Distribution model of understory vegetation in beech forests from Central Apennines (Italy) in relation to edaphic parameters
Forest@ - Journal of Silviculture and Forest Ecology, Volume 4, Pages 439-449 (2007)
doi: https://doi.org/10.3832/efor0496-0040439
Published: Dec 20, 2007 - Copyright © 2007 SISEF
Research Articles
Abstract
The first results of a study of monitoring species and habitats in the framework of the Life 04NAT/IT/000190 “Tutela dei siti Natura 2000 gestiti dal Corpo Forestale dello Stato” are presented. This study was carried out by the Department of Plant Biology of the University of Rome (La Sapienza) and the National Forest Service. We focused the investigation on the relationships between herbaceous species and pedological parameters (humus and soil) in Central-Apennine beech forests. Data have been collected through 40 phytosociological relevés, 15 soil profiles and 40 humus profiles in 40 forest plots, between March 2005 and September 2006. In this paper we presented data elaborated on a subset of 15 plots where soil profiles were available. The species of undergrowth showed different ecological requirements to some edaphic and humus parameters (pH, sand, loam, clay, organic matter, nitrogen, carbon/nitrogen ratio, Ca++, K+;thickness and carbon content of the organic layers): significant correlations have been found using Pearson correlation test. The multiple regression analysis allowed to identify the factors more influencing the species distribution: thickness of the organic layers, carbon content (% C) and carbon/nitrogen ratio (C/N). Basing on the thickness of humus horizons (OL, OF, OH), and on carbon content, two species groups with different ecology have been recognized: (a) nemoral species typical of shady beech stands on oligomull/dysmull/thin amphimull (0 < 1cm.; 5.7 < %C < 9.9); (b) heliophilous species of more xeric stands on thick amphimull (1.25 < 11.5 cm.; 9.9 < %C < 13.7). The relationships among species and soil parameters and humus forms allow to recognize small differences within a homogeneous habitat and therefore they can provide management indications also at micro-scale level.
Keywords
Vegetation, Understory, Humus, Soil, Succession, Italy, Fagus sylvatica
Introduzione
Dalla selvicoltura tradizionale alla selvicoltura sistemica
Nel corso degli ultimi decenni, soprattutto a partire dalla fine del secondo conflitto mondiale, sono intervenuti cambiamenti a livello socio-economico che hanno condizionato il rapporto uomo - bosco, soprattutto nelle aree montane interne dell’Appennino dove la pressione antropica è drasticamente diminuita.
L’abbandono della montagna ha permesso alla vegetazione forestale il recupero di vaste superfici rispetto al precedente periodo in cui il bosco era stato fortemente utilizzato a fini produttivi per rispondere alle esigenze delle popolazioni montane. In questo periodo storico vengono acquisite al Demanio dello Stato molte proprietà, sulle quali il Corpo Forestale dello Stato istituisce, a partire dagli anni ’70, le prime Riserve Naturali, grazie alle quali è iniziato anche un importante processo di tutela e gestione diretta del territorio che ha costituito la base per la rete delle aree protette a livello nazionale e comunitario. È proprio in questa fase storica che anche grazie ad una sostanziale riduzione del prelievo legnoso, viene avviata una importante trasformazione gestionale: comincia ad affermarsi un approccio ecosistemico alla gestione delle foreste. In quest’ottica il bosco è considerato un ecosistema complesso in cui oltre alla componente arborea, assumono importanza anche quella erbacea, arbustiva, faunistica e microbiologica. La gestione passa quindi da una selvicoltura di tipo classico, finalizzata a massimizzare le produzioni legnose, ad una “naturalistica“ fino ad arrivare, negli ultimi anni, al recente concetto di selvicoltura “sistemica” ([4]).
In tale contesto si inserisce l’attività gestionale dell’Ufficio Territoriale per la Biodiversità di Castel di Sangro (UTB) che, nelle aree gestite direttamente dal Corpo Forestale dello Stato nell’Appennino abruzzese, ha avviato esperienze finalizzate al recupero ed alla salvaguardia degli ecosistemi forestali e a garantire livelli occupazionali e di reddito coerenti con le necessità del contesto socio economico.
L’esperienza maturata negli anni ’70 e ’80, è stata interamente applicata e sviluppata in una serie di progetti di tutela condotti dall’UTB, e in particolare il progetto “Conservazione dell’orso bruno nell’Appennino Centrale” (Life 99NAT/IT/006244) e il progetto “Tutela dei siti Natura 2000 gestiti dal Corpo Forestale dello Stato” (Life04NAT/IT/000190).
Su questi presupposti è stato avviato un progetto di ricerca condotto dal CFS e dal Dipartimento di Biologia Vegetale dell’Università La Sapienza, per elaborare un inventario della biodiversità vegetale nelle faggete dell’Abruzzo meridionale (area del progetto LIFE 2004 “Tutela dei Siti Natura 2000 gestiti dal Corpo Forestale dello Stato”, LIFE04NAT/IT/000190) e per individuare le principali minacce alla conservazione della biodiversità.
Fattori di disturbo
Le faggete dell’area di studio hanno una lunga storia di interventi selvicolturali, come in generale la maggior parte delle foreste a Fagus sylvatica dell’Appennino. Sovrapascolo, tagli indiscriminati ed incendi hanno causato nel tempo un deterioramento di questo habitat ([1], [15]). L’alterazione della canopy e l’assottigliamento dei suoli, costituiscono le conseguenze più evidenti di questo disturbo antropico. In ambiente mediterraneo non sono molti gli studi che si occupano degli aspetti strutturali e funzionali delle faggete in relazione al disturbo ([26]). La vegetazione primaria inalterata è attualmente quasi inesistente su scala planetaria ([22]) e in particolare i boschi italiani sono soggetti ad un antico e prolungato sfruttamento risalente alle prime popolazioni italiche ([5]), ma va sottolineato come anche in condizioni naturali esistano fattori di disturbo come la caduta degli alberi senescenti, gli incendi e le slavine che accendono una successione secondaria. Pertanto occorre analizzare gli effetti del disturbo a scale diverse e sulle diverse componenti dell’ecosistema (flora, fauna, vegetazione, paesaggio, suolo, ecc.) e individuare indicatori sensibili in grado di esprimerlo e misurarlo ([9]).
Variabilità delle faggete centro-appenniniche
La composizione dello strato erbaceo delle faggete appenniniche si presenta alquanto omogenea ([24]). Tuttavia, sono presenti lievi differenze che stimolano la ricerca di fattori ecologici che possano spiegare questa variabilità. In molti casi, infatti, i boschi del territorio in esame si presentano come un mosaico di parcelle con basse coperture e bassa diversità di specie erbacee accanto ad altre più ricche e spesso con estese popolazioni clonali di Cardamine spp., Galium odoratum, Viola reichenbachiana ecc. Tali discrepanze possono essere spiegate dal disturbo antropico che genera eterogeneità ambientale, a livello di specie, comunità, paesaggio ([19], [23]), creando dinamiche spesso di difficile interpretazione, in cui possono essere implicati fattori ambientali quali suolo, humus, clima, ma anche e non secondari, fattori di dispersione delle specie su cui non si hanno conoscenze sufficienti. Il dinamismo che ne consegue implica quindi l’esistenza di un complesso di fattori ecologici interagenti, la cui individuazione riveste una particolare importanza per la comprensione degli attuali processi ecologici, per poter quindi avanzare previsioni e programmare interventi gestionali mirati.
L’approccio fitosociologico ed ecologico in sinergia
L’approccio fitosociologico alle foreste enfatizza l’importanza dello strato erbaceo, perché permette di evidenziare differenze sottili nelle condizioni ecologiche dei boschi di caducifoglie, che la sola componente arborea non permette di rilevare; per questo, il lavoro si è concentrato sulle relazioni tra i parametri del suolo e dell’humus e la distribuzione delle specie erbacee. Conoscendo infatti i parametri edafici, è possibile prevedere quali specie erbacee siano potenzialmente presenti e, viceversa, osservando la presenza di determinate specie erbacee, si può stabilire quali siano le caratteristiche del suolo più probabili.
Pertanto, sono state approfondite le relazioni tra determinate specie e i fattori fisico-chimici che ne regolano la crescita ed è stata data priorità ad una componente dell’ecosistema che svolge un ruolo centrale, il suolo; in particolare, si è scelto di focalizzare la ricerca su due aspetti del “sistema suolo”: humus e parametri pedologici.
- Il primo consente inferenze sul ciclo del carbonio e dei nutrienti, essenziali per l’interpretazione della dinamica forestale, e quindi di inequivocabile interesse ai fini gestionali. Gli humus sono infatti il risultato integrato di una serie di processi fondamentali nell’ecosistema, in particolare il ciclo dei nutrienti e del carbonio. Il profilo dell’humus è collegato con la velocità di decomposizione della lettiera e di incorporazione della sostanza organica nel suolo. Il tipo di humus è anche correlato con la fertilità del suolo, e quindi indirettamente con la potenzialità di rigenerazione della foresta ([29]); rappresenta uno dei pool terrestri di carbonio più importanti, e viene quindi attivamente studiato nell’ambito delle indagini sui cambiamenti climatici globali. Esso è infine è influenzato dalle condizioni climatiche e dal tipo di pedofauna e pedoflora. In ambiente appenninico lo studio degli humus rappresenta una novità; non esistono infatti lavori sulle forme di humus e sul loro ruolo ecologico. La letteratura specifica è di origine francese ([30]a, [31]b) e gli studi finora effettuati si riferiscono principalmente all’Italia settentrionale ([3]), mentre scarsi sono quelli relativi all’Italia peninsulare ([32]).
- I parametri pedologici analizzati sono quelli definiti “di base” per identificare le caratteristiche fondamentali del suolo e la sua dotazione in elementi nutritivi ([17], [12], [13]). Anche il suolo è di fondamentale importanza per capire la dinamica evolutiva di un bosco; infatti la silvicoltura stessa dipende dal suolo per l’apporto di acqua e nutrienti. Il suolo svolge inoltre un ruolo centrale per la protezione dell’acqua e lo scambio di gas con l’atmosfera, grazie a funzioni di magazzinaggio, filtraggio, tampone e trasformazione: immagazzina e in parte trasforma minerali, materia organica, acqua, energia e diverse sostanze chimiche. È stata posta particolare attenzione al rapporto carbonio/azoto [C/N], indicatore dell’evoluzione della sostanza organica nel suolo; tale rapporto è infatti indice del turnover della sostanza organica e in particolare della velocità con la quale essa è riciclata dal sistema suolo-vegetazione ([6], [7], [16]).
Area di Studio
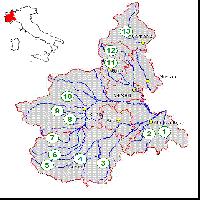
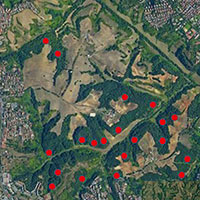
L’area presa in esame corrisponde a 6 siti di importanza comunitaria ubicati nell’Alto corso del fiume Sangro, tra Abruzzo e Molise (Fig. 1, Tab. 1).
Fig. 1 - Localizzazione dei siti di studio. Con i triangoli sono indicate le parcelle in cui sono stati effettuati rilievi fitosociologici, profili di humus e profili di suolo, mentre i cerchi indicano le parcelle in cui sono stati effettuati rilievi fitosociologici e profili di humus.
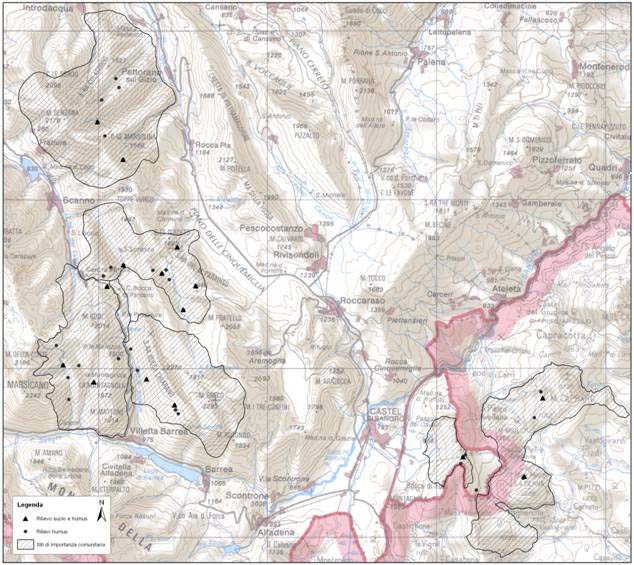
Tab. 1 - Sono riportati i nomi dei SIC e le codifiche delle parcelle campionate con le relative coordinate.
| SIC | Superficie totale (ha) | Superficie faggeta (%) | Codice parcelle | Coord. Est | Coord. Nord |
|---|---|---|---|---|---|
| UTM ED50 | |||||
| M.te Genzana | 5818 | 40.9 | 1.1 | 411443 | 4642193 |
| 1.4 | 409862 | 4644629 | |||
| M.te Pagano e Feudozzo | 733 | 17.1 | 2.1 | 431521 | 4624777 |
| Chiarano - Sparvera | 2754 | 48.2 | 3.1 | 414757 | 4636917 |
| 3.3 | 413740 | 4635695 | |||
| 3.6 | 411548 | 4636107 | |||
| 3.9 | 415713 | 4634910 | |||
| Feudo Intramonti - M.te Godi e Ferroio di Scanno | 3819 | 54.1 | 5.2 | 410501 | 4634826 |
| 5.3 | 409243 | 4633806 | |||
| 5.6 | 407856 | 4630196 | |||
| 5.9 | 409708 | 4629479 | |||
| Serra di Rocca Chiarano - M.te Greco | 3468 | 18.3 | 6.1 | 412636 | 4629419 |
| 6.4 | 414441 | 4627867 | |||
| M.te di Mezzo - M.te Miglio - Pennataro - M.te Capraro - M.te Cavallerizzo | 3954 | 25.4 | 7.3 | 435148 | 4623609 |
| 7.5 | 436246 | 4628253 | |||
L’area presenta caratteristiche tipiche dell’Appennino centrale su substrato calcareo e suoli poco profondi. I boschi analizzati rappresentano l’espressione tipica della faggeta interna appenninica, classificata come Polysticho-Fagetum Feoli & Lagonegro ([11]): si tratta della vegetazione finale della fascia subatlantica tra 1200-1900 m s.l.m., caratteristica delle vallate interne dell’Appennino centrale con clima subcontinentale, costituito da uno strato arboreo composto quasi esclusivamente da Faggio e sottobosco con specie erbacee tipiche della faggeta centroeuropea.
Il Polysticho-Fagetum è diffuso all’interno dell’Appennino centrale, generalmente nelle aree a maggiore continentalità, cioè sottratte all’influenza delle correnti umide provenienti dal Tirreno.
Di questa associazione sono state descritte diverse tendenze evolutive, corrispondenti alle differenti caratteristiche di suolo, pendenza e copertura vegetale ([24]).
Le faggete rilevate nell’area di studio sono comprese tra 1114 e 1992 m di quota.
I rilievi effettuati
I dati presentati sono relativi a 15 parcelle in cui sono stati effettuati profili di suolo e per le quali sono disponibili i risultati delle analisi floristiche, vegetazionali e pedologiche.
La scelta delle stazioni di rilievo è stata effettuata tenendo conto congiuntamente degli aspetti morfologici, pedologici, vegetazionali e selvicolturali. Ciascun sito è stato considerato campionabile solo se omogeneo rispetto ai quattro aspetti sopra indicati.
La caratterizzazione della stazione è stata eseguita mediante:
- apertura e campionamento di ogni orizzonte per le analisi fisico-chimiche di routine per i profili sia di humus che del suolo;
- descrizione macroscopica del profilo di suolo ([28]) e dell’humus ([16]) in corrispondenza del profilo pedologico e in altri punti dell’area campione;
- descrizione dello strato arboreo e valutazione della copertura arborea in corrispondenza del profilo;
- rilievo fitosociologico ([20], [22]).
Per ogni orizzonte sia del profilo di humus che di quello pedologico è stata raccolta una quantità sufficiente di campioni da analizzare. I campioni sono stati conservati in sacchetti di plastica, avendo cura di evitare contaminazioni di qualsiasi tipo e successivamente portati in laboratorio.
I campioni di suolo, prima di essere analizzati sono stati sottoposti alla seguente procedura: essiccamento all’aria (3-5 giorni); eliminazione dello scheletro >2 mm e delle foglie; frantumazione degli aggregati più duri con l’ausilio di un mortaio; setacciamento con vaglio a 2 mm. Sui campioni così preparati sono state effettuate le analisi fisico-chimiche standard ([18]): Granulometria e Tessitura; pH in H2 O; Carbonati totali; Carbonio organico e Sostanza organica; Acidità di scambio; Azoto totale; Fosforo assimilabile; AWC (Acqua disponibile per le piante) (vedi Tab. 4 in seguito). Sono riportati nella tabella citata i valori delle analisi effettuate relativi a tutti gli orizzonti dei profili. Particolare attenzione è stata posta all’orizzonte A, in quanto è quello che contrae rapporti più stretti con gli strati dell’humus e con le specie erbacee.
Tab. 4 - Risultati delle analisi pedologiche. Sono riportati per ogni parametro le diverse unità di misura e i relativi valori. (Sost.org. = sostanza organica; C = carbonio organico; N tot. = azoto totale; P ass. = fosforo assimilabile; Ac scambio = acidità di scambio; CSC = capacità di scambio cationico; Sat. in basi = saturazione in basi; % s. gr.= sabbia grossa; % s.m. = sabbia media; % s.fi. = sabbia fine; % arg. = argilla; AWC = Acqua disponibile per le piante).
| Parcelle | Orizzonti | pH in H2 O | CaCO3 % | sost.org. % | C org. % | N tot. % | C/N | P ass. Ppm | Basi scambiabili (meq/100gr) | Ac. scambio meq/ 100gr |
CSC meq/ 100gr |
Sat.in basi % | Granulometria | AWC (mm) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mg | Ca | Na | K | %s.gr | %s.m. | %s.fi. | %limo | %arg. | |||||||||||||
| 1.1 | A1 | 5.38 | 0 | 13.43 | 7.79 | 0.74 | 10.53 | 23.20 | 2.97 | 16.91 | 0.49 | 0.92 | 29.19 | 50.48 | 42.2 | 9.30 | 2.91 | 3.15 | 52.29 | 32.34 | 247.17 |
| A2 | 6.55 | 0 | 8.72 | 5.06 | 0.42 | 12.05 | 2.61 | 1.37 | 22.31 | 2.79 | 0.28 | 17.89 | 44.65 | 59.9 | 9.40 | 3.00 | 3.38 | 56.83 | 27.39 | ||
| 1.4 | A1 | 7.14 | 0 | 16.19 | 9.39 | 0.72 | 13.04 | 16.53 | 2.97 | 32.85 | 0.36 | 0.92 | 10.43 | 47.53 | 78.1 | 20.76 | 5.03 | 4.64 | 51.67 | 17.90 | 104.72 |
| A2 | 7.53 | 0 | 10.24 | 5.94 | 0.41 | 14.49 | 5.04 | 1.60 | 41.72 | 0.36 | 0.50 | 7.58 | 51.76 | 85.4 | 21.08 | 5.55 | 5.48 | 37.81 | 21.08 | ||
| 2.1 | A1 | 7.24 | 3.84 | 21.86 | 12.68 | 1.33 | 9.53 | 47.50 | 2.89 | 40.19 | 0.36 | 1.35 | 8.24 | 53.04 | 84.5 | 5.94 | 1.56 | 3.00 | 57.50 | 31.99 | 155.54 |
| A2 | 7.55 | 4.8 | 17.67 | 10.25 | 1.15 | 8.91 | 58.99 | 2.06 | 79.97 | 1.21 | 1.14 | 9.29 | 93.67 | 90.1 | 8.93 | 2.94 | 5.42 | 69.34 | 13.36 | ||
| 3.1 | A1 | 7.2 | 0 | 22.77 | 13.21 | 1.32 | 10.01 | 32.36 | 2.97 | 45.32 | 5.79 | 0.71 | 10.31 | 65.10 | 84.2 | 0.53 | 1.21 | 1.58 | 87.61 | 9.06 | 254.65 |
| A2 | 7.62 | 2.88 | 19.19 | 11.13 | 1.05 | 10.60 | 17.75 | 5.48 | 69.30 | 3.27 | 0.71 | 5.83 | 84.60 | 93.1 | 7.10 | 1.54 | 2.65 | 80.04 | 8.66 | ||
| 3.3 | A | 5.66 | 0 | 10.29 | 5.97 | 0.46 | 12.98 | 7.40 | 2.51 | 17.46 | 0.61 | 0.64 | 22.67 | 43.89 | 48.4 | 1.88 | 1.06 | 2.19 | 55.86 | 39.01 | 249.82 |
| B | 5.46 | 0 | 4.27 | 2.48 | 0.20 | 12.40 | 4.00 | 1.83 | 14.83 | 0.49 | 0.36 | 22.32 | 39.82 | 43.9 | 1.28 | 0.90 | 1.66 | 58.16 | 38.00 | ||
| 3.6 | A1 | 7.06 | 0 | 23.57 | 13.67 | 1.14 | 11.99 | 20.88 | 4.80 | 39.08 | 0.49 | 1.35 | 13.13 | 58.85 | 77.7 | 4.06 | 3.63 | 2.97 | 69.71 | 19.62 | 144.53 |
| A2 | 7.74 | 3.36 | 19.10 | 11.08 | 0.72 | 15.39 | 12.18 | 2.06 | 63.89 | 0.49 | 0.71 | 6.46 | 73.60 | 91.2 | 16.47 | 4.71 | 5.83 | 44.55 | 28.43 | ||
| 3.9 | A1 | 6.42 | 0 | 15.52 | 9.00 | 0.75 | 12.00 | 34.00 | 3.43 | 20.79 | 0.36 | 1.14 | 17.42 | 43.14 | 59.6 | 6.41 | 3.67 | 6.11 | 56.59 | 27.22 | 121.24 |
| A2 | 6.85 | 0 | 6.72 | 3.90 | 0.32 | 12.19 | 3.31 | 2.06 | 28.14 | 0.61 | 0.78 | 12.37 | 43.95 | 71.9 | 9.93 | 5.54 | 9.64 | 57.74 | 17.15 | ||
| 5.2 | A | 6.59 | 0 | 13.34 | 7.74 | 0.54 | 14.33 | 34.45 | 3.88 | 22.59 | 0.61 | 1.92 | 14.72 | 43.72 | 66.3 | 1.08 | 1.28 | 2.54 | 59.32 | 35.77 | 146.69 |
| B | 6.96 | 0 | 5.90 | 3.42 | 0.24 | 14.25 | 3.83 | 1.83 | 30.08 | 0.61 | 0.92 | 9.79 | 43.22 | 77.3 | 0.46 | 1.42 | 3.05 | 57.88 | 37.19 | ||
| 5.3 | A1 | 6.34 | 0 | 17.72 | 10.28 | 0.72 | 14.28 | 27.60 | 4.80 | 46.85 | 3.76 | 0.64 | 19.76 | 75.80 | 73.9 | 0.38 | 1.10 | 3.70 | 69.29 | 25.52 | 315.39 |
| A2 | 7.59 | 0 | 6.84 | 3.97 | 0.32 | 12.41 | 0.00 | 1.37 | 28.14 | 3.88 | 0.28 | 7.66 | 41.33 | 81.5 | 0.22 | 0.99 | 7.73 | 74.96 | 16.10 | ||
| B | 7.63 | 0 | 5.40 | 3.13 | 0.26 | 12.04 | 0.35 | 1.60 | 40.05 | 0.97 | 0.28 | 7.97 | 50.88 | 84.3 | 0.30 | 0.88 | 6.57 | 76.40 | 15.86 | ||
| 5.6 | A1 | 6.35 | 0 | 14.88 | 8.63 | 0.54 | 15.98 | 56.20 | 3.20 | 23.42 | 1.46 | 1.42 | 14.06 | 43.56 | 67.7 | 1.28 | 0.45 | 0.64 | 65.91 | 31.72 | 169.13 |
| A2 | 6.39 | 0 | 9.55 | 5.54 | 0.38 | 14.58 | 94.00 | 2.51 | 27.58 | 1.09 | 0.92 | 13.55 | 45.66 | 70.3 | 0.46 | 0.62 | 1.28 | 65.58 | 32.06 | ||
| B | 6.73 | 0 | 2.38 | 1.38 | 0.14 | 9.86 | 69.25 | 5.25 | 29.52 | 1.58 | 0.99 | 10.19 | 47.54 | 78.6 | 0.33 | 0.53 | 0.81 | 66.33 | 32.00 | ||
| 5.9 | A1 | 6.52 | 0 | 9.77 | 5.67 | 0.52 | 10.90 | 119.02 | 2.51 | 31.88 | 1.09 | 0.71 | 26.72 | 62.91 | 57.5 | 0.55 | 0.79 | 4.14 | 77.90 | 16.62 | 215.49 |
| A2 | 6.71 | 0 | 7.48 | 4.34 | 0.42 | 10.33 | 95.87 | 1.60 | 32.71 | 0.97 | 0.36 | 22.99 | 58.62 | 60.8 | 0.48 | 0.85 | 5.24 | 77.22 | 16.21 | ||
| B | 7.37 | 0 | 4.46 | 2.59 | 0.23 | 11.26 | 76.39 | 1.37 | 36.87 | 0.97 | 0.21 | 14.21 | 53.63 | 73.5 | 0.46 | 1.05 | 6.76 | 76.03 | 15.70 | ||
| 6.1 | A | 6.67 | 0 | 14.08 | 8.17 | 0.54 | 15.13 | 3.83 | 2.97 | 34.51 | 1.09 | 0.99 | 5.30 | 44.87 | 88.2 | 0.84 | 0.55 | 1.74 | 65.44 | 31.42 | 285.16 |
| B1 | 7.33 | 0 | 8.57 | 4.97 | 0.39 | 12.74 | 1.74 | 2.28 | 35.34 | 1.09 | 0.43 | 13.73 | 52.87 | 74.0 | 0.64 | 0.70 | 1.95 | 69.36 | 27.34 | ||
| B2 | 7.79 | 0 | 6.27 | 3.64 | 0.29 | 12.55 | 0.00 | 1.14 | 47.12 | 1.58 | 0.78 | 10.41 | 61.03 | 82.9 | 0.80 | 0.65 | 2.20 | 68.61 | 27.74 | ||
| 6.4 | A1 | 6.5 | 0 | 16.34 | 9.48 | 0.77 | 12.31 | 12.60 | 3.88 | 27.30 | 0.97 | 1.07 | 18.72 | 51.94 | 64.0 | 4.85 | 2.89 | 5.07 | 66.87 | 20.32 | 214.78 |
| A2 | 6.41 | 0 | 11.52 | 6.68 | 0.79 | 8.46 | 6.60 | 1.14 | 26.61 | 1.09 | 0.43 | 22.36 | 51.63 | 56.7 | 3.74 | 1.20 | 3.42 | 80.33 | 5.65 | ||
| 7.3 | A | 7.44 | 0 | 17.12 | 9.93 | 0.72 | 13.79 | 18.62 | 4.11 | 41.72 | 1.46 | 1.92 | 6.32 | 55.52 | 88.6 | 1.21 | 1.58 | 4.83 | 50.15 | 42.23 | 266.96 |
| B1 | 7.42 | 0 | 4.93 | 2.86 | 0.29 | 9.86 | 5.57 | 3.20 | 28.00 | 1.09 | 1.14 | 4.74 | 38.16 | 87.6 | 0.41 | 1.60 | 9.58 | 51.60 | 36.81 | ||
| B2 | 7.81 | 1.92 | 1.34 | 0.78 | 0.09 | 8.67 | 2.26 | 2.51 | 23.01 | 0.85 | 0.50 | 2.69 | 29.56 | 90.9 | 2.71 | 8.26 | 19.15 | 45.88 | 24.00 | ||
| C | 8.35 | 10.55 | 0.88 | 0.51 | 0.06 | 8.50 | 3.83 | 4.57 | 36.59 | 0.85 | 0.36 | 0.00 | 42.36 | 100.0 | 1.07 | 1.28 | 5.82 | 75.82 | 16.00 | ||
| 7.5 | A | 7.55 | 0 | 17.01 | 9.87 | 0.87 | 11.34 | 19.66 | 3.20 | 39.36 | 5.31 | 1.84 | 6.73 | 56.44 | 88.1 | 4.26 | 2.74 | 3.93 | 46.89 | 42.17 | 95.77 |
| B | 7.14 | 0 | 8.31 | 4.82 | 0.40 | 12.05 | 7.83 | 1.60 | 28.41 | 7.00 | 1.41 | 5.03 | 43.46 | 88.4 | 6.20 | 3.05 | 4.80 | 45.59 | 40.35 | ||
I parametri degli humus considerati nell’analisi sono stati: spessore totale dell’episolum umifero (cioè l’insieme degli orizzonti - OL, OF e OH - gli orizzonti organici di un suolo, la cui organizzazione dipende essenzialmente dall’attività biologica), spessori dei singoli orizzonti OL (costituto da residui vegetali interi o poco frammentati, suddivisibile a sua volta in sotto orizzonti OLn - lettiera di recente apporto - e OLv - lettiera invecchiata -), OF (costituito da residui vegetali sminuzzati, trasformati, ma ancora riconoscibili ad occhio nudo, mescolati ad una quantità variabile di sostanza organica fine < 70%), OH (costituito da < 30% di residui vegetali ancora riconoscibili ad occhio nudo e da più del 70% del volume di sostanza organica fine) e % di carbonio organico in essi misurato utilizzando il metodo di Green et al. ([14]).
Elaborazione dei dati
I dati floristici, vegetazionali e pedologici sono stati organizzati in una matrice floristica/ecologica “coperture delle specie/humus/parametri edafici”. La matrice floristica è ricavata dai rilievi fitosociologici e, secondo i criteri standard, è strutturata in righe e colonne, rispettivamente specie e rilievi. I numeri all’incrocio delle righe con le colonne esprimono i valori di copertura attribuiti secondo la scala da a 5 di Braun-Blanquet modificata da Pignatti ([20]).
Per una prima indagine esplorativa tesa a evidenziare l’esistenza di relazioni significative tra coperture delle specie e parametri del suolo è stato applicato il test di correlazione di Pearson. Per quest’analisi le specie sono state ripartite in sciafile, con valore di luce < 5, ed eliofile, con valore di luce > 5, utilizzando il modello di bioindicazione secondo Ellenberg ([8]), in base al quale ad ogni specie è attribuito un valore di indicazione per la luce (L) secondo una scala compresa tra 1 e 10. I valori di bioindicazione sono disponibili in banche dati ([25], [10]). I parametri risultati più correlati sono: spessore dell’episolum umifero, % di carbonio organico misurato in humus e rapporto carbonio/azoto (C/N); è stata applicata un’analisi di regressione multipla, utilizzando la procedura stepwise, per individuare quale delle tre variabili fosse la più correlata con la variazione della copertura delle singole specie.
La nomenclatura delle specie segue Flora d’Italia ([21]).
Risultati
Specie e parametri del suolo
I risultati delle analisi pedologiche associate ai rilievi degli orizzonti organici e di vegetazione hanno evidenziato i parametri più influenti sulla distribuzione delle specie nel sottobosco delle faggete esaminate. Su un totale di 61 specie censite, 48 (78.7%) mostrano correlazioni significative tra la loro copertura ed uno o più parametri del suolo misurati (pH, sostanza organica, rapporto carbonio/azoto, calcio, azoto, potassio, argilla, sabbie, carbonio negli strati organici), e la metà di queste correlazioni mostra un valore elevato del coefficiente di correlazione, con r > 0.6.
Su 39 specie sciafile, 8 sono risultate significativamente e positivamente correlate con lo spessore dell’episolum, 5 sono negativamente correlate con la % di carbonio organico negli orizzonti organici; nessuna delle specie eliofile è risultata correlata positivamente con lo spessore dell’episolum, 2 di esse (su un totale di 11) mostra una correlazione negativa con lo spessore dell’episolum e altre 2 risultano correlate con il rapporto C/N nell’orizzonte A.
La stepwise regression ha permesso di selezionare il set variabili correlato con ciascuna specie (Tab. 2). Appare una risposta chiaramente diversificata: specie decisamente nemorali e sciafile, come Actaea spicata, Cephalanthera damasonium, Epipactis helleborine, Viola riviniana, etc, risultano significativamente e positivamente correlate con lo spessore dell’episolum; Lamium garganicum, specie eliofila ed euriecia, risulta negativamente correlata con la % di carbonio organico misurato negli orizzonti organici (O); infine specie più legate ai margini, come Digitalis micrantha e Vicia cracca, risultano legate al rapporto carbonio/azoto, indicando condizioni localizzate di accumulo di carbonio organico (indecomposto), dal quale probabilmente traggono vantaggio.
Tab. 2 - Risultati della regressione multipla ottenuti con la procedura stepwise. Sono riportate le significatività e i valori di bioindicazione per la luce (L). (*): p < 0.05; (**): p < 0.01.
| L | Specie | Spessore episolum | C% in humus | C/N | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Beta | SE | R | R2 | Adj. R2 | Beta | SE | R | R2 | Adj. R2 | Beta | SE | R | R2 | Adj. R2 | |||
| 2 | Actaea spicata | 0.71** | 0.06 | 0.71 | 0.51 | 0.47 | - | - | - | - | - | - | - | - | - | - | |
| 4 | Epipactis helleborine | 0.56* | 0.13 | 0.57 | 0.32 | 0.27 | - | - | - | - | - | - | - | - | - | - | |
| 3 | Cephalanthera damasonium | 0.68** | 0.12 | 0.67 | 0.45 | 0.30 | - | - | - | - | - | - | - | - | - | - | |
| 3 | Sanicula europaea | 0.45* | 0.14 | 0.47 | 0.22 | 0.007 | - | - | - | - | - | - | - | - | - | - | |
| 4 | Euonymus europaeus | 0.54* | 0.11 | 0.54 | 0.29 | 0.24 | - | - | - | - | - | - | - | - | - | - | |
| 4 | Euphorbia amygdaloides | 0.54* | 0.11 | 0.54 | 0.29 | 0.24 | - | - | - | - | - | - | - | - | - | - | |
| 3 | Viola riviniana | 0.71** | 0.06 | 0.07 | 0.51 | 0.47 | - | - | - | - | - | - | - | - | - | - | |
| 8 | Lamium garganicum | - | - | - | - | - | - 0.52* | 0.10 | 0.50 | 0.30 | 0.20 | - | - | - | - | - | |
| 8 | Vicia cracca | - 0.53* | 0.17 | 0.53 | 0.3 | 0.22 | - | - | - | - | - | 0.44* | 0.50 | 0.58 | 0.34 | 0.16 | |
| 6 | Digitalis micrantha | - | - | - | - | - | - | - | - | - | - | 0.59** | 0.30 | 0.60 | 0.40 | 0.30 | |
Specie e forme di Humus
La tabella specie/rilievi può essere ordinata secondo diversi assi relativi ai parametri edafici, ciascuno evidenziando un gruppo di specie con ecologia significativa e caratteristica. Particolarmente interessante si presenta un ordinamento secondo un asse corrispondente agli spessori degli orizzonti OH e OF dell’episolum umifero, e quindi coincidente con la classificazione delle diverse forme di humus rilevate (Tab. 3).
Tab. 3 - Tabella ordinata per gruppi di specie e forme di humus. Sono evidenziati nei riquadri i gruppi di specie discriminati dagli humus. (A = strato arboreo dominante; B = strato arboreo dominato; C = strato arbustivo; pl.= plantule).
| SIC | 1.1 | 3.3 | 6.1 | 5.6 | 3.9 | 5.2 | 3.1 | 5.9 | 1.4 | 7.5 | 7.3 | 6.4 | 3.6 | 5.3 | 2.1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Carbonio in orizzonte A (%) | 7.8 | 6 | 8.2 | 8.6 | 9 | 7.7 | 13.2 | 5.7 | 9.4 | 9.9 | 9.9 | 9.5 | 13.7 | 10.3 | 12.7 |
| OLn+Olv (cm) | 1.25 | 3.5 | 2.3 | 2.5 | 3.5 | 3.5 | 4.5 | 2.5 | 1.5 | 5 | 3.5 | 1.25 | 2.74 | 2.5 | 3.5 |
| OF (cm) | 0.75 | 1.5 | 1.5 | 2.3 | 3.5 | 3.5 | 9.5 | 0.5 | 0.75 | 3.5 | 2.5 | 0.75 | 0.5 | 1.5 | 3.5 |
| OH (cm) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.5 | 0.5 | 0.75 | 1.25 | 1.5 | 4.5 | 5 | 12 |
| Forme di Humus | Oligomull | Dysmull | Amphimull sottile | Amphimull spesso | |||||||||||
| Ubiquiste | |||||||||||||||
| Fagus sylvatica A | 4 | 3 | 3 | 4 | 2 | 3 | 4 | 4 | 4 | - | - | 2 | 4 | 3 | 4 |
| Fagus sylvatica B | 2 | 1 | 3 | 1 | - | 2 | 1 | 1 | 1 | 2 | - | 1 | 3 | 1 | 2 |
| Fagus sylvatica C | 2 | - | 4 | 2 | 1 | 3 | - | 1 | 1 | 1 | - | - | 2 | 3 | - |
| Fagus sylvatica pl. | + | + | - | + | + | + | - | + | + | + | - | - | + | - | + |
| Oligomull/dysmull/amphimull sottile | |||||||||||||||
| Mycelis muralis | + | + | - | + | + | + | + | + | - | - | - | - | + | - | - |
| Cardamine bulbifera | + | + | + | 1 | 1 | - | - | 1 | + | - | - | - | - | - | + |
| Cardamine enneaphyllos | 1 | - | + | + | + | - | + | + | 1 | - | - | - | - | - | + |
| Corydalis cava | + | + | + | + | + | - | + | + | - | - | - | - | - | - | - |
| Chaerophyllum aureum | - | + | - | + | + | - | + | + | - | - | - | - | - | - | - |
| Adenostyles australis | - | + | + | - | + | - | - | - | + | - | - | - | - | - | - |
| Galium aparine | - | + | - | - | 1 | - | - | + | - | - | - | - | - | - | - |
| Digitalis micrantha | - | - | + | - | - | + | - | - | + | - | - | - | - | - | - |
| Campanula trachelium | - | - | - | + | - | - | + | + | - | - | - | - | - | - | - |
| Lathyrus venetus | - | - | 1 | - | - | - | + | - | - | - | - | - | - | - | - |
| Dysmull | |||||||||||||||
| Myosotis sylvatica | - | - | - | + | - | - | + | - | - | - | - | - | - | - | - |
| Senecio fuchsii | - | - | + | + | + | - | - | - | - | - | - | - | - | - | - |
| Cephalanthera longifolia | - | - | - | - | + | + | - | - | - | - | - | - | - | - | - |
| Oligomull/dysmull/amphimull sottile (amphimull spesso) | |||||||||||||||
| Vicia cracca | - | + | + | + | - | + | - | + | 1 | + | - | + | - | - | - |
| Milium effusum | 1 | - | + | - | - | + | - | + | + | - | - | 2 | 1 | - | - |
| Viola reichembachiana | + | + | - | - | - | - | - | + | - | - | - | + | - | - | - |
| Dysmull/amphimull | |||||||||||||||
| Cephalanthera damasonium | - | - | - | - | - | + | - | - | - | - | - | - | + | - | + |
| Lathyrus vernus | - | + | - | - | - | - | - | - | + | - | - | - | 1 | - | + |
| Rubus idaeus | - | - | - | - | - | + | - | + | - | 2 | - | + | - | - | 1 |
| Acer campestre pl. | - | - | - | - | - | + | + | - | - | + | - | - | + | - | - |
| Amphimull spesso | |||||||||||||||
| Arabis alpina | - | - | - | - | - | - | - | - | + | - | - | + | - | - | - |
| Arum italicum | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + |
| Q. cerris | - | - | - | - | - | - | - | - | - | 2 | - | - | - | - | 2 |
| Acer campestre | - | - | - | - | - | - | - | - | - | + | - | - | + | - | - |
| Acer obtusatum | - | - | - | - | - | - | - | - | - | - | - | - | + | - | 1 |
| Euonymus europaeus pl. | - | - | - | - | - | - | - | - | - | + | - | - | - | - | + |
| Euphorbia amygdaloides | - | - | - | - | - | - | - | - | - | + | - | - | - | - | + |
| Amphimull sottile | |||||||||||||||
| Galium album | - | - | - | - | - | + | - | 1 | - | - | - | - | - | - | - |
| Geranium versicolor | - | - | - | - | - | - | - | + | + | - | - | - | - | - | - |
| Hedera helix | - | - | - | - | - | - | - | - | - | + | - | - | - | - | 3 |
| Hepatica nobilis | - | - | - | - | - | - | + | - | + | - | - | - | - | - | - |
| Hieracium sylvaticum | - | - | - | - | - | - | - | + | + | - | - | - | - | - | - |
| Lamium garganicum | - | - | - | - | - | - | - | + | + | - | - | - | - | - | - |
Questo asse di spessore degli strati ectorganici evidenzia prima di tutto un corposo gruppo di specie legato alle faggete (Mycelis muralis, Cardamine bulbifera, Corydalis cava, ecc.) che si sviluppano su oligomull/dysmull/amphimull sottile, nettamente distinto da un gruppo più sparso di specie trasgressive delle cerrete (Quercus cerris, Acer obtusatum, ecc.), legati ad amphimull spesso. È possibile, ma l’ipotesi richiede accurate verifiche, che il rallentato tasso di decomposizione e quindi il maggiore spessore degli strati ectorganici dell’amphimull spesso sia in questo contesto legato allo stress idrico, con conseguente ingresso di specie delle cerrete più xerotermofile. Si noti che nel primo gruppo, gli spessori dell’orizzonte OH sono marcatamente inferiori a quelli del secondo gruppo, indicando una più veloce decomposizione dei residui vegetali; corrispondentemente, nei siti di questo gruppo si riscontrano anche valori più bassi della percentuale di carbonio organico misurato nell’orizzonte A del suolo (Tab. 3). Accanto a questa distinzione principale si osservano gruppi più piccoli di specie legati a singole forme di humus. In particolare un gruppo di specie (Myosotis sylvatica, Senecio fuchsii, Cephalanthera longifolia) è legato esclusivamente a dysmull; un altro di specie genericamente termofile (Geranium versicolor, Hedera helix, Hepatica nobilis, Lamium garganicum), è legato esclusivamente ad amphimull sottile; mentre un ultimo gruppo (Milium effusum, Viola reichenbachiana) si sviluppa su una maggiore diversità di humus, dagli oligomull agli amphimull sottili (Tab. 3).
Discussione
La risposta differenziata delle specie ai parametri del suolo
Le specie delle faggete studiate mostrano risposte multifattoriali ai parametri del suolo e delle forme di humus, evidenziando la multidimensionalità della nicchia: tra i molteplici fattori che la descrivono, si è visto che la distribuzione delle specie del sottobosco appare legata a parametri statici collegati al ciclo del carbonio ([2]). Molte specie sono legate allo spessore degli strati ectorganici; si può ipotizzare - ma ulteriori studi, anche di carattere fitosociologico, sono necessari per confermare queste ipotesi - che questa variabile sia legata al pool di carbonio nell’ecosistema, e anche al bilancio idrico dello strato erbaceo. Altre specie sono legate al rapporto carbonio/azoto (C/N), che è un indice del turnover della sostanza organica e in particolare della velocità con cui essa è riciclata dal sistema suolo-vegetazione[1] ([33]). I due gruppi sono anche distinti da valori medi, risultati significativamente diversi con il test T (p < 0.05), di carbonio organico misurato nell’orizzonte A: valore più basso (8.5%) nel gruppo delle specie più nemorali e valore più elevato (11.2%) in quello con specie trasgressive di cerrete più termoxerofile, con ciclo più lento di decomposizione. Infine, si osserva che la somma delle coperture delle specie del primo gruppo è il 90.5 % della copertura totale, mentre la somma delle coperture del secondo gruppo è il 70% del totale.
Il modello osservato di distribuzione delle specie del sottobosco esprime una diversità funzionale basata su una molteplicità di fattori, tra i quali è stato possibile selezionare quelli più influenti, che possono fungere da indicatori dello stato dell’habitat, discriminando differenze sottili nelle parcelle forestali esaminate:
- Specie prettamente nemorali come Actaea spicata, Epipactis helleborine, Viola riviniana, Cephalanthera damasonium, Sanicula europea, correlate con lo spessore dell’episolum umifero potrebbero essere buone indicatrici di boschi più maturi.
- Specie ruderali come Vicia cracca o più legate ai margini della foresta, come Digitalis micrantha, inversamente correlate con la quantità di carbonio organico nell’episolum umifero o legate a valori più alti del rapporto C/N, indicano una condizione di disturbo più accentuato probabilmente dovuto al pregresso carico di pascolo e di tagli ([26]).
Va osservato che tra le specie più nemorali ne compaiono alcune generalmente considerate sinantropiche come Chaerophyllum aureum, Elymus caninus, e che invece, oltre a essere probabilmente presenti nella composizione primordiale della faggeta, rappresentano una importante risorsa pabulare per l’orso, nonché specie come Mycelis muralis, Digitalis micrantha, legate a maggiori quantità di carbonio organico e a un rapporto C/N più alto, indicatrici di aspetti diradati; alcune di queste specie sono caratteristiche dell’associazione Digitalidi-Fagetum [11]; ci si attende una loro diminuzione con il procedere della rinaturalizzazione del bosco.
Conclusioni
L’apparente estrema omogeneità floristico-strutturale delle faggete dei SIC dell’Alto Sangro nasconde in realtà una notevole differenziazione ecologica che può essere messa in evidenza attraverso una serie di parametri degli strati più superficiali del suolo. Attraverso questa analisi, appaiono gruppi di specie indicatrici di diverse microtipologie di faggete. La disomogeneità osservata nella distribuzione delle specie del sottobosco delle faggete esaminate nell’alta valle del Sangro sembra essere legata al ciclo del carbonio. In particolare sembra di poter distinguere due assi di variazione: da una parte una differenziazione tra specie nettamente nemorali e sciafile, tipiche di boschi freschi e produttivi, e specie di boschi chiari o addirittura di margine; dall’altra una differenziazione tra specie tipiche delle faggete e un moderato numero di specie di fasce vegetazionali inferiori su amphimull spesso. Queste differenziazioni potrebbero rispecchiare una variabilità locale naturale oppure potrebbero essere legate a un maggiore o minore disturbo (anche in epoche relativamente lontane, vista la lentezza con cui si ricostituiscono i suoli in ambienti mediterranei), o ancora potrebbe esserci un’interazione tra variabilità naturale e disturbo antropico ([27])
Va inoltre sottolineato che in queste faggete, al limite della loro ottimalità ecologica, caratterizzate da una marcata povertà floristica di alcune parcelle, anche le misure gestionali sostenibili adottate, che in altri contesti geografici risultano funzionali alla conservazione della biodiversità, in quest’area possono produrre impatti imprevisti. La presenza dell’associazione Digitalidi-Fagetum, aspetto povero di questa vegetazione, che prende il sopravvento nelle faggete degradate, ne è infatti indicatrice. In questo contesto, l’individuazione di ecoindicatori sensibili che funzionino a livello di micro-scala, si rivelano particolarmente utili per calibrare e selezionare interventi selvicolturali che possano indirizzare l’ecosistema verso processi di rinaturazione.
Sviluppi futuri
Questo studio apre una serie di possibilità di ricerca sia teorica che applicativa:
- l’individuazione di differenziazioni non facilmente riconoscibili a un primo esame nella composizione della flora erbacea, e la dimostrazione della loro correlazione con alcuni fattori edafici, può permettere di elaborare un inventario funzionale, anche cartografico, delle singole parcelle. Conoscendo infatti i parametri edafici, è possibile prevedere quali specie erbacee siano potenzialmente presenti e, viceversa, osservando la presenza di determinate specie erbacee, si può stabilire quali siano le caratteristiche pedologiche attese (più probabili);
- è possibile impostare studi per verificare se nella distribuzione delle specie erbacee, per esempio delle Orchidaceae, non siano presenti limitazioni nella dispersione, o se invece la variabilità nella distribuzione delle specie non sia dovuta interamente ai fattori dell’habitat;
- il fatto che i parametri individuati sembrano legati al ciclo del carbonio stimola future ricerche sulle caratteristiche precipue del ciclo del carbonio (e a quelli collegati dell’acqua e dei nutrienti) nelle faggete centroappenniniche; questo aspetto è strettamente legato alla produttività delle faggete e influenza profondamente la sostenibilità delle misure di gestione così come anche la potenzialità per le specie animali della foresta.
Ringraziamenti
Si ringraziano la Dott.ssa G. Dowgiallo per averci ospitato nel laboratorio di pedologia del Dipartimento di Biologia Vegetale dell’Università di Roma La Sapienza; il Dr. D. Cicuzza che ha iniziato gli studi su forme di humus e vegetazione nel territorio oggetto dell’ indagine, M. Consalvo per la revisione critica di una prima stesura di questo lavoro, A. Cardillo, A. Di Benedetto e C. Sulli per aver collaborato attivamente al lavoro consentendoci di effettuare i rilevamenti nella Riserva Biogenetica della Biosfera “Monte di Mezzo” e nel Parco Nazionale d’Abruzzo (SIC Feudo Intramonti e Monte Godi). Studio cofinanziato dal progetto Life04NAT/IT/000190, “Tutela dei siti Natura 2000 gestiti dal Corpo Forestale dello Stato”.
References
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar