Structure of an old-growth stand (Reserve of Lom, Republic of Bosnia Herzegovina) and two over-mature forest stands from the Italian eastern Alps (Ludrin, TN, and Val Navarza, UD)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 5, Pages 100-111 (2008)
doi: https://doi.org/10.3832/efor0512-0050100
Published: Mar 26, 2008 - Copyright © 2008 SISEF
Research Articles
Abstract
The Republic of Bosnia Herzegovina is a country relatively rich (2000 ha) of primeval and old-growth forests that are very rare in central and southern Europe. One of the best Bosnian old-growth is located in the forest of Lom where a Forest reserve of 278 ha was established in 1956. Inside this reserve, 1 ha long-term intensive monitoring plot was established in 2005 to investigate the origin, spatio-temporal development and disturbance history of the stand. Structural characteristics of the stand were than compared with other Bosnian old-growth and with two well-preserved Italian forests belonging to the same forest type. The permanent plot is very rich in living biomass (1158 m3 ha-1) and in coarse woody debris (383 m3 ha-1 approximately 33% of living trees). The oldest trees are more than 450 years old. The stand has developed without large-scale disturbances and, according to data coming from other Bosnian old-growth, appears to be highly stable. The main differences between a true old-growth (Lom) and the previously managed forests of Ludrin and Navarza are the quality and the quantity of coarse woody debris, the size of the largest trees and the age of the oldest trees. The importance of Forest Reserves as a reference for managed forest and as a site for long-term ecological research is discussed.
Keywords
Old-growth forest, Natural reserve, Forest structure, Silviculture
Introduzione
La ricerca ecologico-forestale nelle riserve forestali ha una lunga tradizione sia in Europa ([46]), sia in America settentrionale ([49]) ma solo negli ultimi decenni i ricercatori hanno acquisito una maggiore consapevolezza dell’importanza della ricerca di lungo periodo. La raccolta di serie di dati di lungo periodo provenienti da diversi ecosistemi e la disponibilità di mezzi di prelievo ed analisi che non erano disponibili nel passato ha recentemente portato a dei radicali cambiamenti di alcuni postulati della “saggezza tradizionale” in campo ecologico (p. es., Stevens in Oliver & Larson 1996) rimettendo in discussione strategie di gestione e conservazione delle risorse naturali ed evidenziando la necessità di aumentare il numero delle aree di monitoraggio di lungo periodo (LTER, Long Term Ecological Research), di aree sperimentali, di studi retrospettivi e di modelli di previsione più accurati ([17], [22]). Questo tipo di studi, necessitando di aree indisturbate dall’uomo e di sicurezza sulla destinazione d’uso futura di queste, è preferibilmente svolto all’interno di Parchi, aree protette o di Riserve forestali.
Le riserve forestali sono dei comprensori forestali nei quali le attività selvicolturali sono escluse e la foresta viene lasciata evolvere naturalmente, in assenza dell’intervento diretto dell’uomo. Le riserve forestali sono presenti in tutti i continenti anche se ogni stato, o regione, ha una sua definizione e un suo regime di tutela (Parviainen & [8]). Le Riserve forestali svolgono diverse funzioni: sono riserve biogenetiche, rappresentano aree prioritarie di ricerca ecologico-forestale, sono poli di educazione ambientale e hanno dei valori culturali intrinseci che meritano di essere conservati e valorizzati ([15], [18], [32], [42]).
Le riserve forestali sono preferibilmente istituite in foreste primarie indisturbate dall’attività dell’uomo, ma questo aspetto è di difficile applicazione in Europa centro-meridionale dove la quasi totalità delle foreste è stata, direttamente od indirettamente, soggetta a delle modifiche di struttura e di composizione da parte dell’uomo ([40]). Nelle foreste temperate europee le riserve forestali sono quindi prevalentemente istituite in popolamenti forestali un tempo coltivati ma che hanno potuto evolversi naturalmente per un periodo di tempo abbastanza lungo acquisendo caratteristiche di naturalità ([42]).
All’interno delle riserve forestali sono particolarmente importanti i popolamenti che hanno raggiunto le fasi di sviluppo terminali della dinamica forestale, quelle caratterizzate dalla presenza di alberi vecchi e di grosse dimensioni, di legno morto in piedi ed a terra (coarse woody debris o CWD) e da una struttura pluristratificata. Queste fasi prendono il nome di old-growth ([16], [18], [29], [33]) o di foreste vetuste ([7]).
In Italia ([33], [44]) e nelle Alpi ([31], [48]) ci sono pochi popolamenti che hanno le caratteristiche strutturali di foreste vetuste mentre nella fascia temperata del continente europeo i migliori esempi di foreste vetuste si trovano nei Carpazi e nella penisola balcanica ([8], [21], [24], [36], [38], [40], [52]) e, in particolare, nella Repubblica di Bosnia Erzegovina ([43]). La Bosnia Erzegovina é stata oggetto da molti decenni dell’interesse dei forestali dell’Europa occidentale proprio per la presenza di estesi lembi di foreste naturali o semi-naturali che hanno una composizione e delle caratteristiche ecologiche-strutturali simili alle foreste presenti nelle Alpi ed in Europa centrale ([20], [26], [50]).
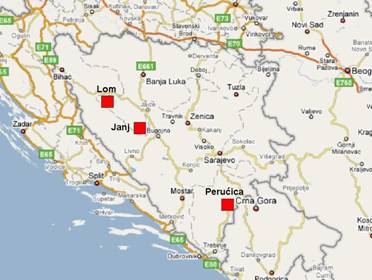
La Bosnia Erzegovina ha una copertura forestale del 43.1%. A causa della morfologia del territorio, delle vicende storiche, che hanno visto susseguirsi diversi tipi di dominazione e di conflitti, e della lontananza dalle principali vie di comunicazione della penisola balcanica, ancora all’inizio del XX secolo oltre il 50% delle foreste bosniache erano foreste vergini ([53]). Nell’ultimo secolo si é verificato un intenso sfruttamento delle risorse forestali ed attualmente solo lo 0.1% delle foreste bosniache (pari a circa 2000 ha) può essere considerato foresta primaria ([10]). Le più importanti di queste foreste primarie sono protette da tre Riserve forestali: Perućica (inserita nel Parco nazionale Sutjeska), Janj e Lom (Fig. 1, Tab. 1).
Fig. 1 - Mappa della Bosnia con la localizzazione delle tre più importanti foreste vergini.
Tab. 1 - Foreste vergini della Bosnia.
| Riserva forestale |
Comune | Anno istituzione |
Superficie [ha] | Settore altitudinale [m] |
|---|---|---|---|---|
| Lom | Drinic | 1956 | 298 | 1250-1522 |
| Janj | Sipovo | 1954 | 295 | 1180-1510 |
| Perućica | Foča | 1952 | 1434 | 500-2387 |
Le finalità di questo lavoro sono quelle di analizzare le caratteristiche strutturali di un popolamento appartenente agli stadi più maturi della successione (avente quindi caratteristiche di vetustà) nella riserva di Lom ed effettuare un primo confronto con due popolamenti delle Alpi orientali italiane appartenenti alla stessa categoria forestale identificati dalle rispettive amministrazioni (Provincia Autonoma di Trento e Regione Autonoma Friuli Venezia Giulia) come riserve forestali per il loro elevato grado di naturalità (assenza di interventi antropici negli ultimi decenni). Nello stesso tempo lo studio è stata l’occasione per insediare un’area di studio permanente (LTFER) all’interno della riserva forestale di Lom. Questa riserva forestale è stata oggetto di ricerca da diversi decenni ([9], [53]) e, nel 2001 ([30]) è stata dotata di un piano di gestione grazie ad un contributo della World Bank - The International Development Association (IDA). La foresta si trova sull’attuale confine tra la Repubblica Srpska e la Confederazione Croato-musulmana, le due entità che secondo gli accordi di Dayton costituiscono attualmente la Repubblica di Bosnia Erzegovina, ma è stata risparmiata, a differenza di altre foreste primarie come quella di Jani, da danni e dall’insediamento di campi minati durante le recenti guerre che hanno interessato il territorio della ex-Jugoslavia ([45]).
Materiali e metodi
Descrizione dell’area di studio
La riserva forestale di Lom (Fig. 2) ha una superficie di 297.7 ha ed è situata nel comune di Drinic ad una latitudine nord compresa tra 44° 27´ e 44° 28´ ed una longitudine est compresa tra 16° 27´ e 16° 30’.
Fig. 2 - La foresta di Lom con le caratteristiche più evidenti dei popolamenti old-growth: struttura pluristratificata, grossi diametri ed abbondante coarse-woody debris.

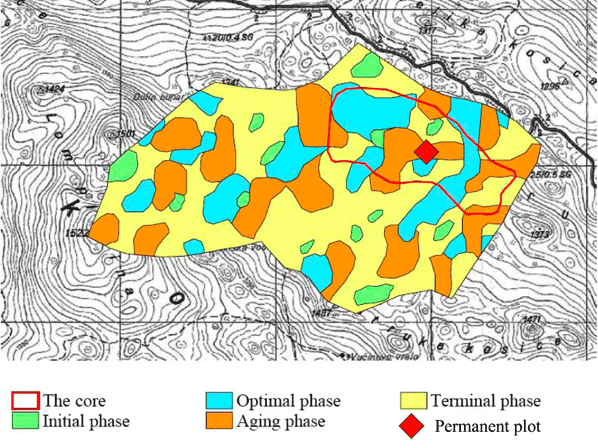
La riserva é localizzata sulle pendici del Monte Klekovaca ad una altitudine compresa tra 1250 e 1522 m s.l.m. Il substrato è calcareo e la morfologia carsica. Le precipitazioni annuali sono mediamente di circa 1600 mm. La riserva è suddivisa tra un’area di protezione assoluta (core area) avente una superficie di 55.8 ha ed una superficie più esterna di rispetto (buffer zone). La foresta appartiene all’Azienda Forestale Pubblica Srpske šume della municipalità di Drinic. La vegetazione ([5]) è rappresentata per il 75% della superficie da Piceo-Abieti-Fagetum illyricum, per il 18% da Aceri-Fagetum subalpinum e per la restante parte da Abieti - Piceetum (Fig. 3). Da un punto di vista strutturale la riserva è caratterizzata da un’alternanza, su ampie superfici, di fasi ottimali, stramature e di crollo con una presenza limitata delle fasi più giovanili (Fig. 4).
Fig. 3 - Caratterizzazione vegetazionale della riserva forestale di Lom ([30])
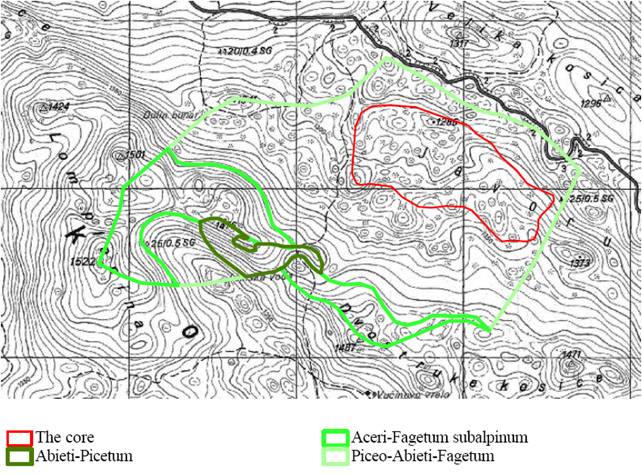
Fig. 4 - Fasi di sviluppo dei popolamenti forestali all’interno della riserva forestale di Lom ([30]).
Metodi
L’inventario effettuato nel 2001 ha evidenziato un volume medio ad ettaro di 727.8 m3 di cui il 68.4 % (497.7 m3) di conifere e la restante parte di (231.0 m3) di latifoglie ([30]). Nel corso dello stesso inventario è stata effettuata una prima sommaria classificazione dei popolamenti forestali nelle diverse fasi di sviluppo utilizzando la metodologia di Korpel ([23]). Tra le conifere si è osservata una leggera prevalenza dell’abete bianco (Abies alba Mill.) rispetto all’abete rosso (Picea abies (L.) Karst.) mentre tra le latifoglie prevale nettamente il faggio (Fagus sylvatica L.) e, in misura nettamente inferiore, l’acero di monte (Acer pseudoplatanus L.).
Nell’ottobre 2005, all’interno della core area in un popolamento classificato come appartenente alla aging phase ([30]) è stata delimitata un’area avente una superficie di circa 1 ettaro (1.0855 ha). In quest’area sono stati mappati tutti gli alberi e gli elementi di necromassa mediante il rilievo, con la precisione del cm, delle coordinate polari (angolo azimutale e distanza orizzontale) a partire da una origine.
In tutti gli alberi (diametro a 130 cm di altezza > 7.5 cm) sono stati misurati, con la precisione del cm, i diametri a 50 e 130 cm di altezza dal suolo e, con la precisione di 10 cm, l’altezza totale e l’altezza di inserzione della chioma e quattro proiezioni della chioma al suolo lungo due assi cartesiani. La statura è stata calcolata mediando l’altezza dei 5 alberi dal diametro maggiore ([51]).
La necromassa (detriti legnosi grossolani o coarse woody debris, CWD) è stata rilevata sull’intera superficie dell’area suddivisa in tre distinte categorie ([34]): fusti a terra (logs), piante morte in piedi (snags) e ceppaie (stumps).
Per i fusti a terra sono stati misurati i diametri dei due estremi del fusto e la lunghezza. La soglia inferiore di rilievo è stata di 10 cm di diametro nel lato di diametro maggiore.
Delle ceppaie sono stati misurati il diametro superiore, il diametro inferiore della ceppaia al netto dei contrafforti e l’altezza della ceppaia. Una altezza di 130 cm è stata la soglia per distinguere le ceppaie (altezza < 130 cm) dai tronchi in piedi. La soglia inferiore di rilievo è stata di 7 cm di diametro nella sezione superiore.
Delle piante morte in piedi sono stati misurati il diametro a 130 cm da terra e l’altezza (intesa sia come altezza complessiva per gli individui integri, sia come altezza del fusto per gli alberi troncati).
Per tutti gli elementi del CWD é stato anche valutato il grado di decomposizione secondo una scala qualitativa di 4 (per le ceppaie) o 5 (per gli alberi morti in piedi e per i tronchi a terra) valori ([34]).
Il volume della componente arborea (alberi vivi e alberi morti in piedi) è stato stimato, per le tre più importanti specie, con l’ausilio di tavole locali (rispettivamente per abete bianco, abete rosso e faggio). La stima del volume della restante necromassa (tronchi a terra, ceppaie, fusti troncati) è avvenuta approssimando la forma di questi elementi a quella di un tronco di cono secondo l’equazione (eqn. 1):
dove V = volume, h = altezza o lunghezza, R = raggio maggiore e r = raggio minore.
L’approssimazione comporta una sottostima del volume reale della necromassa poiché si esclude il volume dei contrafforti e dell’apparato radicale superficiale.
Infine in ogni individuo mappato é stata prelevata una carota incrementale, mediante il succhiello di Pressler, al fine di determinare l’età.
Risultati
Il popolamento oggetto di studio è una fustaia mista di faggio, abete bianco ed abete rosso. La densità del popolamento forestale è di 617 alberi ad ettaro (Tab. 2, Fig. 5) di cui 377 faggi (61%), 165 abeti bianchi (27%) e 75 abeti rossi (12%).
Tab. 2 - Caratteristiche strutturali dell’area di studio della foresta di Lom
| Specie | Densità [n ha-1] |
Area basimetrica [m2ha-1] |
Diametro medio [cm] |
Altezza media [m] |
Volume [m3ha-1] |
|---|---|---|---|---|---|
| Abete bianco | 165 | 31.4 | 49.2 | 28.0 | 522.8 |
| Abete rosso | 75 | 23.6 | 63.0 | 33.3 | 430.5 |
| Faggio | 377 | 14.3 | 22.0 | 17.2 | 204.6 |
| Totale | 617 | 69.3 | - | - | 1157.9 |
Fig. 5 - Tessitura e composizione dell’area di monitoraggio permanente (chiome del piano dominante e posizione di tutti gli alberi all’interno del plot).
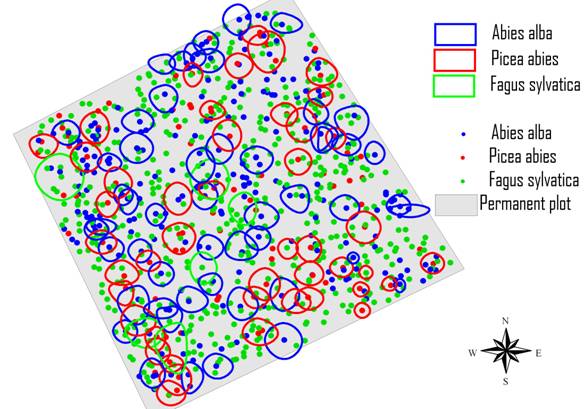
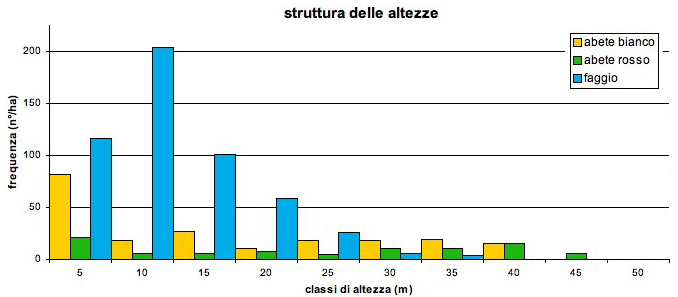
L’area basimetrica totale è di 69.3 m2 ha-1. Il volume complessivo è di 1157.9 m3 ha-1 di cui il 18% è di faggio, il 45% di abete bianco ed il 37% di abete rosso. La curva di distribuzione diametrica (Fig. 6) ha il tipico andamento esponenziale negativo dei popolamenti disetanei con una maggiore frequenza del faggio nelle classi medie e piccole. Sia l’abete bianco che quello rosso presentano i diametri maggiori con individui che raggiungono rispettivamente i 115 cm ed i 120 cm di diametro. La statura ([51]) del popolamento è di 44 m (Fig. 7). Il piano dominante e codominante costituito dagli alberi di altezza uguale o superiore ai 29 m ([25]) è costituito quasi esclusivamente da abete bianco (50.5%) ed abete rosso (43.7%) con presenza marginale di faggio (5.8%). Il piano intermedio (altezza compresa tra 20 e 29 m) ha una distribuzione delle specie più equilibrata ma con dominanza di faggio (60.3%) rispetto ad abete bianco (28.9%) ed abete rosso (10.7%); i piani dominati (altezza < 20 m) sono costituiti maggiormente dal faggio che rappresenta il 72.5% degli individui ed è di gran lunga più rappresentato dell’abete bianco (21.6 %) e dell’abete rosso (5.8%).
Fig. 6 - Curva di distribuzione dei diametri dell’area di monitoraggio permanente.
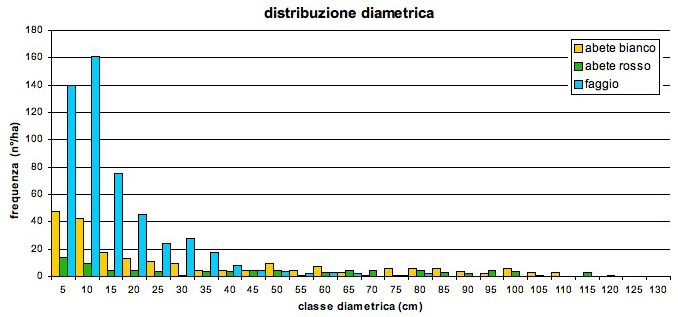
Fig. 7 - Struttura delle altezze dell’area di monitoraggio permanente.
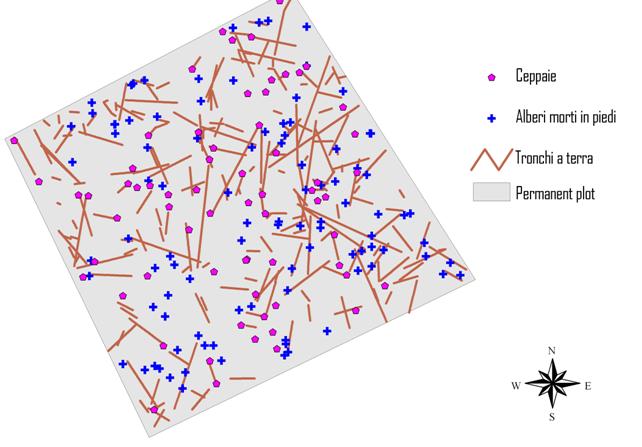
La necromassa (Fig. 8) presente nell’area ha un volume di 383 m3 ha-1, pari al 33% circa del volume cormometrico degli alberi vivi (Tab. 3). Numericamente la componente più frequente è rappresentata dai tronchi a terra (192 elementi ha-1) e secondariamente dalle piante morte in piedi (91 elementi) e dalle ceppaie (61 elementi). Dal punto di vista del volume prevalgono invece gli alberi morti in piedi (51% del volume) rispetto ai tronchi a terra (46 %) ed alle ceppaie (3 %).
Fig. 8 - Distribuzione spaziale della necromassa (ceppaie, alberi morti in piedi e tronchi a terra) all’interno dell’area permanente.
Tab. 3 - Necromassa dell’area di studio della foresta di Lom.
| Tipo | Volume [m3ha-1] |
Elementi | Classe 1 [m3ha-1] |
Classe 2 [m3ha-1] |
Classe 3 [m3ha-1] |
Classe 4 [m3ha-1] |
|---|---|---|---|---|---|---|
| Ceppaie | 11.1 | 61 | 0.0 | 0.0 | 10.6 | 0.5 |
| Tronchi a terra | 175.2 | 192 | 0.6 | 39.3 | 86.9 | 48.3 |
| Alberi morti in piedi | 197.0 | 91 | 52.1 | 52.6 | 90.1 | 2.2 |
| Totale | 383.3 | 344 | 52.7 | 92.0 | 187.6 | 51.1 |
In relazione allo stato di decomposizione della necromassa vi è una maggiore incidenza delle classi di decomposizione 3 e, in misura inferiore, 2 rispetto alle classi 1 e 4. La presenza marginale di tronchi a terra nella classe 1 è riconducibile al fatto che la maggior parte degli alberi morti iniziano il processo di decomposizione come elementi in piedi per poi arrivare a terra come tronchi già in uno stadio di decomposizione più avanzato.
Dalle prime analisi effettuate le età massime raggiunte dalle tre specie presenti sono circa 450 anni per l’abete bianco, 400 anni per l’abete rosso e circa 330 anni per il faggio.
Discussione
Il popolamento di Lom rappresenta un tipico esempio di foresta mista e disetanea del piano montano con tessitura per piede d’albero o per piccoli gruppi. Questa categoria forestale è quella maggiormente diffusa nelle foreste del piano montano dell’Europa centro-meridionale e, potenzialmente, è quella più diffusa sia nel versante settentrionale sia nel versante meridionale della catena alpina. La fase di sviluppo può essere definita di old-growth secondo la classificazione utilizzata in America settentrionale (p. es., Oliver & Larson 1996) oppure può essere definita fase stramatura o vetusta (aging phase) utilizzando la classificazione sviluppata in Europa centrale ([23], [27]). Questa fase è caratterizzata da elevata quantità di biomassa, elevata quantità di legno morto, singoli individui o gruppi di alberi di grandi dimensioni, alberi molto vecchi e struttura pluristratificata con presenza di nuclei di rinnovazione. Nell’ambito delle foreste primarie bosniache già segnalate da Tregubov ([53]) e Drinic ([9]), il popolamento studiato a Lom si caratterizza tra quelli che hanno il volume cormometrico più alto anche se, data la quota più elevata, Lom ha una fertilità stazionale inferiore rispetto a Janj e, soprattutto, a Perućica. La minore fertilità comporta però una maggiore longevità e, nell’insieme, una maggiore stratificazione e mescolanza delle tre specie presenti (Tab. 4).
Tab. 4 - Confronto l’area di monitoraggio permenente insediata a Lom e altre aree di studio situate in foreste vergini bosniache. (*) Fs: faggio, Aa: abete bianco, Pa: abete rosso. Le specie sono elencate in ordine di importanza numerica. (**) Al fine di confrontare i dati con le altre aree questi sono riferiti ad una soglia di cavallettamento di 10 cm a 130 cm di altezza.
| Foresta | Densità [n ha-1] |
Composizione* | Area basimetrica [m2ha-1] |
Volume [m3ha-1] |
Statura [m] |
Quota [m s.l.m.] |
Riferimento |
|---|---|---|---|---|---|---|---|
| Lom** | 488 | Fs, Aa, Pa | 68.6 | 1151 | 44 | 1300 | Questo lavoro |
| Lom | 330 | Fs, Aa, Pa | 50.9 | 717 | 41 | 1260 | [9] |
| Lom | 447 | Fs, Aa, Pa | 57.9 | 773 | 39 | 1340 | [9] |
| Janj | 386 | Pa, Fs, Aa | 65.9 | 893 | 40 | 1440 | [9] |
| Janj | 367 | Aa, Fs, Pa | 59.0 | 901 | 40 | 1340 | [9] |
| Janj | 360 | Pa, Aa, Fs | 61.6 | 873 | 43 | 1260 | [9] |
| Perućica | 408 | Aa, Pa, Fs | 87.1 | 1353 | 47 | 1150 | [9] |
| Perućica | 470 | Aa, Fs, Pa | 79.5 | 1111 | 48 | 1460 | [9] |
| Perućica | 434 | Aa, Pa, Fs | 63.3 | 933 | 42 | 1100 | [9] |
| Perućica | 535 | Aa, Fs, Pa | 77.3 | 1010 | 44 | 1440 | [9] |
I popolamenti descritti nelle diverse aree di studio bosniache appaiono molto simili dal punto di vista strutturale (in termini di composizione, distribuzione dei diametri, distribuzione delle altezze). Nell’intera riserva di Lom è evidente l’assenza di disturbi di grande estensione ed intensità (stand replacing disturbances) e la dinamica forestale appare guidata soprattutto da disturbi a scala di dettaglio che interessano il singolo albero o piccoli gruppi. Questo regime di disturbi è molto importante dal punto di vista selvicolturale, in quanto rappresenta un riferimento per il trattamento a taglio saltuario applicato nelle foreste coltivate, ma in natura è relativamente poco diffuso sia in Europa e sia in altri continenti ([39]). Il regime di disturbi di Lom è, ad esempio, diverso da quello osservato nella foresta di Perućica (Nagel, comunicazione personale), e da quelli osservati in foreste vetuste slovene ed in altri paesi centro-europei ([3], [36], [37]) dove la frequenza, l’intensità e l’estensione dei disturbi, soprattutto gli schianti provocati dal vento, appaiono maggiori.
Il confronto tra l’area permanente della foresta di Lom e due aree permanenti recentemente insediate nella Provincia Autonoma di Trento (Foresta di Ludrin nel Comune di San Lorenzo in Banale) e nella Regione Autonoma Friuli Venezia Giulia (Foresta di Navarza nel Comune di Ampezzo) permette di evidenziare le differenze esistenti tra i popolamenti più maturi e rappresentativi presenti attualmente nel versante meridionale delle Alpi e foreste vergini appartenenti alla stessa categoria forestale (Tab. 5).
Tab. 5 - Confronto tra la foresta di Lom e le foreste di Ludrin e Navarza. *Fs: faggio, Aa: abete bianco, Pa: abete rosso. Le specie sono elencate in ordine di importanza numerica.
| Foresta | Superficie [m2] |
Densità [n ha-1] |
Composizione* | Area basimetrica [m2ha-1] |
Volume [m3ha-1] |
CWD [m3ha-1] |
Statura [m] |
Età degli alberi più vecchi [anni] |
|---|---|---|---|---|---|---|---|---|
| Lom | 10085 | 617 | Fs, Aa, Pa | 69.2 | 1157.9 | 383.3 | 44 | 450 (Aa) |
| Navarza (UD) | 5052 | 500 | Fs, Aa, Pa | 55.6 | 841 | 90.1 | 40 | 150 (Aa) |
| Ludrin (TN) | 12800 | 407 | Aa, Pa, Fs | 49.7 | 678 | 172.3 | 42 | 180 (Aa) |
Entrambi i popolamenti di Ludrin e Navarza sono stati nel passato coltivati dall’uomo ma hanno potuto evolversi in modo naturale per diversi decenni. Le tracce dell’attività dell’uomo sono ancora evidenti (ad esempio le ceppaie delle vecchie utilizzazioni) ma, nello stesso tempo, i due popolamenti hanno sviluppato alcune caratteristiche delle fasi di sviluppo più mature (o dei popolamenti classificabili come vetusti o old-growth). Le maggiori differenze tra queste foreste ed un popolamento di riferimento come quello di Lom consistono soprattutto nella quantità e nella qualità della necromassa, nelle dimensioni raggiunte dagli alberi più grossi e nell’età degli alberi più vecchi. Nonostante le differenze geografiche, climatiche e stazionali esistono delle evidenti similitudini tra i popolamenti dell’Italia nord-orientale e gli analoghi popolamenti bosniaci come ad esempio la distribuzione dei diametri, la struttura delle altezze ed i valori di statura (che evidenziano le potenzialità dei popolamenti italiani). Nonostante il relativamente breve periodo in cui i popolamenti italiani sono stati lasciati evolvere naturalmente (pochi decenni) sia il volume degli alberi viventi e sia la necromassa hanno già raggiunto valori molto importanti evidenziando una dinamica molto vivace e diversa da quella osservata in popolamenti vetusti del piano subalpino delle Alpi italiane caratterizzati da un dinamismo molto più lento ([34], [35]). Sia i popolamenti italiani, sia la foresta di Lom rappresentano esempi della massima maturità ed evoluzione strutturale raggiunta da popolamenti forestali in un determinato settore geografico e possono costituire degli importanti punti di riferimento per lo studio della dinamica dei popolamenti forestali e per il confronto con popolamenti coltivati.
L’importanza delle riserve forestali, ed in particolare delle fasi di sviluppo più mature, come riferimento per le foreste coltivate e per i modelli selvicolturali è stata ampiamente discussa in Italia ([50], [51]), in Europa centro-settentrionale ([26], [28], [47]) ed in America settentrionale ([13], [15]).
Dal punto di vista scientifico le riserve forestali rappresentano quindi dei siti privilegiati per la ricerca selvicolturale e per le ricerche di lungo periodo in quanto in esse è possibile predisporre dei monitoraggi di lungo periodo in assenza di disturbo diretto da parte dell’uomo. Solo attraverso questi studi è possibile acquisire informazioni fondamentali sui processi e sulle dinamiche che avvengono nei popolamenti forestali e quindi elaborare modelli e mettere a disposizione riferimenti per una gestione selvicolturale su basi naturalistiche ([1], [2], [4], [13], [14]). In questi ultimi decenni la disponibilità di popolamenti forestali primari, o che hanno potuto evolversi naturalmente per un lungo periodo di tempo ed hanno caratteristiche di naturalità, ha acquisito una grandissima importanza anche alla luce del climate change in quanto in essi è possibile osservare gli effetti dei cambiamenti climatici sui processi che avvengono in foresta con una prospettiva di lungo periodo ed in assenza di interventi da parte dell’uomo ([11], [54]).
Infine, oltre all’indiscusso valore scientifico e naturalistico, gli ultimi lembi di foreste vergini del continente europeo devono essere assolutamente protetti per il loro enorme ed insostituibile valore culturale e paesaggistico ([6], [12], [19], [42]). Le foreste vergini, e soprattutto i popolamenti che sono nelle fasi più mature e nelle fasi di crollo, fanno parte del nostro immaginario collettivo perché, grazie alla presenza di alberi plurisecolari di grandi dimensioni ed alla presenza abbondante di alberi morti in piedi ed a terra, forniscono una immagine della natura primigenia che dominava il nostro continente prima della massiccia antropizzazione. Le emozioni ed i sentimenti evocati da queste foreste rappresentano un valore culturale che abbiamo il dovere di conservare, valorizzare e trasmettere alle generazioni future.
Ringraziamenti
Il lavoro è stato svolto con il contributo della Provincia Autonoma di Trento (Foresta di Ludrin nel Comune di San Lorenzo in Banale) e della Regione Autonoma Friuli Venezia Giulia (Foresta di Navarza nel Comune di Ampezzo).
Si ringraziano i Prof. Midhat Uscuplic e Pietro Piussi per averci fatto conoscere la riserva di Lom e per averci messo a disposizione la loro grande esperienza in riserve forestali, foreste vetuste ed in aree di monitoraggio di lungo periodo.
Si ringrazia infine la Faculty of Forestry, University of Banja Luka (Republic of Srpska, Republic of Bosnia Herzegovina) per il supporto logistico.
References
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar