Relationships between silvicultural system, forest type and floristic diversity in chestnut coppices
Forest@ - Journal of Silviculture and Forest Ecology, Volume 5, Pages 136-150 (2008)
doi: https://doi.org/10.3832/efor0521-0050136
Published: May 21, 2008 - Copyright © 2008 SISEF
Research Articles
Abstract
Chestnut stands are among the main forest formations in Italy. The aim of this work is to assess the floristic diversity of chestnut coppice stands of Sabatini Mts. (central Italy), highlighting the relationships between the silvicultural system and the ecological processes. In the management of these coppices almost exclusive importance has been given to the wood production, rating the stands on the basis of the rotation that maximizes the production of the most valuable assortments. The assessment carried out is based on floristic and dendrometric surveys within permanent circular plots (10 meters radius) randomly selected within three chronological strata. Surveys have been realized on a total of 15 plots, 5 for each stratum. Indices of floristic richness and diversity have been elaborated to point out variations of forest types and to suggest possible alternatives to the current management to mitigate its impact on biodiversity.
Keywords
Castagneti, ceduo, diversità floristica, gestione, tipo forestale
Introduzione
Il castagno (Castanea sativa) è una delle specie forestali più importanti per il territorio europeo e da tempi remoti ha avuto una ampia diffusione al di fuori dell’area di indignato. In Italia la sua attuale distribuzione (circa 790000 ha secondo [20]) è il risultato dell’azione dell’uomo che lo ha favorito a scapito di formazioni naturali quali querceti di rovere, cerro, roverella o faggete ([8]), soprattutto nella forma di governo a ceduo (oltre 600000 ha secondo [20]).
Sui rilievi vulcanici pre-appenninici del Lazio il castagno trova un ambiente climatico e pedologico ottimale ed è in grado di manifestare pienamente la sua elevata capacità di accrescimento, che viene esaltata dal governo a ceduo. Sui Monti Sabatini, nel territorio di Bracciano, Manziana, Oriolo Romano, Bassano Romano e Sutri sono presenti oltre 2000 ha di cedui matricinati di castagno, che rappresentano un’importante risorsa economica per molti proprietari, sia pubblici (circa il 20% del totale) che privati.
Il territorio è rinomato dal punto di vista turistico-ricreativo e naturalistico ed è inserito nel Parco Naturale Regionale di Bracciano-Martignano e in alcune aree della Rete Natura 2000. Gli obiettivi perseguiti tramite queste aree protette, cioè la conservazione di habitat e di specie animali e vegetali di significativo valore naturalistico, spesso si scontrano con le aspettative strettamente economiche che i proprietari forestali hanno nei confronti dell’utilizzo del ceduo castanile.
La sostenibilità ambientale della gestione di questi soprassuoli non ha avuto finora adeguata considerazione. È quasi sempre stata ritenuta prioritaria la sostenibilità economica, assicurata dall’applicazione di un turno tecnico che consentisse di ottenere: assortimenti da opera di ottima qualità, elevata estensione delle tagliate a fine turno, coetaneità e monospecificità dei popolamenti.
Il ceduo matricinato, sebbene garantisca all’ecosistema forestale una buona capacità di ripresa a seguito delle perturbazioni apportate con il taglio a fine turno, è considerato un tipo di trattamento con marcato impatto ambientale e limitato valore naturalistico ([11]). Gli studi finora condotti portano a conclusioni parzialmente contrastanti sulla capacità del ceduo matricinato di mantenere un elevato livello di biodiversità. Per quanto riguarda la flora superiore, le ricerche effettuate dimostrano che il disturbo dovuto alla ceduazione aumenta la numerosità delle specie presenti, ma diminuisce la naturalità della compagine vegetale che necessita di tempi lunghi per ricostituirsi sufficientemente (ad es., [34], [17], [24], [14], [16], [33], [18], [25]). Età del ceduo e trattamento selvicolturale sembrano i fattori più determinanti l’evoluzione della diversità floristica dopo la ceduazione, anche più delle caratteristiche del suolo. Se è assodato che la ricchezza di specie è massima nei primissimi anni del nuovo ciclo per poi declinare abbastanza rapidamente con l’età ([5], [33], [25]), meno studiato è stato il ruolo che il regime di diradamento applicato può avere, avendo questo il significato di un disturbo di minore intensità rispetto alla ceduazione. Nonostante l’importanza assunta dalla conservazione della biodiversità forestale, nel nostro Paese sono ancora relativamente limitate le indagini sperimentali in merito all’influenza dei trattamenti selvicolturali su questa funzione.
Pertanto, si è ritenuto opportuno avviare un’azione di monitoraggio nell’area dei Monti Sabatini al fine di trarre indicazioni utili per la selvicoltura dei cedui castanili dei rilievi vulcanici laziali. L’obiettivo specifico di questo contributo è una prima valutazione quantitativa e qualitativa della diversità floristica di alcuni cedui castanili nella seconda metà del ciclo colturale, considerando sia il numero di specie presenti, sia le variazioni del corteggio floristico in funzione dell’età del ceduo, dei trattamenti selvicolturali applicati e delle caratteristiche stazionali dell’area di studio. Scopo finale della ricerca, che prevede un approfondimento e uno sviluppo nei prossimi anni, è di evidenziare possibili relazioni tra gestione selvicolturale e parametri floristici, tentando di individuare non solo i fattori di criticità, ma anche gli aspetti positivi, attuali e potenziali, insiti nel governo a ceduo, e di suggerire possibili misure correttive allo scopo di mitigare l’impatto sulla biodiversità vegetale.
Area di studio
I cedui castanili oggetto di studio sono localizzati nel comprensorio dei Monti Sabatini, in particolare all’interno dell’azienda agricola Vicarello, nel comune di Bracciano (RM), e in località Faggeta, nel comune di Oriolo Romano (VT); in entrambi i casi si tratta di proprietà private. Il territorio rientra nel Parco Naturale di Bracciano - Martignano e nei seguenti siti Natura 2000: SIC IT 6010034 “Faggete di Monte Raschio e Oriolo”; ZPS IT6030085 “Comprensorio Bracciano Martignano”; ZPS IT 6030005 “Comprensorio Tolfetano-Cerite-Manziate”.
Le formazioni forestali dell’area di studio possono essere suddivise in: faggete termofile; boschi misti caratterizzati dalla compenetrazione di elementi di querceti e faggeti; boschi cedui di castagno con sottobosco caratterizzato da un contingente di specie mesofile e della faggeta. Le faggete e i boschi misti dell’area sono per lo più ascrivibili all’alleanza Doronico-Fagion Ubaldi et al. 1987, con ingressioni di elementi acidofili dei Quercetalia robori-petraeae Tx. (1931) 1937 ([10]).
Nel settore nord-occidentale dell’apparato vulcanico Sabatino si ritrovano principalmente suoli di natura piroclastica, andosuoli e suoli bruni acidi su letti di scorie e ceneri vulcaniche ([22]). In base alla carta del fitoclima del Lazio ([9]), l’area di studio rientra interamente nel termotipo mesomediterraneo medio/termocollinare, ombrotipo subumido superiore/umido inferiore, regione xerotermica (sottoregione mesomediterranea).
Materiali e metodi
Generalità
Il protocollo sperimentale ha previsto i seguenti passi:
- delimitazione di tre strati cronologicamente differenti (A = metà turno, B = fine turno, C = oltre turno) in condizioni macroambientali tra loro omogenee, in un contesto ecologico intermedio tra quello delle faggete termofile a bassa quota e quello dei querceti mesofili (vedi sezione Risultati);
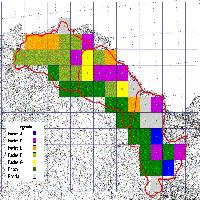
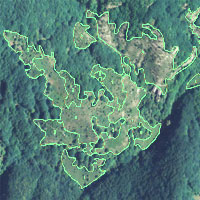
- selezione casuale di 5 punti di sondaggio in ciascuno strato (campionamento casuale stratificato - [12]), per un totale di 15 punti (Fig. 1);
- rilevamento dendrometrico e floristico all’interno di aree di saggio permanenti (forma circolare, raggio pari a 10 metri) centrate sui punti di sondaggio selezionati (Fig. 2 e Fig. 3);
- elaborazione dei dati dendrometrici;
- elaborazione dei dati floristici (confronto dei contingenti fitosociologici e degli spettri biologici e corologici degli strati, indici di biodiversità, analisi degli indici di Ellenberg, classificazione e ordinamento delle aree di rilievo sulla base della composizione floristica);
- validazione statistica dei risultati.
Fig. 1 - Punti di sondaggio e strati cronologici nell’area di studio.
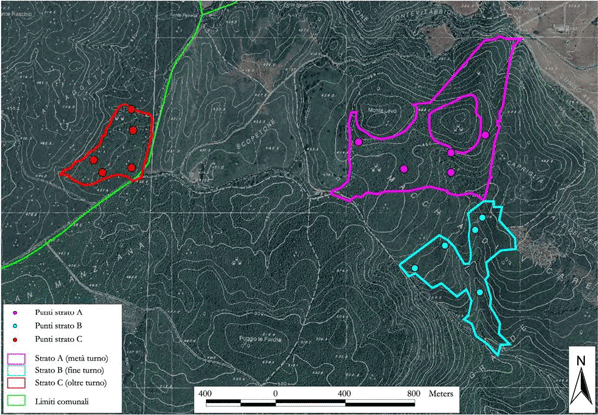
Fig. 2 - Centro di un’area di saggio (indicato dal picchetto) nello strato C.

Fig. 3 - Ceppaie esterne a un’area di saggio nello strato A.

Rilievi
Ai fini di questo lavoro sono state individuate tre aree a ceduo (strati) della stessa fisionomia (castagno), caratterizzate da condizioni similari per quanto riguarda gli aspetti fitoclimatici ([9]) e geologici ([22]), ma differenziate per età del ceduo. Gli strati sono indicati con le seguenti sigle:
- Strato A = metà turno: cedui a metà turno in località Vicarello - Bracciano (RM), che saranno interessati da taglio intercalare nel biennio 2006/2008 (età: 10-12 anni);
- Strato B = fine turno: cedui a fine turno in località Vicarello - Bracciano (RM), che saranno interessati dal taglio di fine turno nel biennio 2006/2008 e che sono stati oggetto di tagli intercalari nel biennio 2002/2004 (età: 17 anni);
- Strato C = oltre turno: cedui oltre turno in località Faggeta - Oriolo Romano (VT), non oggetto di diradamento e la cui ultima utilizzazione risale all’annata silvana 1988/1989 (età: 19 anni).
Gli strati A e B fanno parte di una compresa di cedui castanili e misti di estensione pari a 242.31 ha. Lo strato C è, invece, costituito da una piccola proprietà di circa 35 ha che si trova all’interno del SIC “Faggeta di Monte Raschio” e direttamente confinante con la faggeta stessa. I tre strati distano tra loro meno di 2000 metri (Fig. 1).
Le principali caratteristiche stazionali e cronologiche di ciascuna area di saggio sono riportate in Tab. 1.
Tab. 1 - Caratteristiche cronologiche e stazionali dei punti di sondaggio.
| Punto | Età (anni) |
Pendenza (%) | Quota (m slm) |
Esp. |
|---|---|---|---|---|
| A1 | 12 | 50 | 430 | NE |
| A2 | 12 | 17 | 435 | N |
| A3 | 10 | 25 | 450 | O |
| A4 | 12 | 8 | 450 | N |
| A5 | 10 | 8 | 412 | SE |
| B1 | 17 | 17 | 405 | NE |
| B2 | 17 | 17 | 400 | NO |
| B3 | 17 | 10 | 375 | S |
| B4 | 17 | 25 | 330 | S |
| B5 | 17 | 25 | 390 | O |
| C1 | 19 | 5 | 430 | S |
| C2 | 19 | 7 | 430 | NE |
| C3 | 19 | 5 | 430 | S |
| C4 | 19 | 7 | 420 | N |
| C5 | 19 | 5 | 435 | NO |
Il centro dell’area di saggio è stato reso permanente mediante un picchetto conficcato nel terreno, registrando in DGPS-post-processing la posizione topografica con precisione submetrica. All’interno di ciascuna area di saggio è stato eseguito il cavallettamento totale con soglia minima di rilevamento pari a 1 cm per il castagno e a 5 cm per tutte le altre specie arboree. Sono stati inoltre rilevati: il numero di matricine; il numero di fusti arborei, suddivisi tra vivi e morti; il numero di ceppaie. Sono stati successivamente determinati: il numero medio di polloni per ceppaia; il diametro medio dei fusti arborei; l’area basimetrica, suddivisa in fusti vivi e morti; l’incremento medio di area basimetrica.
Per quanto riguarda i rilievi floristici è stata registrata la presenza di tutte le specie di piante vascolari all’interno delle aree di saggio, indicando se la specie rinvenuta appartenesse allo strato erbaceo, arbustivo o arboreo. Sono state annotate anche alcune specie significative rinvenute subito al di fuori delle aree di saggio.
I rilievi floristici sono stati svolti periodicamente, con cadenza da quindicinale a mensile tra luglio del 2006 e giugno del 2007 per coprire i differenti periodi di fioritura delle diverse specie, soprattutto per quanto riguarda la componente erbacea. La determinazione delle specie raccolte è stata basata principalmente su Pignatti ([27]) e, per alcuni taxa, su Tutin et al. ([35]). Per la nomenclatura delle specie rinvenute e indicate nel testo si fa sempre riferimento a Pignatti ([27]).
I campioni raccolti sono stati essiccati e conservati (Fig. 4) presso l’Erbario della Tuscia. A partire dai dati raccolti sono stati calcolati gli spettri biologici e corologici normali e ponderati ed è stata creata una matrice presenza-assenza delle specie censite, ordinata per frequenze e per strato cronologico.
Fig. 4 - Campione di Scilla bifolia (n. 25762) conservato presso l’Erbario della Tuscia.

Analisi floristica
Allo scopo di evidenziare eventuali differenze fitogeografiche nella composizione floristica dei singoli strati, la matrice “specie x rilievi” è stata organizzata in chiave ecologica, distinguendo le specie attribuibili a ciascuno dei seguenti contingenti: Fagetalia sylvaticae Pawl. 1928; Quercetalia pubescenti-petraeae Klika 1933, Querco-Fagetea Br.-Bl. & Vlieger in Vlieger 1937, Quercetea ilicis Br.-Bl. 1947; specie di ambienti prativi; specie nitrofile di suoli eutrofici. Diversamente da studi simili ([33]), non è stato ritenuto opportuno utilizzare le cosiddette ancient forest species evidenziate per discriminare siti che hanno mantenuto a lungo caratteristiche forestali in termini di storia dell’uso del suolo (indipendentemente dal tipo di gestione selvicolturale) rispetto alle foreste secondariamente insediatesi in seguito all’abbandono dell’uso agricolo del suolo ([19], [7]), e non per discriminare differenti tipi di gestione selvicolturale.
Sono stati inoltre calcolati gli spettri biologici e corologici per ciascuno strato, seguendo Pignatti ([27]) per l’attribuzione delle specie alle categorie; sono stati calcolati sia gli spettri normali (percentuale del numero di specie appartenenti a ciascuna categoria rispetto al numero totale di specie rinvenute nello strato) sia gli spettri ponderati sulle frequenze (percentuale del numero di ritrovamenti delle specie appartenenti a ciascuna categoria rispetto al numero totale di celle con valore 1 nella matrice binaria “specie x rilievi” di uno strato).
Analisi della diversità
La ricchezza di specie (species richness) viene considerata tra i più importanti parametri per caratterizzare la diversità di una comunità e si identifica con la α-diversità, ovvero il numero di specie censite in un determinato territorio ([33]).
A partire dal numero di specie presenti è stata calcolata la β diversità, intesa come diversità sia intra-strato che tra i vari strati cronologici. La β diversità può essere considerata come la diversità tra punti di campionamento distribuiti lungo un transetto o un gradiente ambientale, riflettendo quindi i cambiamenti biotici e la diversità tra le specie ([23]), ed è calcolata con la formula (indice di Whittaker - eqn. 1):
dove, nel caso della diversità intra-strato: γ = numero totale di specie ritrovate nello strato; α = media del numero di specie in ogni punto dello strato; mentre, nel caso della diversità inter-strato: γ = numero totale di specie in tutti gli strati considerati; α = media del numero di specie in ciascuno strato.
Questa formula presenta lo svantaggio di fornire un valore di β diversità adimensionale, quantificato in maniera non commensurata rispetto alla γ e alla α diversità. Per risolvere questo inconveniente è stata applicata anche la seguente formula che considera la β diversità in maniera additiva ([32] - eqn. 2):
È stata inoltre calcolata la varianza del numero di specie per ogni strato. Successivamente, sono stati applicati i due indici considerati più efficaci tra le misure di β diversità per dati di presenza-assenza (indice di similarità di Jaccard, Cj; indice di similarità di Sørensen, Cs - eqn. 3 e eqn. 4):
dove a = numero di specie presenti nei due strati a confronto; b = numero di specie presenti solo nel primo strato; c = numero di specie presenti solo nel secondo strato.
La matrice floristica (“specie x rilievi”), inoltre, è stata sottoposta a classificazione e ordinamento mediante il programma Syntax2000 ([30]) allo scopo di indagare il grado di somiglianza fra i rilievi e fra gli strati, nonché per contribuire a chiarire in che proporzione le differenze floristiche fra gli strati siano dovute all’età del ceduo.
Alle specie rinvenute sono stati anche associati i rispettivi valori degli indici di Ellenberg (secondo [29]), allo scopo di confrontare la diversa risposta del corteggio floristico ai caratteri medi stazionali e ottenere una misura della “distanza ecologica” fra i rilievi e fra gli strati. I valori degli indici (luce, temperatura, continentalità del clima, umidità del suolo, pH del suolo, nutrienti) costituiscono la valutazione numerica del segnale che ciascuna specie fornisce sull’incidenza dei principali fattori ecologici nel determinare le caratteristiche del sito ([28]). La tabella “specie x indici” è stata elaborata a partire dalla tabella “specie x rilievi”, calcolando per prima cosa la distanza tra tutte le possibili coppie di specie rinvenute negli strati per mezzo della distanza multivariata descritta in Podani & Schmera ([31]) (eqn. 5):
dove Sijk è la distanza tra le specie j e k per la variabile i; xij e xik sono i valori assunti delle specie j e k per la variabile i e al nominatore è presente il range tra il valore massimo e minimo assunto dalla variabile i.
Poi sulla base di questa nuova tabella, è stata calcolata la dissimilarità funzionale di Ellenberg (D) tra gli strati presi a coppie (A-B; A-C; B-C) con la modalità descritta in Bacaro et al. ([4]) (eqn. 6):
dove wiB è la minima distanza di Ellenberg tra le specie i dello strato A e tutte le specie dello strato B; wjA è la minima distanza di Ellenberg tra le specie j dello strato B e tutte le specie dello strato A e nA e nB sono rispettivamente tutte le specie rinvenute negli stratiA e B.
Infine, la significatività della diversità intra-strato e tra gli strati è stata validata (con 999 permutazioni) tramite il programma Permanova v.1.6 (Permutational analysis of variance - [1]) che tramite test F permette di ottenere la probabilità P che la quantificazione della diversità ottenuta sia dovuta al solo caso.
Risultati
Caratteri dendrometrici e strutturali
I parametri dendrometrici rilevati nelle aree di saggio (Tab. 2) evidenziano come la struttura dei soprassuoli sia piuttosto variabile anche all’interno del singolo strato, in particolare per quanto riguarda la densità di ceppaie, polloni e matricine. Senza dimenticare che si tratta di una caratteristica tipica dei sistemi naturali anche se sensibilmente antropizzati, tale situazione è riconducibile comunque alla mancanza di trattamenti uniformi, continui e pianificati che ha caratterizzato per lungo tempo la gestione selvicolturale dei boschi esaminati. Un disordine colturale cui sta cercando di rimediare, almeno per quanto riguarda la tenuta di Vicarello (strati A e B), il recente piano di assestamento forestale ([15]).
Tab. 2 - Principali parametri dendrometrici nei tre strati cronologici.
| Parametro | Strato metà turno (A) | Strato fine turno (B) | Strato oltre turno (C) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | A2 | A3 | A4 | A5 | B1 | B2 | B3 | B4 | B5 | C1 | C2 | C3 | C4 | C5 | |
| Numero di matricine (n ha-1) |
96 | 32 | 96 | 64 | 64 | 64 | 32 | 0 | 32 | 0 | 0 | 0 | 0 | 32 | 32 |
| Numero di polloni con diametro < 5 cm (n ha-1) | 96 | 191 | 32 | 478 | 0 | 3662 | 1943 | 2452 | 2325 | 2038 | 0 | 0 | 0 | 0 | 0 |
| Numero di polloni con diametro > 5 cm (n ha-1) | 4140 | 3885 | 2261 | 4204 | 1752 | 796 | 764 | 2866 | 2452 | 350 | 1592 | 446 | 1083 | 1306 | 1178 |
| Numero di polloni vivi (n ha-1) |
4236 | 4076 | 2293 | 4682 | 1752 | 4459 | 2707 | 5318 | 4777 | 2389 | 1592 | 446 | 1083 | 1306 | 1178 |
| Numero di polloni morti (n ha-1) | 3153 | 3758 | 2102 | 3089 | 1210 | 0 | 32 | 541 | 1242 | 0 | 1815 | 223 | 796 | 1019 | 541 |
| Totale ceppaie (n ha-1) | 478 | 223 | 446 | 287 | 159 | 433 | 382 | 287 | 255 | 32 | 446 | 191 | 287 | 414 | 255 |
| Numero di polloni per ceppaia | 15 | 35 | 10 | 27 | 19 | 11 | 7 | 20 | 24 | 75 | 8 | 4 | 7 | 6 | 7 |
| Diametro medio (cm) | 9.1 | 8.0 | 12.6 | 8.3 | 14 | 15 | 14.7 | 8.5 | 9.08 | 11.3 | 17.5 | 22.2 | 17 | 18.5 | 17.8 |
| Area basimetrica fusti vivi (m2 ha-1) | 27 | 19.6 | 28.4 | 21.8 | 26.8 | 14 | 12.9 | 16.3 | 15.9 | 3.5 | 38.1 | 17.3 | 24.6 | 35.2 | 29.2 |
| Area basimetrica fusti morti (m2 ha-1) | 5.3 | 3.5 | 3.2 | 3.5 | 1.9 | 0 | 0 | 1.1 | 0.3 | 0 | 7.6 | 0.6 | 3.2 | 4.6 | 3.1 |
| Incremento medio area basimetrica (m2 ha-1 anno-1) | 2.26 | 1.63 | 2.84 | 1.82 | 2.68 | 0.82 | 0.76 | 0.96 | 0.93 | 0.19 | 2.24 | 1.02 | 1.45 | 2.07 | 1.72 |
Sulle variabili dendrometriche, naturalmente, influiscono anche gli ultimi trattamenti effettuati, insieme ai naturali dinamismi evolutivi del ceduo coetaneo. Il numero di polloni, a partire dalla classe 10, è molto elevato nello strato A rispetto a B, dove è stato effettuato il diradamento, e a C dove il processo di selezione naturale ha operato più a lungo. La presenza di polloni di dimensioni molto piccole ha significato solo nello strato B ed è la conseguenza dell’intenso riscoppio di nuovi getti dalle ceppaie diradate. Con il taglio intercalare sono stati asportati in media il 40% dei polloni, quasi esclusivamente sul piano dominato, pari al 20% dell’area basimetrica.
Il confronto con gli studi alsometrici relativi a cedui castanili localizzati su altri rilievi dell’antiappennino vulcanico laziale ([26], [21]) ha evidenziato la maggiore fertilità media degli strati A e C rispetto a B e la generale ridotta densità dei popolamenti studiati rispetto ai modelli di normalità proposti.
Va inoltre sottolineato che un numero maggiore numero di specie partecipa allo strato arboreo dello strato C (circa il 16% della componente specifica totale): in particolare, sono abbondanti carpino bianco (Carpinus betulus) e nocciolo (Corylus avellana), accompagnati in misura minore da acero campestre (Acer campestre) e sambuco (Sambucus nigra).
Dati floristici
Durante i rilievi floristici sono state rinvenute complessivamente 142 entità ripartite in 63 famiglie di piante vascolari, 129 delle quali all’interno delle aree circolari campionate; di queste: 82 nello strato A, 83 nello strato B, 74 nello strato C. Alcune di esse sono di particolare interesse floristico, in quanto nel Lazio sono rare e/o poco comuni ([2], [3]) oppure protette: si tratta di Iberis umbellata (rinvenuta nello strato B), Galanthus nivalis (strato B), Hypericum androsaemum (strato C), Lilium bulbiferum subsp. croceum (al di fuori dei punti campionati, nei pressi dello strato A) e Narcissus pöeticus (strato A) . Il ritrovamento di una entità del genere Rubus nello strato C, riferibile con buona probabilità a Rubus scaber, se confermata, si potrebbe rivelare di particolare interesse in quanto questa specie era stata indicata per l’Italia, secondo Pignatti ([27]), “solamente una volta nel Teramano e molto probabilmente per errore”. Approfondimenti sono in corso presso l’Erbario della Tuscia.
Spettri biologico e corologico
Lo spettro biologico normale (Tab. 3) non mostra grandi differenze rispetto a quello ponderato per frequenze (Tab. 4): in entrambi i casi si nota una maggiore percentuale delle fanerofite nello strato C e un aumento delle nanofanerofite all’aumentare dell’età del soprassuolo arboreo. Interessante è l’andamento delle emicriptofite: si ha un valore simile per gli strati A e C e un valore più elevato per lo strato B sia nello spettro normale che in quello ponderato per frequenze. Le terofite sono presenti con una percentuale maggiore nello strato A, per diminuire nello strato B e aumentare nuovamente, ma in misura minore, nel terzo strato come illustrato da entrambi gli spettri. Andamento simile è mostrato dalle geofite.
Tab. 3 - Spettro biologico normale (valori %).
| Tipo | Strato A | Strato B | Strato C |
|---|---|---|---|
| Fanerofite | 25.61 | 25.30 | 33.78 |
| Nanofanerofite | 4.88 | 6.02 | 6.76 |
| Camefite | 4.88 | 4.82 | 4.05 |
| Emicriptofite | 29.27 | 42.17 | 28.38 |
| Terofite | 9.76 | 6.02 | 8.11 |
| Geofite | 25.61 | 15.66 | 18.92 |
Tab. 4 - Spettro biologico ponderato per frequenze (valori %).
| Tipo | Strato A | Strato B | Strato C |
|---|---|---|---|
| Fanerofite | 36.16 | 30.50 | 40.33 |
| Nanofanerofite | 2.82 | 4.00 | 6.63 |
| Camefite | 3.95 | 7.00 | 3.87 |
| Emicriptofite | 20.34 | 37.50 | 22.10 |
| Terofite | 8.47 | 4.50 | 6.08 |
| Geofite | 28.25 | 16.50 | 20.99 |
Gli spettri corologici (Tab. 5 e Tab. 6) mostrano una netta predominanza di specie eurasiatiche.
Tab. 5 - Spettro corologico normale (valori %).
| Tipo | Strato A | Strato B | Strato C |
|---|---|---|---|
| Endemiche | 1.22 | 2.41 | 1.35 |
| Stenomediterranee | 8.54 | 8.43 | 8.11 |
| Eurimediterranee | 15.85 | 13.25 | 16.22 |
| Medit.-Mont. | 0.00 | 1.20 | 0.00 |
| Eurasiatiche | 58.54 | 53.01 | 59.46 |
| Atlantiche | 1.22 | 3.61 | 1.35 |
| Orof. S-Europ. | 1.22 | 1.20 | 0.00 |
| Boreali | 8.54 | 12.05 | 5.41 |
| Gruppi ad ampia distribuzione |
4.88 | 4.82 | 8.11 |
Tab. 6 - Spettro corologico ponderato per frequenze (valori %).
| Tipo | Strato A | Strato B | Strato C |
|---|---|---|---|
| Endemiche | 0.56 | 4.00 | 0.55 |
| Stenomediterranee | 7.34 | 9.50 | 8.84 |
| Eurimediterranee | 16.95 | 14.50 | 16.57 |
| Medit.-Mont. | 0.00 | 0.50 | 0.00 |
| Eurasiatiche | 59.32 | 53.50 | 61.33 |
| Atlantiche | 1.69 | 3.00 | 1.66 |
| Orof. S-Europ. | 1.69 | 0.50 | 0.00 |
| Boreali | 8.47 | 9.50 | 2.76 |
| Gruppi ad ampia distribuzione |
3.95 | 5.00 | 8.29 |
Le stenomediterranee e le eurimediterranee sono presenti in tutti e tre gli strati senza grandi variazioni, le mediterranee - montane sono presenti in bassissime percentuali solo nello strato B. Le specie atlantiche sono scarsamente rappresentate; le orofite sono completamente assenti nello strato C. Le specie boreali invece sono presenti con percentuali maggiori nei due strati A e B, si riducono fortemente nello strato C. La ripartizione delle specie ad ampia distribuzione varia: costante nei due strati A e B e con aumento quasi doppio nello strato C.
Matrice floristica
La matrice floristica “specie x rilievi” consta di 132 righe x 23 colonne e indica tutte le entità censite, la loro presenza nei punti di sondaggio e nei vari strati. La matrice, ordinata in base alle frequenze delle specie nei vari punti di campionamento, mostra come siano 32 le specie presenti in tutti e 3 gli strati, tra queste quelle più frequenti sono: Castanea sativa, Hedera helix e Acer pseudoplatanus; 9 invece sono le specie mancanti nello strato A (presenti dunque in B e C soltanto), 20 quelle mancanti nello strato B (presenti in A e C), 17 quelle mancanti nello strato C (presenti in A e B). All’estremo opposto della matrice vi sono numerose specie sporadiche esclusive dei singoli strati o di singoli punti: 13 sono le specie esclusive dello strato A (tra cui il già citato Narcissus p ö eticus), 25 quelle dello strato B (ad es., Iberis umbellata, Galanthus nivalis), 13 quelle dello strato C (Hypericum androesamum, Prunus avium, Rubus scaber). La riorganizzazione in chiave fitosociologica della matrice come descritto precedentemente ha permesso di elaborare gli spettri delle affinità fitosociologiche, sia normali che ponderati per frequenze (Tab. 7 e Tab. 8), che hanno mostrato alcune differenze.
Tab. 7 - Spettro delle affinità floristiche normale (valori %).
| Tipo | Strato A | Strato B | Strato C |
|---|---|---|---|
| Specie di Fagetalia sylvaticae | 23.17 | 21.69 | 27.03 |
| Specie di Quercetalia pubescenti-petraeae | 13.41 | 24.1 | 10.81 |
| Specie di Querco-Fagetea | 34.15 | 28.92 | 29.73 |
| Specie di Quercetea ilicis | 4.88 | 3.61 | 2.7 |
| Specie di prati e incolti | 8.54 | 12.05 | 1.35 |
| Specie di margine nitrofilo | 14.63 | 6.02 | 24.32 |
| Altre specie | 1.22 | 3.61 | 4.05 |
Tab. 8 - Spettro delle affinità floristiche ponderato per frequenze (valori %).
| Tipo | Strato A | Strato B | Strato C |
|---|---|---|---|
| Specie di Fagetalia sylvaticae | 25.42 | 24.50 | 24.31 |
| Specie di Quercetalia pubescenti-petraeae | 15.25 | 23.50 | 11.60 |
| Specie di Querco-Fagetea | 38.42 | 32.00 | 33.70 |
| Specie di Quercetea ilicis | 5.08 | 3.00 | 3.31 |
| Specie di prati e incolti | 4.52 | 7.50 | 0.55 |
| Specie di margine nitrofilo | 10.73 | 4.50 | 23.76 |
| Altre specie | 0.56 | 5.00 | 2.76 |
Entrambi gli spettri evidenziano la maggiore percentuale di specie appartenenti ai Quercetalia pubescenti - petraeae nello strato B e la scarsità di queste specie nello strato C. L’andamento delle specie dei Querco - Fagetea mostra in entrambi gli spettri una diminuzione della percentuale dallo strato A allo strato B, per poi crescere nuovamente nello strato C. Le specie dei Quercetea ilicis decrescono con l’aumentare dell’età degli strati in termini di numero di specie (spettro normale); anche lo spettro ponderato mostra una maggiore frequenza di queste specie nello strato A. Le specie di prati e incolti sono più frequenti nello strato B, e pressoché assenti nello strato C. Per quel che riguarda le specie riconducibili alle cenosi di margine nitrofilo è netto il loro maggior peso nello strato C, sia in termini di diversità (spettro normale) che di frequenza nei punti di sondaggio (spettro ponderato).
Indici di diversità
I valori di α, β, γ diversità e di varianza di ciascuno strato vengono riportati nella Tab. 9. Lo strato B presenta il maggiore numero di specie complessivo (γ-diversità) e la più alta varianza di numero di specie. La α-diversità non risulta significativamente diversa tra gli strati considerati. Anche la β diversità, calcolata con le due modalità già descritte nella sezione Materiali e metodi, non mostra differenze tra i tre strati.
Tab. 9 - Valori di α, β, γ diversità e varianza per ciascuno strato.
| Indice | Strato A | Strato B | Strato C |
|---|---|---|---|
| Numero totale di specie (γ diversità) |
82 | 83 | 74 |
| Media numero di specie (α) | 38 | 39.8 | 36.4 |
| Varianza numero di specie | 28.5 | 99.2 | 8.3 |
| β = γ / α | 2.16 | 2.09 | 2.03 |
| β = γ - α | 44.0 | 43.2 | 37.6 |
Peraltro, confrontando gli strati a coppie (Tab. 10), l’indice di Whittaker assume il valore più elevato per la coppia di strati B-C, il valore più basso per la coppia A-C. Questo risultato è confermato anche dagli indici di similarità di Jaccard e Sørensen: si ha infatti una maggiore similarità (e quindi una bassa β diversità) tra gli strati A e C, una minore similarità (corrispondente a una più elevata β diversità) tra gli strati B e C.
Tab. 10 - Valori degli indici di Whittaker, di Jaccard e Sørensen per ogni coppia di strati.
| Indice | Strato A | Strato B | Strato C | |
|---|---|---|---|---|
| Indice di Whittaker |
Strato A | - | 0.41 | 0.32 |
| Strato B | 0.41 | - | 0.48 | |
| Strato C | 0.32 | 0.48 | - | |
| Indice di Jaccard |
Strato A | - | 0.42 | 0.50 |
| Strato B | 0.42 | - | 0.35 | |
| Strato C | 0.50 | 0.35 | - | |
| Indice di Sørensen |
Strato A | - | 0.59 | 0.67 |
| Strato B | 0.59 | - | 0.52 | |
| Strato C | 0.67 | 0.52 | - | |
Classificazione e ordinamento dei rilievi
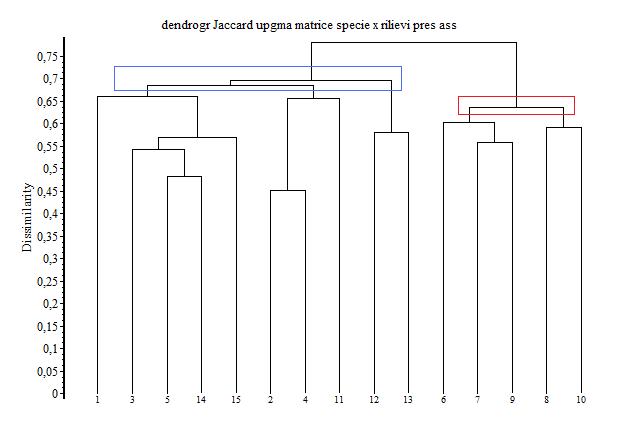
I risultati degli indici di diversità e similarità sono confermati dalla classificazione e dall’ordinamento dei rilievi in base alla presenza-assenza delle specie. Il dendrogramma (Fig. 5) ottenuto dall’algoritmo di Jaccard mostra due grandi gruppi di rilievi, uno più omogeneo che comprende tutti i rilievi dello strato B e l’altro comprendente i rilievi degli strati A e C, indicando una maggiore affinità tra questi.
Fig. 5 - Dendrogramma ottenuto dalla classificazione dei rilievi in base alla presenza-assenza delle specie.
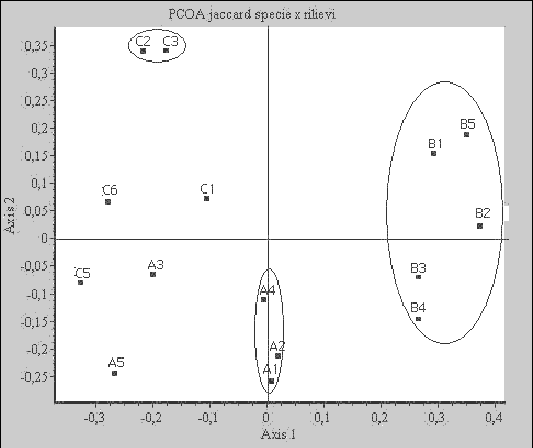
Il diagramma ottenuto a partire dall’ordinamento dei rilievi (Fig. 6) mostra risultati in linea a quelli del dendrogramma, con i rilievi appartenenti allo strato B nettamente discostati da quelli degli strati A e C.
Fig. 6 - Diagramma ottenuto dall’ordinamento dei rilievi mediante Principal Coordinates Analysis (PCOA) con indice di similarità di Jaccard
Indici di Ellenberg
I valori degli indici di Ellenberg sono stati associati alle specie ritrovate nei singoli punti di sondaggio. A quelle coppie specie-fattore ecologico che nella matrice di dati di Pignatti et al. ([29]) presentano valore 0 (comportamento non precisato) è stato attribuito un valore basato sulla conoscenza personale degli Autori del comportamento di tali specie nel contesto geografico dell’area indagata, allo scopo di rendere la matrice elaborabile dai programmi di analisi multivariata. Nella Tab. 11 si riportano i valori medi assunti dai sei fattori nei punti di sondaggio ottenuti come somma di tutti i valori assunti per un determinato indice di Ellenberg dalle singole specie - matrice “specie x indici” - diviso il numero delle specie presenti nel punto di sondaggio).
Tab. 11 - Valori medi assunti dai sei fattori di Ellenberg (L= luce; T= temperatura; C= continentalità del clima; U= umidità del suolo; R= reazione del terreno o pH; N= nutrienti presenti nel suolo) nei punti di sondaggio.
| Fattore | A1 | A2 | A3 | A4 | A5 | B1 | B2 | B3 | B4 | B5 | C1 | C2 | C3 | C4 | C5 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | 5.1 | 5.4 | 5.1 | 5.3 | 5.3 | 5.4 | 5.6 | 5.5 | 5.3 | 6.0 | 5.4 | 5.2 | 5.0 | 5.4 | 5.2 |
| T | 6.3 | 6.3 | 6.3 | 6.2 | 6.1 | 6.5 | 6.3 | 6.3 | 6.3 | 6.5 | 6.2 | 6.4 | 6.1 | 6.3 | 6.3 |
| C | 4.6 | 5.0 | 4.6 | 4.7 | 4.6 | 4.6 | 4.7 | 4.9 | 4.7 | 4.7 | 4.5 | 4.6 | 4.6 | 4.7 | 4.8 |
| U | 4.6 | 4.7 | 4.8 | 4.8 | 4.9 | 4.6 | 4.6 | 4.4 | 4.7 | 4.7 | 4.7 | 4.8 | 5.0 | 4.8 | 4.9 |
| R | 6.2 | 6.3 | 6.0 | 6.4 | 6.3 | 6.0 | 6.1 | 6.2 | 6.0 | 6.1 | 5.8 | 5.9 | 5.5 | 6.3 | 6.0 |
| N | 5.2 | 5.0 | 5.7 | 5.2 | 5.9 | 4.9 | 4.8 | 4.8 | 5.0 | 4.9 | 5.8 | 6.2 | 5.6 | 6.0 | 5.8 |
A partire dagli indici di Ellenberg, allo scopo di meglio visualizzare le informazioni ed effettuare confronti tra le comunità rilevate e le associazioni vegetali note in letteratura, sono stati realizzati gli ecogrammi, che costituiscono una sorta di “impronta digitale” di una comunità e permettono l’individuazione immediata delle sue caratteristiche ecologiche.
I 15 ecogrammi ottenuti sono stati raggruppati per strato allo scopo di confrontare la diversa risposta del corteggio floristico ai caratteri medi stazionali e confrontati con quelli realizzati da Pignatti et al. ([28]) per 33 associazioni presenti in Italia, in modo da individuare quelle con temperamento più affine ai boschi studiati. Tale procedura ha valore solo preliminare, in quanto non avendo la nostra ricerca previsto rilievi condotti secondo il metodo fitosociologico (quindi su aree omogenee), gli ecogrammi realizzati a partire da una tabella basata su aree di saggio randomizzate (che includono una discreta eterogeneità alla scala di indagine propria della fitosociologia) non sono del tutto comparabili con quelli proposti da Pignatti et al. ([28]), che si basano su tabelle di associazioni. Comunque, secondo il procedimento adottato, le comunità studiate mostrano ecogrammi affini a quelli delle associazioni Aquifolio-Fagetum Gentile 1969 ed Ornithogalo pyrenaici-Carpinetum Marincek et al. 1982, che presentano però una certa componente di continentalità, e Physospermo-Quercetum petraeae Oberd. et Hofmann 1967 ed Echinopo siculi-Quercetum frainetto Blasi et Paura 1993 che mostrano invece valori maggiori di luce e temperatura, e indicano che i tre strati di cedui castanili indagati possono inserirsi in un contesto ecologico intermedio tra quello delle faggete termofile di bassa quota e quello dei querceti mesofili.
Validazione statistica
Le matrici presenza-assenza delle specie e di dissimilarità tra plot ottenuta dagli indici di Ellenberg sono state validate dal punto di vista statistico con il programma Permanova v.1.6 che ha mostrato, per entrambe le matrici, come i tre strati siano significativamente distinti tra loro (Tab. 12 e Tab. 13).
Tab. 12 - Risultati ottenuti dalla validazione statistica Permanova in base alla presenza-assenza delle specie.
| Layer | t | P_perm | P_MC |
|---|---|---|---|
| (A,B) | 1.8166 | 0.0030 | 0.0120 |
| (A,C) | 1.3866 | 0.0210 | 0.0690 |
| (B,C) | 2.1023 | 0.0090 | 0.0030 |
Tab. 13 - Risultati ottenuti dalla validazione statistica con Permanova in base agli indici di Ellenberg.
| Layer | t | P_perm | P_MC |
|---|---|---|---|
| (A,B) | 2.3104 | 0.0140 | 0.0040 |
| (A,C) | 1.6432 | 0.0050 | 0.0320 |
| (B,C) | 2.9108 | 0.0090 | 0.0010 |
Nel primo caso (presenza-assenza delle specie), la coppia di strati maggiormente distinti tra loro (cioè con il più basso valore P di probabilità che il risultato sia dovuto al solo caso) risulta A-B (metà turno - fine turno). La coppia di strati che presenta il valore di P più alto è A-C, confermando i risultati precedenti. Gli strati B e C sono anch’essi significativamente distinti ma con un valore di P intermedio alle altre due coppie.
Per quanto la matrice di dissimilarità tra plot si notano alcune differenze rispetto ai risultati avuti in base alla sola presenza-assenza delle specie. In questo caso infatti gli strati maggiormente distinti tra loro sono A e C.
Discussione
Il maggior numero di specie censite negli strati A (ceduo a metà turno ancora non diradato) e B (ceduo a fine turno diradato) rispetto allo strato C (ceduo oltre turno non diradato) conferma quanto osservato da vari Autori: il numero di specie nel sottobosco tende a diminuire con l’età a meno che non intervenga un disturbo (ad esempio, un diradamento) a indurre un nuovo aumento della quantità di luce sotto la copertura arborea ([5], [16], [33]). Anche dagli spettri biologici si può evidenziare l’impatto sulla componente erbacea del diradamento avvenuto nello strato B: le geofite risentono maggiormente delle modifiche al microambiente conseguenti all’intervento selvicolturale, come il maggiore inaridimento relativo del suolo causato dalla maggiore quantità di radiazione che penetra la copertura, al contrario delle specie arboree caratterizzate da una maggiore inerzia nell’occupazione e nell’abbandono di un sito ([33]). L’elevata presenza di emicriptofite e un aumento delle nanofanerofite passando dallo strato A allo strato B è un ulteriore conseguenza del diradamento, che favorisce l’ingresso delle specie arbustive. La bassa percentuale di geofite nello strato C, rimasto indisturbato dall’ultima ceduazione, è probabilmente correlata al maggior disturbo arrecato dalla fauna selvatica (presente in questo strato in misura maggiore in quanto il territorio limitrofo è più ricco di animali selvatici), che nutrendosi di bulbi e rizomi limita la partecipazione delle geofite allo spettro biologico di questo strato. Le terofite, presenti nello strato C con una percentuale simile a quella dello strato A (che risente ancora degli effetti della ceduazione), indicano un altro fattore di disturbo rappresentato dal pubblico che frequenta il castagneto, soprattutto alla ricerca di funghi, provenendo dalla limitrofa e facilmente accessibile faggeta di Monte Raschio, e che diventa vettore di specie caratterizzanti gli ambienti aperti e di margine.
Gli spettri corologici confermano questo tipo di disturbo nello strato C dove le specie ad ampia distribuzione si rinvengono con percentuali superiori rispetto agli altri due strati, indicando la presenza di una flora più banalizzata.
Gli spettri delle affinità floristiche mostrano aspetti interessanti e differenti per i tre strati. La presenza di specie dei Fagetalia sylvaticae in maggiore misura nello strato C, oltre che con le caratteristiche stazionali (soprattutto giacitura e pendenza), è spiegabile con la menzionata presenza in prossimità del ceduo castanile della faggeta di Monte Raschio, aspetto che spiega anche la scarsità di specie dei Quercetea ilicis e dei Quercetalia pubescenti-petraeae; a esempio, alcune specie quali Sanicula europaea, Hypericum androsaemum e Prunus avium sono esclusive di questo strato.
L’ordine dei Quercetalia pubescenti-petraeae è rappresentato in maggior misura nello strato B: alcune specie tipiche dei querceti sono infatti presenti solo in questa parte dell’area di studio come Buglossoides purpurocaerulea, Oryzopsis miliacea, Clinopodium vulgare, Scutellaria columnae. La presenza di specie di prati e incolti, presenti in minima parte nello strato C e maggiormente diffuse negli altri due strati A e B, è spiegabile con il fatto che questi castagneti presentano un minore grado di chiusura, e con la più estesa presenza di pascoli e seminativi nelle aree confinanti.
Lo spettro delle affinità floristiche conferma il disturbo di origine antropica a cui è soggetto lo strato C, con un’elevata presenza e frequenza di specie appartenenti ad ambienti di margine nitrofilo (Sambucus nigra, S. ebulus, Urtica dioica, Veronica hederifolia, Chelidonium majus, ecc.) che peraltro indicano anche una fertilità del suolo relativamente maggiore. Alcune di queste specie mancano nello strato B, indicando la presenza di suoli meno ricchi di nutrienti e relativamente più aridi, anche per la maggiore acclività del terreno.
La matrice floristica complessiva dà indicazioni sul dinamismo in atto che segue il taglio intercalare nello strato B, ed è testimoniata dalla presenza di specie pioniere come Fraxinus ornus. Queste specie sono assenti nello strato C dove si assiste a una situazione di “chiusura”, evidenziata sugli ecogrammi dai bassi valori assunti per il fattore luce e dai valori più alti riguardanti il fattore umidità del suolo, entrambi correlati alla mancanza di interventi selvicolturali da diversi anni.
Gli strati A e C sono quelli caratterizzati da suoli relativamente più fertili sebbene differiscano per acclività e sono anche quelli che da maggiore tempo non sono stati oggetto di trattamenti selvicolturali, il che spiega anche le affinità tra questi due strati sia nella classificazione dei rilievi che nell’ordinamento. Queste somiglianze sono riscontrabili anche dai risultati ottenuti dal calcolo degli indici di Whittaker, Jaccard e Sørensen.
Gli ecogrammi confermano le differenze tra i tre strati: i maggiori valori di luce e temperatura negli indici di Ellenberg ottenuti per lo strato B sono una conseguenza del diradamento, come riportato anche da Decocq ([13]), che include la gestione selvicolturale tra le cause che possono influenzare i valori assunti dagli indici di Ellenberg per il fattore luce. L’intervento selvicolturale ha apportato una maggiore quantità di radiazione solare al suolo che ha ridotto anche l’umidità del terreno, come indicano i valori più bassi registrati per questo fattore nello strato in considerazione. Il valore dei nutrienti del suolo è più basso, indicando una minore fertilità, aspetto che motiva la scarsa presenza in questo strato di specie esigenti di nutrienti come Narcissus p ö eticus e altre specie dei margini nitrofili già menzionate. Si fa notare che questa specie è assente nello strato C sebbene il terreno sia fertile, in quanto necessita anche di luce, elemento ridotto dalla maggiore copertura arborea in questo strato.
L’effetto combinato del diradamento e delle caratteristiche stazionali, in particolare la maggiore acclività, ha portato a un inaridimento relativo del suolo che ha influenzato la componente erbacea differenziando lo strato B dagli altri due.
Non sono state riscontrate relazioni statisticamente significative tra i parametri floristici analizzati nei singoli punti di campionamento e le variabili dendrometriche, in particolare quelle più direttamente espressive della copertura superiore del soprassuolo. Ciò può essere ricondotto all’alta variabilità dell’area basimetrica e della densità di individui all’interno dei singoli strati ma fa anche pensare che lo sviluppo della flora nel sottobosco non dipenda solo dalla quantità di luce che vi arriva e che anche altre condizioni microstazionali, quali la disponibilità di acqua e nutrienti e i processi competitivi, giochino un ruolo importante, come ipotizzato da vari autori ([24], [16]).
Conclusioni
Sulla base dei risultati ottenuti è possibile esprimere una valutazione complessivamente positiva sulle modalità di gestione forestale attualmente applicate. Ciò a motivo dell’elevato numero di specie censite ma soprattutto della presenza di specie rare o che mantengono il legame con il corteggio floristico della vegetazione cosiddetta potenziale, sebbene i cedui castanili possano subire un impatto non trascurabile dovuto agli interventi selvicolturali.
Sono possibili alcune considerazioni sul ruolo del diradamento di media-alta intensità realizzato nella seconda metà del ciclo e che appare come l’evento colturale in grado di determinare due situazioni differenti a fine turno.
Gli strati A e C, anche in virtù della loro somiglianza floristica, rappresentano il momento intermedio e quello finale di un ciclo produttivo che non prevede tagli intercalari. Questo tipo di regime colturale è tutt’altro che eccezionale nei territori dell’antiappennino vulcanico laziale in quanto si verifica spesso nelle numerose piccole proprietà private dove la ridotta superficie non rende economico il diradamento. In questo caso, la flora tende ad accentuare nel tempo, rispetto al ceduo diradato, i caratteri più nemorali che già manifesta a metà del turno. Tuttavia, sulla composizione floristica del ceduo influiscono anche fattori esogeni come il disturbo antropico derivante dalla fruizione turistica, la predazione della fauna selvatica e il tipo di boschi naturali che circonda il ceduo castanile. Nell’area studiata il mosaico forestale si presenta assai variegato poiché le peculiari condizioni microclimatiche interne alla caldera lacustre e il gioco delle esposizioni e delle pendenze mettono a contatto boschi con fisionomia molto diversa quali leccete, cerrete, formazioni miste di specie mesofile e faggete termofile. Un trattamento con turno di 18-20 anni in assenza di diradamenti può ritenersi un modulo colturale particolarmente valido all’interno di un’area protetta in quanto consente una maggiore evoluzione della flora in senso nemorale, nonostante il disturbo turistico. Inoltre, la periodica ceduazione consente il mantenimento del bosco di castagno, che, altrimenti, verrebbe gradualmente sostituito dalla faggeta.
Il confronto tra i risultati relativi agli strati A e B evidenziano le differenze che si vengono a determinare nell’ambiente interno al popolamento e, di conseguenza, sulla compagine floristica, attraverso un diradamento di medio-forte intensità eseguito nella seconda metà del ciclo colturale, in genere 4-5 anni prima del taglio finale.
Il diradamento è un evento di disturbo non trascurabile con molteplici effetti intercorrelati e non sempre facilmente quantificabili ([25]) che interrompe alcuni processi ecologici e ne stimola altri. A esempio, favorisce il repentino aumento dell’irraggiamento del suolo creando condizioni idonee all’affermazione di più specie vegetali che, però, in parte risultano banali e/o dal limitato valore naturalistico. Contemporaneamente, il diradamento accentua le diversità microstazionali in quanto la sua reale intensità può variare anche molto a livello puntuale e ciò si riflette sulle condizioni di illuminazione determinate dal gioco di luci dirette e diffuse. Nello strato B l’intervento colturale ha accentuato gli effetti della minore fertilità e maggiore aridità relativa del suolo rispetto agli altri strati. Il confronto non tiene conto del fatto che gli effetti di un diradamento di simile intensità nello strato A potrebbero essere diversi in termini di velocità di richiusura delle chiome a motivo della maggiore fertilità.
Anche un modulo colturale che preveda l’esecuzione di un diradamento nella seconda metà del ciclo colturale può essere compatibile con gli scopi di conservazione della biodiversità perseguiti dai Parchi, visto che l’intervento mantiene elevata la ricchezza floristica in termini di numero di specie. Tuttavia, nelle stazioni a minore fertilità sarebbe consigliabile l’allungamento del turno da 18 a 22-25 anni e l’esecuzione di più (2-3) diradamenti di bassa intensità. Una riduzione più graduale della densità arborea consentirebbe una migliore protezione del suolo e di limitare l’impatto negativo dell’intervento sulla naturalità del corteggio floristico. Allo stesso tempo, consentirebbe di ottenere un maggiore diametro medio dei polloni a fine turno compensando in parte i maggiori oneri di gestione. Inoltre, recenti indagini hanno dimostrato che diradamenti precoci e regolari nel tempo possono anche ridurre l’incidenza del fenomeno della cipollatura che può compromettere il reddito del taglio finale dei cedui castanili ([6]).
Non sono state trovate relazioni statisticamente significative tra i parametri floristici analizzati e le variabili dendrometriche. L’analisi floristica è risultata comunque un utile strumento di indagine per differenziare fra loro boschi cedui di uguale fisionomia ma differente età e regime selvicolturale.
In conclusione, sebbene i risultati presentati abbiano carattere preliminare, essendo basati su un arco temporale ridotto che non consente di rappresentare in maniera completamente esaustiva i fenomeni indagati, è stato comunque possibile verificare alcuni effetti dell’attuale gestione selvicolturale sulla componente floristica, e porre le basi per ulteriori approfondimenti che prevedano un maggior numero di punti di sondaggio e di strati cronologici in modo da poter giungere a una completa tipologia dei castagneti del Lazio.
Inoltre, non esistendo in Italia il castagneto come forma di vegetazione potenziale, per ricercare un modello neutrale con cui confrontare la composizione floristica dei diversi cedui occorrerà estendere lo studio ai boschi che costituiscono la tappa matura della serie cui i vari castagneti appartengono (generalmente, faggete termofile e cerrete) e ai castagneti da frutto.
Ringraziamenti
Si desidera ringraziare per la preziosa collaborazione Carlo Ricotta (Dipartimento di Biologia Vegetale dell’Università di Roma “La Sapienza”) e l’Azienda Agricola Vicarello S.p.A. di Bracciano.
References
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar