indica la soglia di umidità del suolo del 10%.
Evapotraspirazione
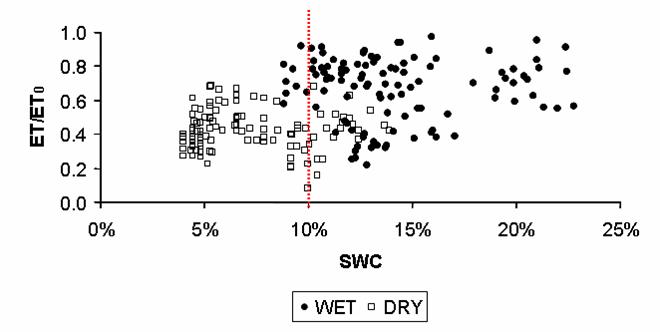
La traspirazione dell’Arbutus unedo mostra una variazione giornaliera determinate dalla radiazione solare e dall’umidità del suolo (dati non riportati). La cumulata durante il periodo di studio non mostra alcuna differenza nel 2004 (291 mm nel WET, 289 mm nel DRY - Fig. 2A) mentre mostra una notevole differenza nel 2005 (388 mm nel WET, 253 mm nel DRY - Fig. 2B) a cominciare da quando l’umidità del suolo è scesa al di sotto del 10% nel trattamento DRY. Infatti, il rapporto tra traspirazione giornaliera misurata (ET) e quella potenziale (ET0) calcolata nel 2005 secondo l’equazione di Penman & Monteith, si dimezza al di sotto del valore soglia di SWC pari al 10% (Fig. 3).
Fig. 2 - Cumulata della traspirazione della canopy nei trattamenti WET (linea continua blu) e DRY (linea continua rosa) ed umidità (linee tratteggiate, stessi colori) durante il periodo estivo del 2004 (grafico A) e 2005 (grafico B). La linea tratteggiata orizzontale indica la soglia si umidità del suolo pari al 10%.
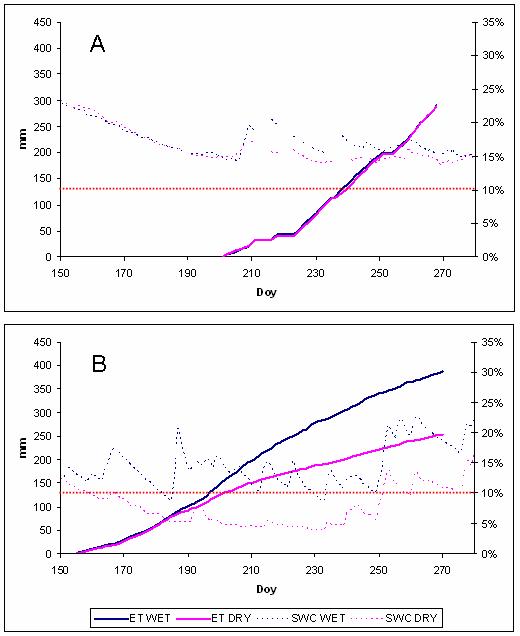
Fig. 3 - Andamento del rapporto tra traspirazione (ET) misurata ed evapo-traspirazione potenziale (ET0) rispetto all’umidità del suolo (SWC, % v/v) nel 2005. I valori si riferiscono alla somma giornaliera. La linea tratteggiata verticale indica la soglia del 10%.
Respirazione del suolo
La temperatura è risultato il principale fattore di regolazione della respirazione del suolo nella maggior parte del periodo in cui è stato condotto l’esperimento. Tuttavia, durante l’estate il suo controllo sulla respirazione è stato limitato a causa di limitazioni nella disponibilità idrica soprattutto nel trattamento DRY (dati non riportati). La respirazione media mensile è significativamente diversa nei due trattamenti nel periodo estivo sia nel 2004 che nel 2005 (Tab. 2), ma se si considera la cumulata annua il trattamento DRY presenta una respirazione leggermente inferiore anche se la differenza non è statisticamente significativa. Infatti, nel periodo autunnale, in seguito alle piogge, il suolo nel trattamento DRY respira in misura uguale o addirittura di più che nel WET.
Tab. 2 - Respirazione media giornaliera ± deviazione standard (μmol m-2 s-1) nei due trattamenti DRY e WET durante il 2004 ed il 2005. I valori statisticamente diversi tra i due trattamenti (P < 0.05) sono evidenziati con l’asterisco.
| Anno | Mese | Respirazione media giornaliera | |
|---|---|---|---|
| (μmol m-2 s-1) | |||
| DRY | WET | ||
| 2004 | 4 | 3.74±0.46 | 4.79±0.75 * |
| 5 | 4.14±0.30 | 4.31±0.44 | |
| 6 | 3.43±0.21 | 4.15±0.39 * | |
| 7 | 3.61±0.18 | 4.06±0.24 * | |
| 8 | 2.55±0.22 | 4.60±0.26 * | |
| 9 | 2.88±0.22 | 3.82±0.27 * | |
| 10 | 3.58±0.20 | 3.67±0.19 | |
| 11 | 3.73±0.32 | 3.99±0.28 | |
| 12 | 3.72±0.29 | 3.69±0.29 | |
| 2005 | 1 | 2.04±0.19 | 2.31±0.15 * |
| 2 | 2.12±0.14 * | 1.60±0.13 | |
| 3 | 2.27±0.18 | 2.07±0.20 | |
| 4 | 3.90±0.48 | 3.55±0.28 | |
| 5 | 4.66±0.30 | 5.09±0.44 | |
| 6 | 4.65±0.23 | 5.15±0.32 | |
| 7 | 3.40±0.20 | 4.89±0.59 * | |
| 8 | 3.33±0.36 | 4.69±0.57 * | |
| 9 | 5.13±0.52 | 4.75±0.78 | |
| 10 | 6.54±0.31 * | 5.33±0.42 | |
| 11 | 3.67±0.29 | 3.47±0.32 | |
| 12 | 2.17±0.14 | 2.02±0.15 | |
Scambio ecosistemico netto
A causa del malfunzionamento dei sistemi eddy in diversi periodi nel corso del 2004, i dati relativi a questo anno non sono riportati nel presente lavoro. Al contrario, nel 2005, le irrigazioni sono state condotte regolarmente nel corso dell’intero periodo estivo (Tab. 1) e non si sono avuti prolungati malfunzionamenti del sistema. In seguito alle irrigazione si è sempre registrato un forte aumento della respirazione ecosistemica (Reco) giornaliera misurata nella parcella WET rispetto alla DRY. Ciò sembra essere confermato dal fatto che il rapporto tra produttività primaria lorda (GPP) e Reco è significativamente diverso nel periodo precedente alle irrigazioni (doy 96-110), mentre cresce gradatamente nel periodo estivo quando sono state condotte le irrigazioni (doy 226-240). Tale comportamento è meglio messo in evidenza se si considerano le cumulate di NEE e GPP nel corso del 2005: infatti la parcella WET ha registrato uno scambio ecosistemico netto significativamente inferiore a quello misurato nella DRY (-492 gC m-2 nel WET contro -255 gC m-2 nel DRY) e la differenza tra le cumulate si è cominciata a verificare a partire dal giorno 150 quando l’umidità del DRY è scesa al di sotto del 10% (Tab. 3). Questi dati sembrano confermare l’ipotesi di un aumento della respirazione ecosistemica e della attività fotosintetica in seguito ad una maggiore disponibilità idrica.
Tab. 3 - Cumulate annue della produzione primaria lorda (GPP), respirazione ecosistemica (RECO) e scambio ecosistemico netto (NEE).
| 2005 | |||
|---|---|---|---|
| Parametro | GPP | RECO | NEE |
| gC m-2 anno-1 | gC m-2 anno-1 | gC m-2 anno-1 | |
| DRY | 1243 | 751 | -492 |
| WET | 1565 | 1310 | -255 |
Discussione
Negli ambienti mediterranei l’accrescimento delle specie è principalmente limitato dalla disponibilità idrica e le precipitazioni esercitano un ruolo fondamentale per quanto riguarda l’attività fotosintetica e gli scambi gassosi tra vegetazione ed atmosfera. Nelle specie tipiche di questi ambienti, la chiusura degli stomi rappresenta un meccanismo per far fronte ai deficit idrici giornalieri e stagionali ([25], [8]): presumibilmente, la riduzione del flusso traspirativo registrata nelle parcelle DRY nell’estate 2005 è causata da un controllo sulle perdite idriche esercitato dagli stomi. Un altro meccanismo che è particolarmente comune nel corbezzolo è l’embolismo xilematico causato da una perdita di conduttività dello xilema a causa dei bassi potenziali idrici nel periodo di stress ([29], [17]). Nel 2004, poiché non si sono registrati periodi di siccità prolungati a causa di un’anomala distribuzione delle precipitazioni, entrambi i trattamenti hanno mostrato sempre un elevato tasso traspirativo. Cherubini et al. ([5]) hanno verificato, utilizzando un metodo dendrocronologico, che il corbezzolo è caratterizzato da una elevata sensitività nei confronti delle precipitazioni probabilmente a causa del suo apparato radicale più superficiale se comparato a quello di altre specie tipiche della vegetazione mediterranea (per es., Quercus sp.p. e Fraxinus ornus - [1]). La crescita del corbezzolo è fortemente influenzata dalle precipitazioni primaverili dell’anno ([15]) e ciò sembra confermato dagli elevati tassi traspirativi, prossimi a quelli potenziali, misurati in condizioni idriche favorevoli (SWC > 10%). Martinez-Vilalta et al. ([17]) riportano una forte sensitività del corbezzolo nei confronti del deficit di umidità del suolo inteso come differenza tra la cumulata del flusso traspirativi e la cumulata delle precipitazioni. Nel caso riportato nel presente lavoro, abbiamo verificato una relazione tra umidità del suolo e rapporto tra ET ed ET0 al di sotto della soglia di umidità del 10%: complessivamente, i trattamenti applicati hanno determinato una riduzione nel tasso traspirativo nel DRY pari al 35% se confrontato con il WET, un valore molto simile a quello riportato da Wullschleger & Hanson ([32]) per un simile esperimento condotto in un bosco di querce.
L’andamento registrato della respirazione del suolo in relazione alla temperatura è in linea con gli andamenti misurati in altri ecosistemi mediterranei ([24], Curiel [7], [23]). Negli ambienti mediterranei le prime piogge dopo il secco estivo stimolano fortemente la respirazione determinando un flusso di CO2 verso l’atmosfera. L’effetto di stimolazione della respirazione del suolo dovuto alle precipitazioni può avvenire secondo tre meccanismi: una stimolazione della respirazione eterotrofa, dissoluzione di eventuali carbonati presenti nel suolo, aumento della respirazione autotrofa.
Nel nostro sito si è registrata una dinamica di respirazione del suolo simile per i due anni di studio: mentre il trattamento WET ha avuto una più alta respirazione del suolo nei mesi estivi durante l’esecuzione delle irrigazioni, in trattamento DRY sono stati registrati i tassi più alti in corrispondenza delle prime piogge autunnali. Ciò potrebbe essere spiegato dal fatto che la maggiore attività microbica del WET durante i mesi estivi, consentita dalle migliori condizioni idriche del suolo, avrebbe determinato una riduzione del substrato più facilmente decomponibile innescando un processo di feedback negativo nel periodo autunnale ([6], [12]).
Goulden et al. ([10]) hanno verificato che un periodo di secco eccezionale esercita una maggiore riduzione sulla respirazione che sulla fotosintesi suggerendo quindi un maggior effetto sulla respirazione eterotrofa che autotrofa. Questo fatto sembra essere confermato da Borken et al. ([3]) che hanno evidenziato come un periodo di siccità riduca la respirazione del suolo indipendentemente dalla temperatura del suolo e come la respirazione radicale sia meno influenzata dalla minore disponibilità idrica rispetto alla decomposizione della sostanza organica. Tutti questi elementi sono confermati a livello ecosistemico dalle misure effettuate con l’eddy covariance: nel WET si ha una riduzione della NEE, dovuta al fatto che, benché la GPP aumentasse, ciò era accompagnato da un maggiore aumento di Reco (respirazione autotrofa [soprassuolo e radici] respirazione eterotrofa), mentre nel DRY, la riduzione della respirazione ha favorito il mantenimento di una maggiore assimilazione netta da parte dell’ecosistema.
Conclusioni
Condizioni di stress idrico estivo determinano una diminuzione della capacità assimilativa da parte della canopy in ecosistemi mediterranei quali quello studiato nel presente lavoro, ma la contemporanea maggiore riduzione della respirazione ecosistemica determina un aumento del sequestro netto di carbonio nel trattamento DRY rispetto al WET. Inoltre, la minor disponibilità idrica nei mesi estivi determina una riduzione della respirazione del suolo anche se poi, in seguito alle piogge autunnali, la respirazione totale è molto simile nei due trattamenti. La disponibilità di substrato facilmente decomponibile ancora presente nel trattamento DRY alla fine della stagione secca sembra quindi controllare i flussi di respirazione non appena le condizioni di umidità del suolo tornano ad essere favorevoli per gli eterotrofi.
Ringraziamenti
Il lavoro è stato finanziato e condotto nell’ambito del Progetto MIND (EU, EVK2-CT-2002-00158).
References
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar