Cultural treatments and woody debris: the study case of beech forests in Casentino (Italy)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 6, Pages 39-49 (2009)
doi: https://doi.org/10.3832/efor0551-006
Published: Jan 29, 2009 - Copyright © 2009 SISEF
Research Articles
Abstract
This paper shows the first results about quantity and quality of dead wood on the ground in beech forests of Foreste Casentinesi, Monte Falterona and Campigna National Park. The presence of dead wood depends on several factors such as forest productivity, natural disturbances and human activities. Data from some mature managed beech stands have been collected and compared with those from unmanaged beech forest (Sasso Fratino full-protected Reserve). Results do not show significant differences between managed and unmanaged forests. The dead wood varies between 5 and 8 Mg ha-1 (in terms of total dry weight) mostly composed by fine woody debris.
Keywords
Faggio, Necromassa legnosa, Gestione forestale, Parco Nazionale, Foreste Casentinesi
Introduzione
La necromassa legnosa in foresta, costituendo un habitat ideale per la vita di numerosi vertebrati ed invertebrati ([5], [42], [44], [48], [49], [51], [55], [60], [61], [26], [40]), e fungendo spesso da nicchia ideale per la germinazione di molte specie arboree ([64], [37]), è considerata come componente fondamentale per il mantenimento e l’incremento della biodiversità ([21], [67], [16], [36], [58], [68], [31], [47]). La quantità di legno morto in foresta è riconosciuta, in base al “criterio 4” del MCPFE (Maintenance, Conservation and Appropriate Enhancement of Biological Diversity in Forest Ecosystems, [35]), come uno degli indicatori di gestione forestale sostenibile ([30], [34]).
La necromassa legnosa grossolana (Coarse Woody Debris - CWD, [45], [57], [23]) e la lettiera fogliare svolgono un’importante azione di accumulo e volano in relazione ai flussi energetici ed al ciclo dei nutrienti ([24], [39], [41]), costituendo il più cospicuo serbatoio di carbonio per i processi di migrazione della sostanza organica nel suolo, anche se ad oggi sono ancora poco note le quantità massime “utili” a tale scopo ([20], [11], [3], [1]). Inoltre il legno morto presente al suolo protegge il terreno dall’erosione limitando l’azione battente dell’acqua, trattiene l’umidità ed offre un’efficace protezione dal congelamento ([33]). La quantità di legno morto in bosco è considerata nel Protocollo di Kyoto come uno dei compartimenti per il calcolo del carbonio assorbito.
Relativamente ai rapporti tra quantità di necromassa e interventi colturali, numerosi autori ([18], [32], [27], [15], [2]) evidenziano come questi ultimi possano incidere negativamente, e che di conseguenza in boschi poco disturbati sia più elevata la quantità di legno morto.
Nel presente lavoro si descrive, dal punto di vista quantitativo e qualitativo, la necromassa legnosa al suolo in aree campione situate in boschi di faggio di ambiente appenninico, cercando una relazione tra i quantitativi di legno morto e gli interventi selvicolturali pregressi. La ricerca è stata condotta nelle faggete del Parco Nazionale delle Foreste Casentinesi, Monte Falterona e Campigna (AR, FC, FI), dove sono stati confrontati soprassuoli avviati all’alto fusto e ordinariamente gestiti con tagli intercalari, con altri sottoposti a regime di tutela integrale.
Materiali e metodi
L’ambiente
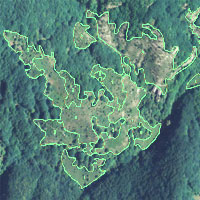
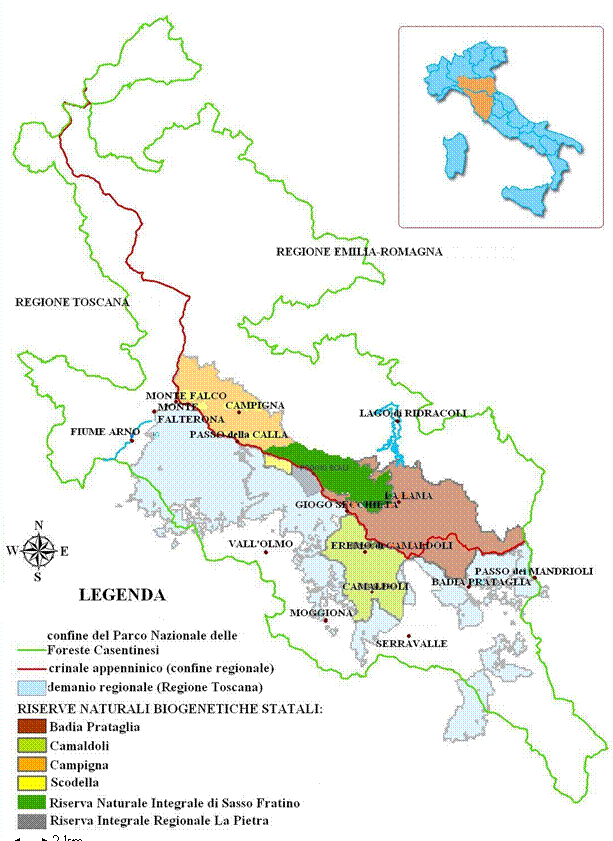
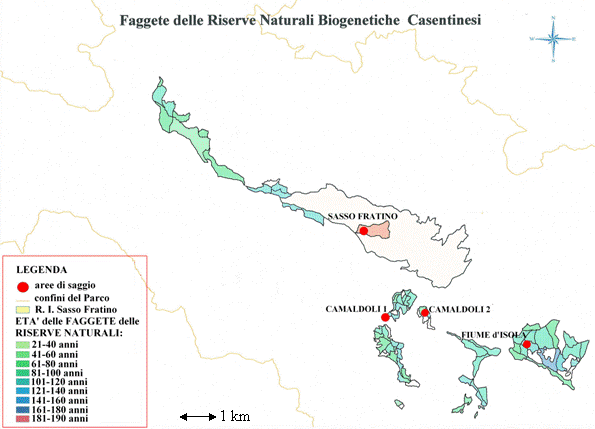
L’area di studio comprende le faggete ubicate all’interno delle Riserve Naturali Biogenetiche del Parco Nazionale delle Foreste Casentinesi, Monte Falterona e Campigna (Fig. 1) per una superficie complessiva di 668 ettari.
Fig. 1 - Le Riserve Naturali Biogenetiche del Parco Nazionale delle Foreste Casentinesi, Monte Falterona e Campigna (AR, FI, FC).
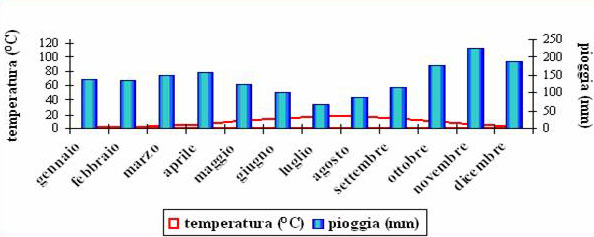
Il versante toscano del Parco è caratterizzato da un substrato pedogenetico prevalente di Arenaria Macigno dell’Oleocene da cui derivano suoli bruno-acidi, con humus di tipo Mull; nel versante romagnolo il substrato pedogenetico è Marnoso Arenaceo e i suoli sono in prevalenza bruno-marmorizzati, con humus di tipo Moder ([54]). Il clima è tipicamente montano ad impronta mediterranea, caratterizzato da maggior piovosità in autunno e primavera, e dall’assenza di periodi di siccità estiva (Fig. 2).
Fig. 2 - Diagramma umbro-termico relativo alla stazione di Camaldoli (AR - 1111 m s.l.m.).
La vegetazione è ascrivibile alle faggete eutrofiche a dentarie della classificazione di Bernetti & Mondino ([4]).
Per quanto riguarda gli aspetti normativi relativi al legno morto è da ricordare che nel Piano del Parco ([43]) è prevista la salvaguardia degli alberi morti in bosco, ed il rilascio di un adeguato quantitativo di necromassa in piedi o a terra (10 piante ad ettaro); nei confinanti territori toscani e romagnoli invece le leggi regionali prevedono l’asportazione dalle tagliate dei residui di lavorazione e l’allontanamento dei materiali di scarto in quanto facilmente infiammabili o di ostacolo per una corretta regimazione delle acque.
Aree di studio
Attraverso una ricerca d’archivio (Corpo Forestale dello Stato, Ufficio Territoriale per la Biodiversità, Pratovecchio - AR) e l’implementazione delle informazioni attraverso l’uso di GIS, sono state selezionate una serie di particelle (aree di indagine) ritenute rappresentative delle condizioni medie dei soprassuoli di altofusto di faggio situati all’interno del Parco Nazionale; fra queste ne sono state individuate tre per le quali si è potuto disporre di una più lunga e dettagliata cronologia degli interventi colturali (denominate “Camaldoli 1”, “Camaldoli 2”, “Fiume d’Isola”). Una quarta particella è stata selezionata nella Riserva Naturale Integrale di Sasso Fratino (denominata “Sasso Fratino”), in un tratto fisionomicamente affine alle precedenti, considerata come area di confronto in quanto da lungo tempo non soggetta ad interventi colturali (Fig. 3).
Fig. 3 - Localizzazione delle aree di saggio all’interno delle Riserve Naturali Biogenetiche.
Rilievi in campo e analisi di laboratorio
In fase di impostazione della ricerca, sono state considerate alcune tra le metodologie proposte per la determinazione del CWD e più in generale della necromassa in foresta ([17], [10], [65], [66], [62]), metodologie nelle quali la precisione di stima è in parte legata alle caratteristiche (di forma, dimensione e stato di alterazione) del materiale da campionare.
Nel presente lavoro, avendo come obiettivo la valutazione di tutto il materiale legnoso morto presente al suolo (ramaglia fine compresa), e dovendo poi procedere alla comparazione tra quantità di necromassa con caratteristiche potenzialmente diverse, è parso opportuno utilizzare una metodologia che fornisse un dato quantitativo poco influenzato dal tipo di materiale da campionare.
Per tali motivi, data anche la difficoltà di misura della necromassa di piccole e medie dimensioni in termini di volume ([53]) si è optato per la determinazione del peso secco del materiale legnoso.
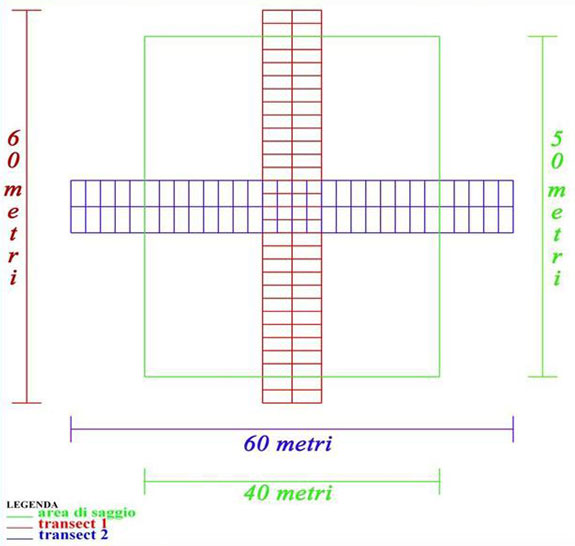
In ciascuna area d’indagine è stata realizzata un’area di saggio (AdS) di 2000 m2 (50x40 m), in cui sono stati eseguiti il cavallettamento totale e la stima dell’altezza su un terzo degli individui censiti. Per le analisi riguardanti la necromassa al suolo, in ciascuna AdS sono stati realizzati due transetti lineari (6x60 m) passanti per il centro dell’area, ortogonali tra loro (orientamento Est-Ovest e Nord-Sud) e suddivisi in 60 sezioni di 6 m2 (3x2 m) (Fig. 4). Su un campione di 20 sezioni, estratte casualmente in ogni coppia di transetti, è stato determinato il peso fresco (Pf) della necromassa legnosa a terra con l’utilizzo di una stadera. Facendo riferimento alla letteratura inerente l’argomento ([59], [46], [56], [64], [50], [36], [63]), il materiale pesato è stato suddiviso in funzione delle dimensioni (D) espresse dal diametro a metà lunghezza (ؽl) e del grado di decomposizione (A, B, C) come riportato in Tab. 1. Per ogni categoria è stata prelevata e conservata in sacchetti di polietilene sigillati una serie di campioni provenienti dalle quattro AdS. Presso i laboratori del DISTAF dell’Università di Firenze ne è stato determinato il peso secco (Ps) ottenuto utilizzando una stufa ventilata (103 °C) in cui il materiale legnoso è rimasto fino al raggiungimento di un peso costante ([52]).
Fig. 4 - Schema rappresentativo dei transetti realizzati in ogni AdS per l’esecuzione dei rilievi della necromassa al suolo.
Tab. 1 - Distinzione della necromassa in nove categorie tipologiche; con “ؽl” è indicato il diametro a metà lunghezza (* = Modificato, da [59], [46], [56], [64], [50], [36], [63]).
| Categorie * | |||
|---|---|---|---|
| Grado di decomposizione | Classe dimensionale |
||
| A | legno con corteccia e tessitura delle fibre compatta | D1 | ؽl < 5 cm |
| B | legno senza corteccia e tessitura delle fibre compatta | D2 | 5 cm < ؽl < 10 cm |
| C | legno senza corteccia, tessitura delle fibre molle o polverosa | D3 | ؽl > 10 cm |
Elaborazione dati
Per ogni area sono stati rilevati i principali parametri dendrometrici ed è stata ricostruita la cronologia e l’entità degli interventi colturali.
La stima del Ps della necromassa rilevata da ciascuna sezione è stata ottenuta applicando il coefficiente medio Ps/Pf rilevato sul campione in laboratorio e distinto per aree d’indagine e per categoria; i valori ottenuti sono stati espressi in Mg/ha.
I confronti fra i quantitativi medi di necromassa sono stati eseguiti attraverso l’analisi della varianza ponendo come fonti di variazione le aree d’indagine; in caso di valori di F almeno significativi (p<0.05), è stato eseguito il test di Duncan al fine di verificare eventuali gruppi omogenei.
Risultati
Le principali caratteristiche di ogni area sono riassunte in Tab. 2 ed in Tab. 3, sia per quanto riguarda i dati dendrometrici che quelli relativi alla storia delle utilizzazioni. Nelle AdS realizzate in soprassuoli ordinariamente gestiti, fustaie transitorie di faggio di circa cento anni di età, il numero medio di piante ad ettaro è 360 (Dev. St. 9), l’area basimetrica media della massa in piedi è 51 m2 ha-1 (Dev. St. 3) e il volume è mediamente pari a 680 m3 ha-1 (Dev. St. 21). Complessivamente le utilizzazioni a cui sono stati assoggetti i soprassuoli variano da 71 a 88 m3 ha-1. Nella AdS localizzata a Sasso Fratino il bosco è una fustaia di età superiore ai cento anni, con una massa in piedi di oltre 900 m3 ha-1 ed area basimetrica di più di 60 m2 ha-1; l’ultima utilizzazione censita è stata eseguita negli anni ’30 ha asportato poco più di 30 m3 ha-1 di massa in piedi.
Tab. 2 - Dati dendrometrici relativi a ciascuna AdS; con “G” è indicata l’area basimetrica, con “V” il volume, con “dg” il diametro medio di area basimetrica, con “Hdg” l’altezza della pianta di diametro medio, con “Hd” l’altezza dominante. (*): è stata utilizzata la tavola di cubatura a doppia entrata di Bernetti (1977) per il faggio di Vallombrosa (FI).
| AdS | Camaldoli 1 | Camaldoli 2 | Fiume d’Isola | Sasso Fratino |
|---|---|---|---|---|
| Quota (m s.l.m.) | 1256 | 1306 | 1267 | 1394 |
| Esposizione | S-SO | E | S-O | N-NE |
| Pendenza (gradi) | 21 | 23 | 17 | 40 |
| Età (anni) | 100 | 100 | 100 | > 100 |
| G (m2/ha) | 51 | 54 | 48 | 60 |
| V (m3/ha) * | 693 | 694 | 657 | 942 |
| dg (cm) | 43 | 36 | 41 | 55 |
| Hdg (m) | 23.1 | 22.3 | 26.2 | 29 |
| Hd (m) | 25 | 24.3 | 27.9 | 29.1 |
Tab. 3 - Dati relativi alle utilizzazioni eseguite nelle particelle in cui sono ubicate le AdS.
| AdS | Periodo di utilizzazione |
Massa asportata totale (m3/ha) |
Anno ultima utilizzazione |
Entità ultima utilizzazione (m3/ha) |
|---|---|---|---|---|
| Camaldoli 1 | 1961-1987 | 88.5 | 1987 | 16.8 |
| Camaldoli 2 | 1951 | 85.5 | 1951 | 85.8 |
| Fiume d’Isola | 1979-1989 | 71.4 | 1989 | 27.6 |
| Sasso Fratino | 1936 | 33 | 1936 | 33 |
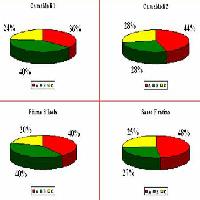
Dall’analisi della varianza emergono differenze molto significative fra le aree rispetto al quantitativo medio di necromassa presente al suolo (F[3, 75] = 3.74; p< 0.01). Dal test di Duncan si osserva che il valore medio più basso (5.03 Mg ha-1) si registra in “Fiume d’Isola”, mentre quello più alto (8.78 Mg ha-1) in “Camaldoli 1”. I valori riguardanti l’area di “Sasso Fratino” e di “Camaldoli 2” si collocano in posizione intermedia (rispettivamente 5.98 Mg ha-1 e 5.51 Mg ha-1), e non differiscono in modo significativo dai due precedenti (Fig. 5, Tab. 4).
Fig. 5 - Valori medi (± errore standard) di peso secco della necromassa in ogni AdS (limiti fiduciari p<0.001).
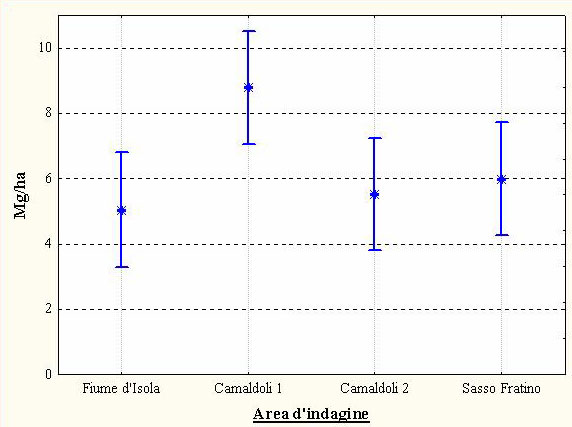
Tab. 4 - Dati di necromassa relativi ad ogni AdS; con il termine “CV” è indicato il coefficiente di variabilità; i gruppi omogenei fanno riferimento al test di Duncan.
| AdS | Necromassa (Mg/ha) |
CV | Gruppi omogenei |
|---|---|---|---|
| Fiume d’Isola | 5035 | 52% | a |
| Camaldoli 2 | 5511 | 60% | ab |
| Sasso Fratino | 5983 | 38% | ab |
| Camaldoli 1 | 8785 | 69% | b |
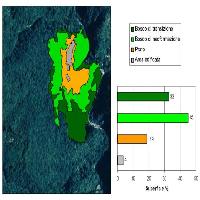
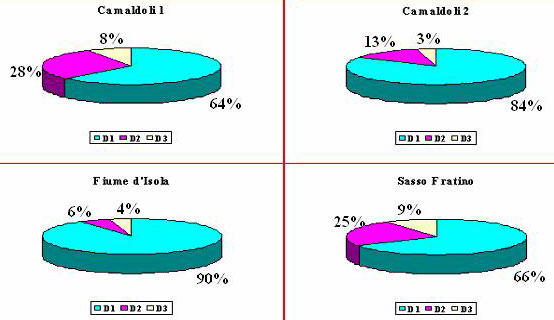
Osservando la ripartizione del legno morto in classi dimensionali (Fig. 6) si nota che il materiale di dimensioni minori (D1) è presente in maggiori quantità in tutte le aree, al contrario quello di dimensioni maggiori (D3) è il meno rappresentato.
Fig. 6 - Ripartizione della necromassa secondo il grado di decomposizione; con “A” è indicato lo stadio meno decomposto, con “C” quello più alterato e con “B” lo stadio intermedio.
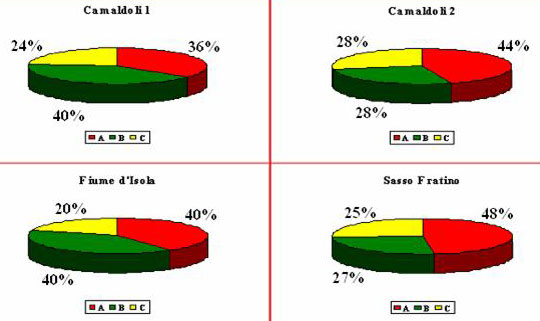
Per quanto riguarda la suddivisione del materiale secondo lo stadio di degradazione (Fig. 7) è stato rilevato che in tutte le aree il legno col maggior grado di decomposizione (“C”) è quello meno presente: in “Sasso Fratino” e “Camaldoli 2” lo stato di alterazione maggiormente presente è “A”, mentre in “Camaldoli 1” è il “B” ed in “Fiume d’Isola” “A” e “B” hanno lo stesso valore.
Fig. 7 - Ripartizione della necromassa in classi dimensionali; con “D1” è indicato il materiale di minori dimensioni, con “D3” quello di maggiori dimensioni e con “D2” quello di dimensioni intermedie.
Discussione
Le aree indagate risultano sostanzialmente omogenee per quanto riguarda il quantitativo di necromassa legnosa presente al suolo: nonostante sia stata osservata una discreta variabilità all’interno delle AdS, solo in “Camaldoli 1” la quantità rilevata è leggermente superiore. Questa differenza è probabilmente imputabile all’entità dell’ultima utilizzazione avvenuta nella particella nel 1987. Visto il modesto quantitativo di legname esboscato (16.8 m3 ha-1) è probabile che siano state lasciate in piedi piante che in seguito sono morte per la concorrenza o che buona parte del materiale utilizzato sia stato rilasciato sul posto influenzando i valori ad oggi rilevati.
Per quanto riguarda la distribuzione del legno morto in classi dimensionali è stato rilevato che in tutte le aree il materiale di piccole dimensioni è quello presente in maggiori quantità, probabilmente perché apportato al terreno con maggiore costanza, provenendo prevalentemente dalla rottura dei rami. Questo fenomeno è già stato segnalato da altri Autori ([21], [9], [25]) i quali hanno rilevato che in soprassuoli adulti (come quelli del presente lavoro) il maggior contributo all’accumulo di necromassa legnosa al suolo, sotto forma di rami o di tronchi atterrati, avviene a seguito di eventi meteorici. Al contrario nei boschi giovani la concorrenza fra le piante è la causa più frequente di mortalità, e la necromassa presente in questi soprassuoli è costituita principalmente da alberi morti in piedi.
A proposito della ripartizione della necromassa secondo il grado di decomposizione si osserva che il legno maggiormente degradato (“C”) è, in tutte le AdS, quello presente in minore quantità: infatti il materiale in avanzato stato di decomposizione è difficilmente distinguibile dal resto della sostanza organica al suolo a cui spesso è intimamente legato. Da studi condotti in faggete abetine della Polonia ([29], [67]) emerge che il legno morto a contatto col terreno si decompone in tempi relativamente “veloci” (circa 15 anni per il faggio) rispetto a quello in piedi poichè gli organismi del suolo accelerano i processi di degradazione. Inoltre l’andamento climatico influisce fortemente su tali processi: in zone caldo-umide la velocità di decomposizione del materiale legnoso è maggiore rispetto a quanto avviene luoghi con clima secco e fresco ([67]).
Secondo alcuni studi ([21], [2]) l’intervento dell’uomo induce cambiamenti sulle dinamiche di “produzione” di legno morto: da un input di necromassa relativamente continuo, tipico di condizioni naturali, ad un input periodico dovuto alla cadenza delle utilizzazioni. Su larga scala anche le variazioni di densità della popolazione umana ed il conseguente cambiamento dell’intensità dei tagli influenzano le dinamiche di accumulo di legno morto in bosco ([25]). Inoltre, considerato che le caratteristiche del materiale legnoso al suolo sono condizionate dalle dinamiche che hanno portato alla morte della pianta o parte di essa ([16], [8], [36]) e che il quantitativo e la tipologia di CWD cambiano a seconda degli stadi successionali della foresta, è logico affermare che col crescere dell’età del bosco la quantità di legno aumenti così come anche le dimensioni medie del materiale accumulato ([21], [67]). Diversi autori ([67], [8], [13], [53]) affermano che la quantità di CWD è minore in boschi sottoposti a tagli colturali rispetto a quella presente in soprassuoli lasciati alla naturale evoluzione. Di Filippo et al. ([12]) sostengono che l’abbondanza di necromassa come conseguenza dell’assenza delle utilizzazioni forestali è uno degli elementi più rappresentativi della vetustà di una foresta.
Nell’ambito del nostro studio è stato invece rilevato che nell’area di Sasso Fratino, nonostante l’assenza dell’attività colturale per un periodo di tempo prolungato (oltre 70 anni), la quantità di legno morto al suolo è paragonabile a quella di soprassuoli soggetti ad ordinaria gestione: ciò è probabilmente da collegare sia alla lenta dinamica di accumulo della necromassa sia alla velocità dei processi di degradazione della stessa. Quanto osservato denota quindi che la fase evolutiva che sta attraversando Sasso Fratino è quella di una foresta matura non ancora in fase senescente: l’apporto di necromassa al terreno dovuto alla caduta di alberi è attualmente estremamente modesto in buona parte della Riserva e del tutto assente nelle AdS campionate. È tuttavia possibile che, in tempi più lunghi, qui si riscontreranno maggiori quantitativi di legno morto al suolo dovuti all’aumento della mortalità di alberi vecchi e di grandi dimensioni, fenomeno caratterizzante boschi nella fase di invecchiamento più marcata (old-growth - [21], [25], [56], [67], [28], [12], [16], [34], [37], [31]).
Secondo quanto riferito da vari Autori ([22], [7], [14], [38]), il quantitativo di sostanza secca del soprassuolo arboreo (radici comprese) rilevato in faggete nord europee (Danimarca, Germania e Svezia) affini per età e stato colturale a quelle oggetto di studio oscilla tra i 300 ed i 200 Mg ha-1. Calamini et al. ([6]) e Gregori & Miclaus ([19]) hanno rilevato per fustaie transitorie di faggio dell’Appennino pistoiese valori di sostanza secca della biomassa epigea di 319 Mg ha-1. Gli stessi Autori rilevano una produzione media annua di lettiera di 2 Mg ha-1 (l’85% della quale costituito da foglie). In rapporto a questi dati dunque, la necromassa da noi misurata risulterebbe essere circa il 2.5% della biomassa totale, più del doppio delle lettiera annualmente prodotta.
Conclusioni
L’analisi comparata dei dati ha evidenziato differenze statisticamente significative tra le aree indagate rispetto alla quantità di legno morto presente al suolo: i risultati ottenuti hanno tuttavia messo in luce una sostanziale omogeneità tra i valori riscontrati in soprassuoli soggetti ad ordinaria gestione e quelli della Riserva Integrale, fenomeno osservato anche nelle foreste di conifere svedesi da Siitonen et al. ([56]).
Nonostante il campione fosse di non grandi dimensioni e limitato ad una sola specie, sembra lecito affermare, nell’ambito dei casi esaminati, l’assenza di relazione tra quantità di legno morto al suolo ed interventi di utilizzazione. Di conseguenza risulta evidente come, almeno nel “breve” periodo, le utilizzazioni forestali realizzate secondo i metodi tradizionali non abbiano inciso negativamente sul quantitativo di necromassa presente sul terreno.
Ringraziamenti
Si ringraziano per la preziosa collaborazione Alessandro Bottacci, Giovanni Quilghini ed Antonio Zoccola dell’UTB (CFS) di Pratovecchio (AR).
References
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar