Silvicultural options in ageing holm oak (Quercus ilex L.) coppices in Gargano: results after 14 growing seasons
Forest@ - Journal of Silviculture and Forest Ecology, Volume 5, Pages 318-336 (2008)
doi: https://doi.org/10.3832/efor0552-0050318
Published: Dec 12, 2008 - Copyright © 2008 SISEF
Research Articles
Abstract
In the frame of a long-term research program concerning management of ageing holm oak coppices, results available after 14 growing seasons are reported in present paper. Experimental treatments include: A) 50 standards per hectare, all of the same age; B) 250 standards per hectare, all of the same age; C) 140 standards per hectare, with two different ages; D) conversion to high forest; E) natural evolution (control). A total of 15 permanent plots were established (5 treatments x 3 replicates per treatment) and the experimental design used is that of randomised blocks. Based on observations concerning seedlings and shoots development and standards growth and competitive effects, the following preliminary results may be highlighted: i) recovering the traditional coppicing system with few standards per hectare represents a valid option from both ecological and shoots growth point of view; the stools, with few standards per hectare, showed a larger number of sprouts, provided with a higher average height and larger diameters; ii) uneven-aged standards represent a good alternative form the points of view of both landscape impact immediately after felling operations and stand resistance to climatic damages; iii) a good alternative is to apply conversion treatments to high forest, whenever their site quality allows these operations.
Keywords
Holm oak, Coppice, Conversion to high forest, Regeneration, Standard density
Introduzione e finalità dello studio
Il leccio rappresenta una delle specie stenomediterranee il cui areale interessa con continuità le coste italiane, quelle della Dalmazia, della Grecia, della Turchia occidentale e del nord Africa, indipendentemente dal substrato pedologico, per raggiungere la massima diffusione nella Penisola iberica. Gli antichi annoverarono il leccio tra le piante sacre, probabilmente per l’importanza che aveva nell’economia dell’epoca, per il temperamento frugale, e per la non comune vigoria e longevità, sia quando è governato a ceduo che a fustaia. L’Italia, come riferisce Pavari ([33]), conserva ancora oggi numerosissimi toponimi conferiti proprio da questa specie.
Oltre all’ottimo combustibile, ancora oggi apprezzato sul mercato, la specie forniva legname per attrezzi e parti di manufatti soggetti a forte usura, corteccia per le industrie conciarie, ghianda e frasca per l’allevamento del bestiame e per la fauna selvatica.
I boschi di questa specie hanno svolto e svolgono ancora oggi un ruolo insostituibile nella protezione del terreno dall’eccessiva insolazione e dal dilavamento da parte delle acque meteoriche, anche in condizioni geomorfologiche estremamente difficili.
Nonostante l’azione dell’uomo, in generale tesa a sostituire il leccio nelle migliori condizioni pedologiche e geomorfologiche, per la sua diffusione, per la non comune resilienza nei riguardi di fattori di disturbo e per la capacità di colonizzare i luoghi più impervi, a giusta ragione questa specie è da considerare tra le specie che caratterizzano il paesaggio dei Paesi del Mediterraneo.
Come riferisce Pignatti ([34]), la lecceta può rappresentare in un’ampia area del Mediterraneo la vegetazione naturale potenziale. La lecceta rappresenta inoltre uno dei rari esempi d’equilibrio nell’interazione tra fattori naturali ed attività antropiche.
La presenza del leccio è rilevata soprattutto laddove le condizioni stazionali risultano non favorevoli alle latifoglie più esigenti; i suoli calcarei e i versanti meridionali, con scarsa ritenzione idrica, sono spesso conquistati dalle leccete ([20]).
La specie in esame presenta un accrescimento policiclico, il getto primaverile è predeterminato; a questo si aggiungono possibili altri getti prolettici che si manifestano in concomitanza di eventi piovosi. Da uno studio di Giovannini et al. ([21]) risulta che in annate caratterizzate da primavere siccitose seguite da stagioni favorevoli, la somma degli accrescimenti prolettici supera la lunghezza del getto primaverile.
Sul promontorio del Gargano le formazioni di leccio, ascrivibili secondo uno studio di Falinski & Pedrotti ([17]) all’associazione Ostryo-Quercetum ilicis, tendono a colonizzare, a basse quote, le aree ai limiti delle zone sottoposte ad agricoltura. Inoltre tali formazioni tendono a spingersi nelle zone interne su esposizioni soleggiate raggiungendo quote superiori a 800 m s.l.m. Si ritiene che la superficie dei boschi di leccio sia in fase di espansione, tenendo conto dell’evoluzione che in generale ha interessato queste formazioni boschive negli ultimi 40-50 anni ([24]).
Come riportato in La Marca et al. ([27]), sul Gargano i cedui di leccio costituiscono accorpamenti, anche di notevole estensione, che in genere coincidono con terreni fortemente degradati. In alcune zone particolarmente impervie il ceduo di leccio rappresenta la formazione vegetale dominante. Sulle pendici esposte a sud è possibile ritrovare il leccio, in forma sporadica o a piccoli gruppi, fino alle quote più elevate del Gargano (circa 1000 m s.l.m.). È certo comunque che da epoche storiche il leccio costituisce una componente non secondaria del paesaggio garganico.
Fino a 40-50 anni fa la forma di governo più diffusa per le formazioni a prevalenza di leccio era il ceduo con rilascio di poche matricine per ettaro. In occasione dei tagli di maturità (il turno minimo stabilito dalle Prescrizioni di Massima e di Polizia forestale è pari a 15 anni) eseguiti con una cadenza molto ravvicinata, sia per soddisfare la forte domanda di legna da ardere che per ottenere il carbone “cannello”, il più apprezzato sul mercato, non era rara la pratica della dicioccatura effettuata per aumentare la quantità di legna da asportare ad ogni fine ciclo ([27]).
È probabile che gli attuali cedui, puri o a netta prevalenza di leccio, siano la diretta conseguenza di un eccessivo sfruttamento da parte dell’uomo ([27]).
Con il diminuire della pressione antropica, grazie alla notevole capacità di ripresa che è caratteristica di questa specie, la situazione generale dei cedui di leccio è sensibilmente migliorata dal punto di vista della copertura del suolo, degli accumuli provvigionali e, nelle situazioni edafiche migliori, anche dal punto di vista della biodiversità.
Le tendenze colturali oggi in atto nei cedui possono ricondursi sostanzialmente al mantenimento del sistema ceduo, alla conversione in fustaia o alla conversione da ceduo matricinato a ceduo composto ([1], [7], [8], [11]).
Vari lavori negli ultimi anni hanno affrontato diverse problematiche inerenti la gestione dei cedui di leccio. In merito alla letteratura più recente, si citano: [23], [21], [27], [2], [3], [4], [6], [10], [16], [24], [9].
Esperienze interessanti sul leccio, in corso in Francia ([5], [38], [12], [13], [14], [15], [39]) e in Spagna ([32], [22], [36], [35], [37], [19]), sono in grado di fornire indicazioni di carattere programmatico di validità generale.
È innegabile, comunque, che sia in atto una progressiva ripresa delle utilizzazioni per legna da ardere, legata per lo più alle condizioni di benessere (barbecue, pizzerie, caminetti, ecc.) che a ragioni di sussistenza.
Il presente lavoro prende in esame una sperimentazione avviata in una lecceta del Gargano nel 1994. I problemi che ci si propose di affrontare furono quelli di una messa a punto di indirizzi selvicolturali appropriati alla nuova realtà economica e ambientale in cui vengono a trovarsi i cedui di leccio, in particolare all’interno del Parco Nazionale del Gargano. Fu anche tenuto presente che una variante da tenere in particolare considerazione era rappresentata dal notevole degrado del suolo, dalla sua origine calcarea, dall’esposizione particolarmente calda dei versanti, dalle precipitazioni talvolta scarsissime (fino a 450-500 mm) e dalla loro sfavorevole distribuzione nel corso dell’anno. La ricerca avviata ha inteso dare riposte su basi sperimentali alle scelte selvicolturali possibili in presenza di cedui invecchiati di leccio a densità pressoché colma. Su questi stessi cedui sono stati affrontati anche tematiche riguardanti l’epoca di taglio ([31]) e lo studio della biomassa ([30]).
Con il presente lavoro si sono poste a confronto le diverse forme di governo e trattamento a partire da cedui invecchiati di leccio a 14 anni dall’inizio della sperimentazione.
Allo stato attuale, tenuto conto della notevole estensione di queste formazioni forestali e della loro importanza dal punto di vista della tutela del suolo, per ragioni estetiche e paesaggistiche, i maggiori problemi da affrontare in sede di pianificazione forestale riguardano la complessa problematica legata alla selvicoltura da applicare nell’ambito delle aree protette nel rispetto sia degli interessi generali (biodiversità, sostenibilità, naturalità) che degli interessi che più direttamente riguardano le tradizioni e le aspettative delle popolazioni locali (paesaggio, cultura, economia locale, attività tradizionali - [24]).
Materiali e Metodi
Geologia e pedologia
L’area oggetto del presente lavoro ricade su una valle con andamento NE-SO, delimitata a nord dal Monte Spigno (1008 m. s.l.m.), formatosi in seguito all’instaurarsi di faglie trascorrenti.
In genere i suoli di quest’area, formatisi sotto copertura forestale, sono profondi e di media fertilità. La loro colorazione rossa è dovuta all’elevato contenuto di idrossidi di ferro e di acidi umici saturi di calcio che non si decompongono velocemente donando agli orizzonti superficiali una colorazione che va dal bruno scuro al rosso bruno al bruno chiaro ([29]). Il rapporto C/N presenta valori elevati a causa della scarsa dotazione di azoto totale e della ridotta attività biologica, dovuta alla degradazione conseguente al disboscamento e all’agricoltura di rapina. Secondo la classificazione Word Reference Base della FAO ([18]) questi suoli rientrano nella categoria dei Chromic Cambisols.
Clima
Il clima del Gargano è influenzato dalla particolare morfologia del territorio. Infatti è un promontorio bagnato su tre lati, proteso verso il mare, disconnesso dall’Appennino dalla Fossa Bradanica. Raggiunge quote massime intorno ai 1000 metri. Nei riguardi del clima in particolare sono determinanti i venti principali con direzione N-O/S-E e N-S: bora e scirocco. I venti che arrivano da S-SE sono solitamente secchi e portano ad elevare la temperatura: lo scirocco infatti giunge sull’Italia ricco di umidità che rilascia come pioggia, per poi interessare il Gargano quando ne è privo. Nel piano basale e nei versanti della valle Carbonara e di Monte Spigno si risentono gli effetti di questi venti, come secchezza e temperature elevate e si ritrovano formazioni di leccio e in generale latifoglie xerotolleranti.
I dati termo-pluviometrici sono riferiti alla stazione Monte S. Angelo, 843 m. s.l.m., la quale dista pochi chilometri dalla località“Inversa di Spigno” dove sono ubicati i boschi oggetto del presente lavoro. Il periodo preso in esame è il quarantennio 1965-2005.
La temperatura media annua è pari a 12.1 °C e la media annua delle precipitazioni è di 774 mm di pioggia. Le piogge sono concentrate in inverno, tra novembre e gennaio, con un picco in marzo. Il periodo di ariditàè compreso tra metà luglio e fine agosto. Dal punto di vista fitoclimatico l’area è ascrivibile alla sottozona fredda del Lauretum di Pavari e, alle quote superiori, alla zona del Castanetum caldo. L’indice di aridità di De Martonne, pari a 74, evidenzia condizioni favorevoli per la vegetazione forestale.
Cenni sulle aree sperimentali
La località studiata si trova a circa 650 m sul livello del mare con esposizione sud, e pendenza di circa il 40%. Le aree sperimentali si trovano in prossimità dei pascoli e dei seminativi situati nella vallata sottostante (Fig. 1). Il terreno si presenta accidentato con rocciosità di tipo calcareo affiorante e pietrosità diffusa (30-40%).
Fig. 1 - Veduta dell’area che ospita la sperimentazione a circa 10 anni dagli interventi.

Le aree sperimentali, realizzate nel 1994 in un ceduo invecchiato di circa 45 anni, sono ubicate in località Inversa di Spigno, nel comune di Monte Sant’Angelo (FG), la zona risulta interamente compresa nel perimetro del Parco Nazionale del Gargano.
La composizione specifica è dominata dal leccio con partecipazione sporadica di carpino nero e orniello. Il sottobosco è composto soprattutto da Asplenium onopteris, Asplenium trichomanes, Cyclamen neapolitanum, Crataegus monogyna e Ruscus aculeatus.
All’interno della lecceta viene praticato, soprattutto nel periodo estivo, il pascolo caprino. È stata inoltre osservata la presenza del cinghiale.
Dal punto di vista fitosociologico la lecceta in questione è attribuibile all’Ostryo-Quercetum ilicis ([17]).
Piano sperimentale
La sperimentazione mette a confronto diverse forme di trattamento e governo, partendo da cedui invecchiati di leccio e utilizza un disegno sperimentale a blocchi randomizzati.
Le tesi prese in esame, replicate tre volte, sono:
- Tesi A: trattamento a ceduo matricinato con 50 matricine ha-1;
- Tesi B: trattamento a ceduo matricinato con 250 matricine ha-1;
- Tesi C: trattamento a ceduo composto con 140 matricine ha-1 di cui 80 di un turno e 60 di 2 turni;
- Tesi D: conversione ad alto fusto;
- Tesi E: evoluzione naturale.
Nel 1994 sono state pertanto individuate 15 aree di saggio permanenti di forma quadrata con superficie di 1600 m2 ognuna e sono state rilasciate delle fasce di rispetto, larghe 10 m, tra un’area di saggio e l’altra per evitare l’effetto margine.
Per tutte le aree all’inizio della sperimentazione (prima del taglio) è stato effettuato un cavallettamento totale con successiva individuazione delle piante da abbattere.
Su un campione delle piante abbattute (100 piante) è stata realizzata una curva ipsometrica ed una tavola di cubatura a una entrata per la determinazione dei volumi per diametri fino a 3 cm a petto d’uomo ([27], [28], [26]).
Le aree riferite alle tesi A, B e C sono state recintate con filo spinato al fine di preservare lo spazio in questione dal pascolamento.
Nelle aree rappresentanti le tesi Ceduo è stata effettuata una planimetria riportante la posizione di tutte le matricine. In ogni area è stato individuato e riportato in planimetria un transect diagonale orientato NE-SO di 20 m di lunghezza e 2 di larghezza, all’interno del quale sono stati censiti e cartografati tutti i semenzali e tutte le ceppaie.
I rilievi sul ceduo, iniziati l’anno successivo al taglio, hanno riguardato l’insediamento della rinnovazione da seme, il ricaccio dei polloni e la selezione naturale che interessa la componente arborea (sia polloni che matricine). In tutte le aree sperimentali è stato effettuato un monitoraggio delle condizioni (crescita, vitalità e danni da agenti meteorici) della componente gamica, agamica, delle matricine e degli allievi che dovranno costituire la futura fustaia transitoria nelle aree a conversione.
Rilievi effettuati
I rilievi effettuati nel 2007 hanno riguardato la componente agamica e gamica del ceduo, a 14 anni dall’inizio della sperimentazione.
Nelle aree sottoposte a conversione ad alto fusto, e in quelle lasciate ad evoluzione naturale è stato eseguito il cavallettamento di tutte le piante presenti nelle aree di saggio.
I rilievi sulla rinnovazione agamica hanno interessato tutte le ceppaie comprese nell’area dei transect di ogni area di saggio. L’indagine ha riguardato la determinazione del numero di polloni per ceppaia, il diametro a m 1.30 di tutti i polloni e la misura dell’altezza totale di 15 polloni per ogni ceppaia.
Per quanto riguarda la rinnovazione gamica, è stato effettuato il censimento di tutti i semenzali presenti sulla superficie dei transect, con misurazione del diametro al colletto, dell’altezza e la loro georeferenziazione.
Nelle tesi A, B e C, inoltre, sono state individuate 27 matricine, tre per ogni area, attorno alle quali sono state tracciate aree di saggio circolari aventi raggio pari a tre volte il raggio della chioma della matricina di riferimento. Anche in queste aree è stato eseguito un inventario particolareggiato dei polloni (georeferenzazione delle ceppaie, conta dei polloni per ceppaia, diametro di tutti i polloni per ceppaia e misurazione dell’altezza di 15 polloni per ceppaia) al fine di mettere in relazione questi parametri con il potere aduggiante della chioma delle matricine. A tal fine sono state individuate classi di distanza dal bordo delle chiome delle matricine di ampiezza di 1 m, indicando con numeri positivi le distanze crescenti dal bordo della chioma e con numeri negativi le distanze sotto copertura.
È stato infine determinato il diametro a m 1.30 di tutte le matricine presenti nelle aree di saggio.
Tutti i parametri rilevati sono stati infine sottoposti ad analisi statistica (ANOVA a due criteri di classificazione) e i confronti post-hoc sono stati eseguiti con il test HSD di Tukey (Honestly Significant Difference Test). Come software è stato utilizzato SAS System.
Risultati e discussione
Tesi “Ceduo”
I rilievi eseguiti hanno permesso di determinare, dopo 14 anni dal taglio, il numero di semenzali ad ettaro e le loro dimensioni, il numero di ceppaie morte, il numero medio di polloni per ceppaia, la mortalità annua dei polloni, il diametro medio, l’altezza media, l’area basimetrica e il volume dei polloni, nonché la distribuzione dei polloni al variare della loro distanza dal bordo chioma delle matricine.
Numero e dimensioni dei semenzali
La Tab. 1 mostra, per ogni area di saggio e per tesi, il numero di semenzali presenti dopo 14 anni dal taglio. Mediamente sono presenti poco più di 3000 semenzali, con una presenza maggiore dove il numero di matricine rilasciate è stato di 50 ha-1 e con una presenza minore dove il numero di matricine rilasciate dopo il taglio è stato di 250 matricine ha-1.
Tab. 1 - Numero di semenzali suddivisi per aree di saggio al 14° anno dal taglio del ceduo.
| ADS | Semenzali ha-1 |
Dcoll (mm) |
Htot (cm) |
Tesi | Semenzali ha-1 |
Dcoll (mm) |
Htot (cm) |
|---|---|---|---|---|---|---|---|
| 5.3 | 2250 | 9.89 | 51.44 | - | - | - | - |
| 6.3 | 3750 | 10.60 | 71.47 | A | 3500 | 10.07 | 57.75 |
| 7.3 | 4500 | 9.72 | 50.33 | - | - | - | - |
| 5.4 | 1750 | 7.86 | 43.86 | - | - | - | - |
| 6.4 | 3750 | 8.87 | 46.27 | B | 2500 | 8.07 | 46.08 |
| 7.4 | 2000 | 7.50 | 48.13 | - | - | - | - |
| 5.5 | 1750 | 9.00 | 41.57 | - | - | - | - |
| 6.5 | 5500 | 9.95 | 34.14 | C | 3250 | 9.58 | 41.94 |
| 7.5 | 2500 | 9.80 | 50.10 | - | - | - | - |
Ciò è facilmente spiegabile dalla minor copertura esercitata da un minor numero di matricine. Effetto che si esplica anche nelle dimensioni dei semenzali: infatti nella tesi A le dimensioni dei semenzali sono maggiori rispetto alle altre due tesi messe a confronto. In ogni transect di ogni area di saggio è stato riscontrato anche un alto numero di semenzali allo stato di plantula, mediamente 6000-7000 semenzali ha-1. Ad un’analisi statistica (ANOVA) non sono comunque risultate differenze significative nel numero di semenzali tra le varie tesi messe a confronto (Fig. 2).
Fig. 2 - Numero di semenzali ad ettaro.
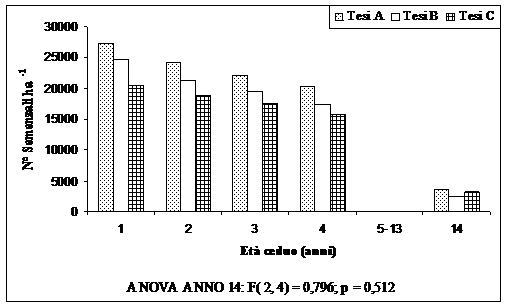
In Fig. 2, per un maggior confronto, si riportano anche i rilievi dei primi quattro anni, integrata dal rilievo al quattordicesimo anno. È riportata anche la figura relativa alla mortalità dei semenzali rispetto al primo rilievo (Fig. 3). È evidente come ci sia stata una forte selezione naturale dei semenzali, intensificatasi nel periodo compreso tra 5 e 13 anni dal taglio del ceduo. Al 14° anno mediamente sono presenti poco meno del 20% dei semenzali presenti al primo anno. Una mortalità leggermente più bassa vi è stata nella tesi C, dove il numero di matricine rilasciate è stato di 140 ha-1.
Fig. 3 - Presenze percentuali dei semenzali rispetto all’anno di inizio della sperimentazione.
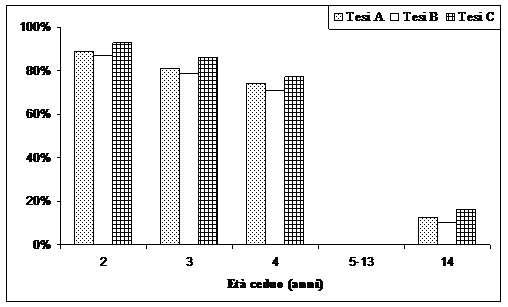
Sulla base dei risultati ottenuti è possibile affermare che la rinnovazione da seme è stata più abbondante dove il taglio è stato più intenso. Un elevato numero di piante rilasciate in piedi non ha originato una maggiore natalità di semenzali. Probabilmente molte piante rilasciate a dote del bosco hanno un effetto negativo, verosimilmente dovuto all’aduggiamento, sulla sopravvivenza delle giovani piantine.
Le Fig. 4 e Fig. 5 mostrano rispettivamente l’andamento delle dimensioni ipsodiametriche dei semenzali. I valori più alti, sia per i diametri che per le altezze, sono presenti nella tesi A. I valori diametrici più bassi sono presenti dove sono state rilasciate 250 matricine ha-1. In queste aree però i valori ipsometrici sono leggermente più alti rispetto alla tesi C. È interessante notare come la tesi A, dove è presente un minor numero di matricine, è caratterizzata sia da un maggior numero di semenzali che da dimensioni ipsodiametriche maggiori rispetto alle altre tesi. Ciò a conferma che un minor numero di portasemi rilasciate ha come conseguenza un maggior sviluppo dei semenzali, sia come numero che come dimensioni.
Fig. 4 - Diametro al colletto dei semenzali (valori medi ripartiti per tesi).
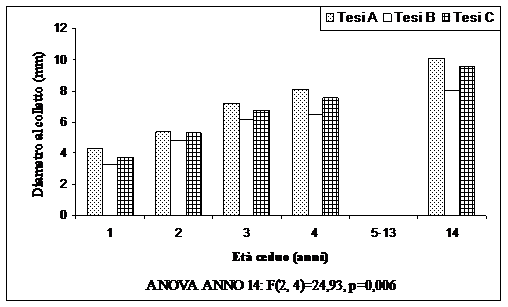
Fig. 5 - Altezza dei semenzali (valori medi ripartiti per tesi).
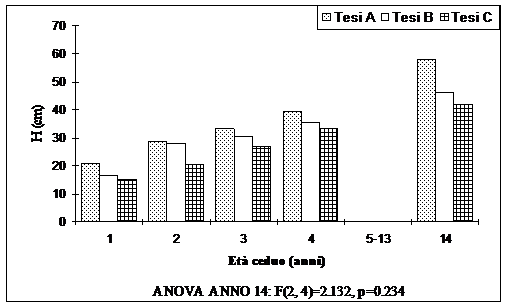
L’analisi della varianza ha evidenziato la significatività delle differenze tra le tesi poste a confronto solo per i diametri (Fig. 4), con differenze significative tra la Tesi A e la Tesi B, la Tesi C e la tesi B. Nessuna differenza significativa è invece presente tra la Tesi A e la Tesi C. Per le altezze non sono state riscontrate differenze significative tra le diverse tesi.
È evidente comunque come la crescita dei semenzali sia molto stentata e la mortalità dovuta alla competizione delle altre componenti del ceduo sia molto forte. Pochi semenzali, dopo 14 anni dal taglio del ceduo, superano i 50 cm di altezza e i 10 mm di diametro. D’altra parte i semenzali subiscono in maniera più forte anche la competizione delle diverse specie arbustive ed erbacee presenti nel sottobosco, oltre all’aduggiamento delle chiome sia delle matricine che dei polloni.
Sono da mettere in evidenza anche le modeste dimensioni ipsometriche nella tesi C (Ceduo composto). È probabile che qui l’ampia chioma delle matricine giochi un effetto significativo nella crescita modesta dei semenzali. In queste aree in effetti la copertura esercitata dalle chiome delle matricine è quasi continua su tutto il ceduo, mentre laddove sono state rilasciate 250 matricine, pur essendo in numero maggiore, queste hanno una chioma meno ampia e vi è maggiore spazio aereo tra una matricina e l’altra.
Non bisogna infine dimenticare che i danni meteorici avvenuti nel 1994 e 1995 hanno avuto i maggiori effetti proprio nelle aree dove sono state rilasciate 250 matricine ha-1, provocando spesso la perdita di parte della chioma, permettendo così una maggiore penetrazione della luce ([27], [28], [24], [26], [25]).
Si segnala infine che in seguito ai tagli non è stata rinvenuta una introduzione spontanea significativa di semenzali di specie forestali diverse dal leccio.
Numero di ceppaie morte
È utile ricordare che ancor prima di avviare la sperimentazione le aree risultavano confrontabili per quanto riguarda il numero di ceppaie comprensivo di quelle che erano “occupate” dalle matricine: mediamente erano di 1159 ceppaie ha-1 per la tesi A, di 1116 ha-1 per la tesi B e di 1131 ha-1 per la tesi C ([27]). Al primo rilievo eseguito tutte le ceppaie avevano emesso polloni vivi. In occasione del secondo rilievo, invece, sono state riscontrate due ceppaie che non avevano più polloni. Di queste, una ricadeva nelle parcella Tesi A e una nella parcella Tesi B. Nei rilievi successivi, in queste due ceppaie, non è stata riscontrata nessuna emissione di nuovi polloni. Al 14° anno nessun’altra ceppaia è morta.
Mediamente, a 14 anni dal taglio, sono presenti 1108 ceppaie ha-1 nelle parcelle dove sono state rilasciate 50 matricine ad ettaro, 865 ceppaie ha-1 nelle parcelle dove sono state rilasciate 250 matricine ad ettaro e 991 ceppaie ha-1 nelle parcelle dove sono state rilasciate 140 matricine ad ettaro.
Numero di polloni per ceppaia
La Tab. 2 mostra la variazione del numero di polloni per ceppaia per i primi 4 anni e al 14° anno. È evidente la diminuzione del numero di polloni per ceppaia dovuta alla selezione naturale nel corso del periodo di sperimentazione. Il numero di polloni vivi per ceppaia (Tab. 2), al primo anno dopo la ceduazione, è risultato in media pari a 33.1, con valori significativamente diversi a seconda del numero di matricine rilasciate a dote del bosco ([27]). Valori più elevati sono stati osservati dove è stato rilasciato un minor numero di matricine (Tesi A).
Tab. 2 - Variazione del numero di polloni per ceppaia. Valori medi per tesi. ANOVA Anno 14: F(2, 4) = 1.319, p = 0.271.
| Età (anni) | 1 | 2 | 3 | 4 | 14 |
|---|---|---|---|---|---|
| tesi A | 36.6 | 35.8 | 30.8 | 24.5 | 17.6 |
| tesi B | 32.0 | 31.2 | 27.5 | 24.1 | 14.4 |
| tesi C | 30.6 | 27.8 | 24.1 | 18.4 | 13.7 |
| Media | 33.1 | 31.6 | 27.4 | 22.4 | 15.2 |
Al quattordicesimo anno il numero di polloni per ceppaia si riduce mediamente a 15.2. Più in particolare il numero di polloni per ceppaia è di 17.6 polloni nella tesi A, di 14.4 polloni nella tesi B e di 13.7 polloni per ceppaia nella tesi C.
La tesi A pur avendo un più alto numero di polloni per ceppaia, non presenta alcuna differenza significativa rispetto alle altre tesi messe a confronto (Tab. 2). Al quattordicesimo anno dal taglio si annullano le differenze significative che esistevano negli anni immediatamente successivi al taglio.
Anche prendendo in considerazione il numero totale di polloni ad ettaro, l’analisi della varianza non ha evidenziato differenze significative tra le tesi ceduo poste a confronto. Mediamente nella tesi A sono presenti circa 20000 polloni ha-1, circa 12500 nella tesi B e circa 13600 nella tesi C.
Mortalità dei polloni
L’analisi della varianza applicata ai dati di mortalità dei polloni tra la prima stagione vegetativa e l’anno dell’ultimo rilievo (14 anni dal taglio), non ha evidenziato differenze significative tra i tre trattamenti messi a confronto. La mortalità dovuta alla selezione naturale ha comunque maggiormente interessato i polloni situati nelle aree a Tesi B: qui infatti la mortalitàè risultata pari al 57.5% rispetto al numero di polloni presenti al primo anno dopo il taglio. Tale valore scende al 48.5% per i polloni situati nelle aree della Tesi A ed è pari al 53% per i polloni situati nelle aree della Tesi C.
Pur se dal punto di vista statistico non vi sono differenze significative nella mortalità, è comunque evidente una tendenza: all’aumentare della copertura esercitata dalle matricine, aumenta anche la mortalità dei polloni.
Diametro medio ed altezza media dei polloni
A 14 anni dal taglio i polloni di leccio hanno dimensioni ipsodiametriche piuttosto modeste (Fig. 6 e Fig. 7): infatti il diametro a m 1.30 dei polloni, in media cm 3.35, varia da cm 3.20 per le aree dove è stato rilasciato un maggior numero di portasemi a dote del bosco (Tesi B), a cm 3.61 per le aree dove il numero di matricine rilasciate è stato di 50 ad ettaro (Tesi A).
Fig. 6 - Diametro dei polloni (valori medi ripartiti per tesi).
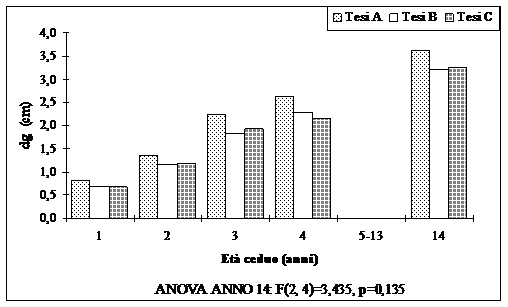
Fig. 7 - Altezza dei polloni (valori medi ripartiti per tesi).
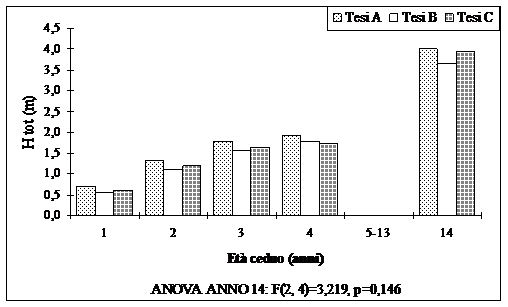
Anche le altezze sono modeste. In media l’altezza dei polloni dopo 14 anni dal taglio, è pari a 3.88 m. Un’ altezza maggiore è presente dove il numero di matricine rilasciate a dote del bosco è stato minore. In queste aree le altezze dei polloni raggiungono i 4.0 m.
Mediamente i polloni di leccio sono cresciuti di appena 0.24 cm all’anno (incremento diametrico) e gli incrementi medi di altezza sono stati mediamente di 0.28 m all’anno. I dati, sottoposti ad analisi della varianza, non hanno evidenziato differenze significative (Fig. 6 e Fig. 7).
Rimane comunque da sottolineare le maggiori dimensioni che hanno raggiunto i polloni nelle aree dove il numero di matricine rilasciate a dote del bosco è stato minore.
Una ulteriore considerazione è possibile fare. Nei primi anni dopo la ceduazione, la rinnovazione agamica tende a riconquistare velocemente lo spazio aereo. Al quarto anno, infatti, l’altezza dei polloni supera i 2.0 m. In seguito l’accrescimento dei polloni, sia in diametro che in altezza, procede molto lentamente. Prendendo in considerazione l’incremento di altezza (ma analogo andamento si nota nei riguardi del diametro), questo parametro al passare degli anni diminuisce sempre di più. Al secondo anno l’incremento medio di altezza è pari a 0.61 m, al terzo anno è pari a 0.55 m, al quarto anno è pari a 0.45 m e al quattordicesimo anno è pari a 0.28 m. Dunque a una prima fase di veloce accrescimento dei polloni, segue una fase di accrescimento molto lenta.
Area basimetrica e volume dei polloni
Tab. 3 - Variazione dei valori di area basimetrica. Valori medi per tesi (dati ad ettaro). ANOVA Anno 14 G: F(2, 4)=4.688, p=0.0894. ANOVA Anno 14 V: F(2, 4)=10.985, p=0.0236
| Età | Tesi A G (m2) |
Tesi B G (m2) |
Tesi C G (m2) |
Tesi A V (m3) |
Tesi B V (m3) |
Tesi C V(m3) |
|---|---|---|---|---|---|---|
| 4 anni | 3.00 | 1.75 | 1.35 | 12.06 | 6.94 | 5.29 |
| 14 anni | 20.43 | 11.22 | 11.15 | 91.89 | 49.87 | 49.60 |
| Variazione | 17.42 | 9.46 | 9.80 | 79.83 | 42.93 | 44.31 |
La Tab. 3 mostra i valori di area basimetrica e di volume ad ettaro dei polloni al quarto e al quattordicesimo anno. Per il calcolo dell’area basimetrica, i valori dei diametri a m 1.30 al quarto anno sono stati ricavati dall’analisi delle coppie di valori “diametro al colletto - diametro a m 1.30” rilevati in occasione del rilievo del quarto anno. L’equazione che esprime la migliore relazione tra le due variabile è una retta (eqn. 1):
Per la determinazione dei volumi è stata utilizzata l’equazione ricavata dalla tavola di cubatura costruita per questi soprassuoli da [27]. L’equazione utilizzata è (eqn. 2):
Nelle aree dove il rilascio di matricine è stato di 50 ad ettaro si è avuto l’incremento periodico maggiore, qui infatti il valore è passato da 3 m2 all’età di 4 anni, a circa 20 m2 a 14 anni di età, con un incremento periodico nei 10 anni di circa 17 m2 ha-1. L’incremento periodico nelle aree riferibili alla tesi B è stato di circa 9.5 m2 ha-1, passando, sempre per le stesse età, da 1.75 m2 a 11.22 m2. Nella tesi C l’area basimetrica è passata da 1.35 m2 a 11.15 m2, con un incremento periodico di circa 10 m2 ha-1.
La tesi che presenta un’area basimetrica maggiore è il ceduo con rilascio di 50 matricine ad ettaro. Questo valore è determinato dal maggior numero di polloni per ceppaia e per ettaro e dalle maggiori dimensioni diametriche rispetto alle altre tesi riguardanti il ceduo.
All’età di 14 anni il ceduo presenta un valore dei volumi dei polloni abbastanza diverso tra la tesi A e le restanti due tesi. Nella tesi A i polloni hanno un volume di circa 92 m3 ha-1 contro circa i 50 m3 ha-1 delle altre due tesi (Tab. 3). Il maggior volume presente nel ceduo con rilascio di 50 matricine ad ettaro è chiaramente correlato al maggior numero di polloni e alle maggiori dimensioni diametriche di questi, rispetto alle altre due tesi ceduo prese in esame. L’analisi della varianza ha confermato la significatività delle differenze tra la tesi A e le restanti due tesi. Nessuna differenza significativa invece vi è tra la tesi B e la tesi C.
La tesi A, invece, si differenzia in maniera significativa sia per quanto riguarda la produttività attuale, sia per quanto riguarda l’incremento periodico dal quarto al quattordicesimo anno, presentando valori pari quasi al doppio rispetto alle altre due tesi ceduo esaminate (Tab. 3). Nelle aree trattate a ceduo matricinato con rilascio di 50 matricine ad ettaro, l’incremento corrente medio, nel periodo di 10 anni, è stato di circa 8 m3 annui ha-1, mentre nelle aree trattate a ceduo composto e a ceduo matricinato con rilascio di 250 portasemi ad ettaro gli incrementi correnti medi sono stati rispettivamente di 4.4 m3 annui ha-1 e di 4.3 m3 annui ha-1.
Accrescimento polloni a diverse distanza dal bordo chioma delle matricine
La Fig. 8, la Fig. 9 e la Fig. 10 mostrano rispettivamente la variazione del numero di polloni per ceppaia, il diametro medio e l’altezza media, al variare delle ceppaie dal bordo chioma delle matricine. Si ricorda che i numeri negativi indicano, in metri, le ceppaie sotto copertura dal bordo chioma delle matricine, e i numeri positivi distanze crescenti dal bordo chioma delle matricine.
Fig. 8 - Numero di polloni per ceppaia al variare della distanza dal bordo chioma delle matricine.
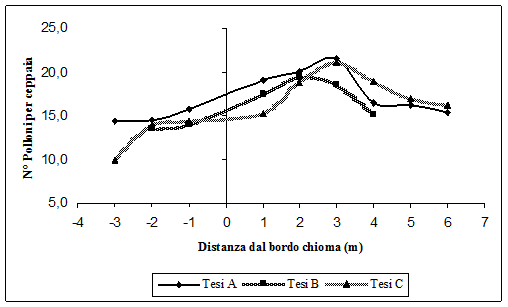
Fig. 9 - Diametro medio dei polloni al variare della distanza dal bordo chioma delle matricine.
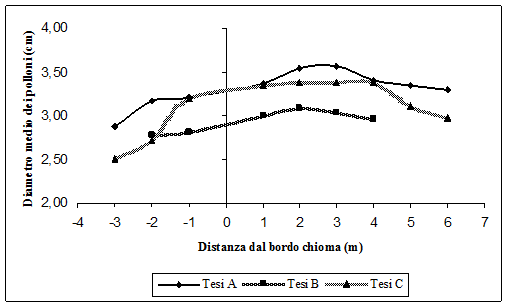
Fig. 10 - Altezza media dei polloni al variare della distanza dal bordo chioma delle matricine.
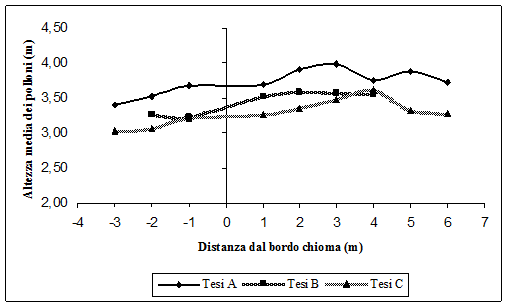
Per i tre parametri analizzati è evidente come l’accrescimento sia più stentato sotto chioma rispetto ai polloni che vegetano fuori dalla chioma delle matricine. Sembra dunque confermato che l’aduggiamento della chioma delle matricine ha un ruolo fondamentale nell’accrescimento dei polloni. Il fenomeno è più accentuato nel caso del numero di polloni per ceppaia (Fig. 8). Questi aumentano sempre di più andando dalle ceppaie sotto chioma verso quello più esterne, fino a circa 3 m di distanza dal bordo chioma (Tesi A e C), dove il numero di polloni per ceppaia è massimo. Dopo tale distanza il numero di polloni tende a diminuire, in quanto le ceppaie si trovano sotto copertura della chioma delle matricine immediatamente più vicine. Per la Tesi B invece il numero massimo di polloni si raggiunge a 2 m dal bordo chioma, poi il numero di polloni per ceppaia diminuisce. Ciò è dovuto al maggiore numero di matricine qui presenti e dunque già a pochissimi metri di distanza dal bordo chioma, le ceppaie vengono a trovarsi sotto la chioma della matricina immediatamente più vicina.
Lo stesso fenomeno si verifica sia per le dimensioni diametriche (Fig. 9) che per quelle ipsometriche (Fig. 10). I polloni sotto chioma hanno dimensioni minori rispetto a quelli fuori chioma. Anche qui i valori massimi si raggiungono a distanze di 2-3 metri dal bordo chioma. Se a questo si aggiunge che i polloni presenti nella Tesi A, e cioè dove è stato rilasciato un minor numero di piante a dote del bosco, sono sia in numero superiore rispetto alle altre tesi che di dimensioni maggiori, diventa assai probabile che l’aduggiamento provocato dalla chioma delle matricine inibisce la crescita dei polloni.
Matricine
Lo studio sulle aree riferibili alle Tesi ceduo ha interessato anche le dinamiche di accrescimento delle matricine, e la Tab. 4 mostra l’evoluzione dei parametri dendrometrici delle portasemi.
Tab. 4 - Parametri dendrometrici delle matricine alle varie età considerate (dati ad ettaro).
| Età | Tesi | Piante [Numero] |
Area Basim. [m2] |
Dg [cm] |
Vol [m3] |
|---|---|---|---|---|---|
| 1 anno | Tesi A | 50 | 1.00 | 16.0 | 5.23 |
| Tesi B | 250 | 3.10 | 12.6 | 15.83 | |
| Tesi C | 140 | 3.12 | 16.8 | 16.42 | |
| 2 anni | Tesi A | 31 | 0.71 | 17.0 | 3.72 |
| Tesi B | 127 | 1.72 | 13.1 | 8.84 | |
| Tesi C | 123 | 2.76 | 16.9 | 14.54 | |
| 3 anni | Tesi A | 31 | 0.81 | 18.2 | 4.29 |
| Tesi B | 127 | 1.92 | 13.9 | 9.91 | |
| Tesi C | 123 | 3.01 | 17.7 | 15.92 | |
| 4 anni | Tesi A | 31 | 0.89 | 19.0 | 4.74 |
| Tesi B | 127 | 1.97 | 14.1 | 10.18 | |
| Tesi C | 123 | 3.22 | 18.3 | 17.08 | |
| 14 anni | Tesi A | 31 | 1.31 | 23.1 | 7.13 |
| Tesi B | 121 | 3.75 | 19.9 | 20.04 | |
| Tesi C | 129 | 5.10 | 22.4 | 27.64 | |
| Variazioni | Tesi | Rispetto al 1° anno |
Rispetto al 2° anno |
Rispetto al 2° anno |
Rispetto al 2° anno |
| Tesi A | -19 | 0.60 | 6.10 | 3.41 | |
| Tesi B | -129 | 2.03 | 6.80 | 11.20 | |
| Tesi C | -11 | 2.34 | 5.50 | 13.10 |
È stata registrata una diminuzione notevole delle piante portaseme rilasciate a causa dei danni provocati dagli eventi meteorici durante il secondo anno di sperimentazione ([27]).
Nella tesi A dopo il secondo anno non è stato registrato nessun ulteriore danno a carico delle matricine. Il numero attuale di portasemi è di 31 piante ad ettaro. La tesi B, dove sono stati registrati il maggior numero di danni in seguito agli eventi meteorici, ha subito ulteriori danni nel periodo tra il quarto e il quattordicesimo anno di sperimentazione, qui infatti sono state trovate mediamente 6 matricine ad ettaro con il fusto spezzato. Il numero di matricine è dunque passato da 250 ad ettaro a 127 ad ettaro dopo gli eventi meteorici, a 121 ad ettaro all’età attuale. Nella tesi C il numero di matricine sane dopo gli eventi meteorici era pari a 123 ad ettaro. Nelle aree riferibili a questa tesi si è verificato che alcune matricine che avevano subito dei danni alla chioma, negli anni successivi al quarto, hanno quasi ricostituito completamente la chioma, per cui sono state considerate sane. Attualmente il numero di matricine è di 129 ad ettaro.
I maggiori danni si sono avuti dove il rilascio di matricine è stato di 250 ad ettaro (Tesi B): qui le piante danneggiate nei 14 anni di sperimentazione sono state 129, mentre i danni minori si sono avuti nel ceduo composto (Tesi C) con una perdita totale di 11 piante ad ettaro. Probabilmente il maggior diametro delle matricine presenti nelle aree a ceduo composto ha contribuito ad una maggiore stabilità del soprassuolo.
L’area basimetrica delle tesi A e B, dopo gli eventi meteorici, dal secondo al quarto anno risulta inferiore a quella registrata durante il primo anno di sperimentazione. La tesi C ha subito una minor perdita di area basimetrica dopo il secondo anno, al quarto anno l’incremento periodico risultava di 0.10 m2 ha-1.
In Tab. 4 è mostrato anche il volume delle matricine nelle varie tesi considerate, durante i 14 anni dall’inizio della sperimentazione. Il volume è stato calcolato con la tavola di cubatura appositamente costruita per questi soprassuoli ([27]).
Nella tesi A e nella tesi B, a causa dei danni provocati dai suddetti eventi meteorici, il volume delle matricine al secondo, terzo e quarto anno è inferiore a quello registrato al primo anno. Nel ceduo con rilascio di 50 matricine ad ettaro (Tesi A) il volume è passato da 5.23 m3 ha-1 al primo anno a 4.74 m3 ha-1 al quarto anno, nel ceduo con rilascio di 250 matricine ad ettaro (Tesi B) la perdita di volume dal primo al quarto anno è stata di 5.65 m3 ha-1 passando da un valore di 15.83 m3 ha-1 a 10.18 m3 ha-1. Nella tesi C invece, la perdita di volume tra il rimo e il secondo anno, è stata di 1.88 m3 ha-1 ma già al quarto anno si è registrato, rispetto al primo anno, un incremento periodico di 0.66 m3 ha-1.
In termini di crescita di volume e di area basimetrica delle matricine, la tesi in cui si sono riscontrati i maggiori incrementi periodici tra il secondo e il quattordicesimo anno risulta essere il ceduo composto: in queste aree l’incremento periodico di volume tra i due periodi è stato di 13.1 m3 ha-1, mentre quello di area basimetrica è stato di 2.34 m2 ha-1.
Alla luce di questi di dati si può affermare che sono state le matricine presenti nelle aree della tesi C quelle che hanno registrato gli incrementi maggiori rispetto alle altre tesi considerate sia come area basimetrica che come volume.
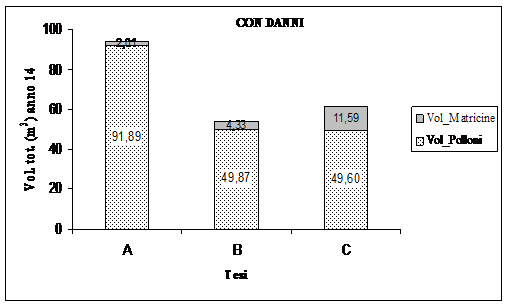
È stato eseguito il confronto sulla produttività delle tesi attraverso il calcolo, al quattordicesimo anno di sperimentazione, del volume complessivo del soprassuolo comprensivo sia del volume delle matricine che del volume dei polloni. Questo per valutare i rapporti tra intensità di matricinatura e sviluppo del soprassuolo. La Fig. 11 mostra come nei soprassuoli cedui in esame il volume totale (polloni e matricine) sia correlato negativamente al numero di piante portaseme rilasciate a dote del bosco. Al quattordicesimo anno il volume totale prodotto è maggiore nelle aree riferibili alla tesi A (scarsa intensità di matricinatura), segue la tesi C (ceduo composto) con circa 60 m3 ha-1 di volume totale e per ultima la tesi B (ceduo intensamente matricinato) con circa 54 m3 ha1.
Fig. 11 - Ripartizione della produttività totale realmente registrata dai polloni e dalle matricine al netto del volume della “dote” (dati ad ettaro).
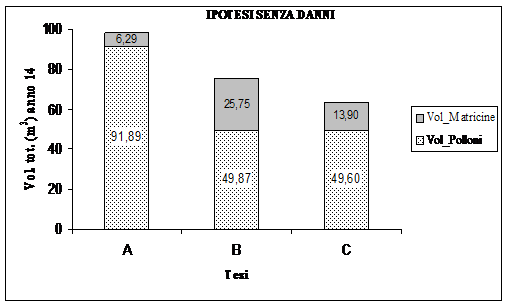
In effetti, come si è detto, già al secondo anno dall’avvio della sperimentazione, una tromba d’aria ha danneggiato in misura diversa le matricine rilasciate a dote del bosco nelle aree sperimentali. Si riporta una simulazione della crescita (Fig. 12) che si sarebbe avuta nelle varie tesi se le matricine non fossero state danneggiate. Il contributo incrementale delle matricine è stato desunto dall’incremento di volume mediamente registrato da ogni portaseme nel periodo considerato.
Fig. 12 - Ripartizione della produttività totale che si sarebbe avuta nell’ipotesi della simulazione di cui sopra, al netto del volume della “dote” (dati ad ettaro).
Tesi “Conversione ad alto fusto” e “Testimone”
Prima di passare all’analisi dei risultati sembra opportuno ricordare che l’algoritmo colturale seguito per l’attuazione del taglio di avviamento è stato quello della conversione con il metodo del rilascio intensivo di allievi, con il rilascio sulle ceppaie di un pollone o al massimo due, scelti fra quelli fenotipicamente migliori. La massa asportata, pari mediamente a 102.15 m3 ha-1, rappresentava circa il 48% della massa presente prima del taglio.
Inoltre, il taglio di avviamento all’alto fusto non ha originato una introduzione spontanea significativa di specie forestali diverse dal leccio e dunque non ha avuto alcun effetto sulla diversità specifica. Le poche specie di semenzali rinvenuti a un anno dal taglio (soprattutto Fraxinus ornus e Ostrya Carpinifolia) non sono più state rinvenute nei rilievi successivi.
La Tab. 5 e la Tab. 6 mostrano l’evoluzione dei parametri dendrometrici medi delle aree sottoposte a conversione ad alto fusto (Tesi D) e delle aree lasciate ad evoluzione naturale (Tesi E). I dati esposti devono essere esaminati tenendo in considerazione gli eventi meteorici avvenuti durante il secondo anno di sperimentazione e, appunto per ciò, la riga degli incrementi si riferisce agli incrementi registrati dal secondo al quattordicesimo anno.
Tab. 5 - Parametri dendrometrici medi riferiti alla Tesi D - Conversione ad alto fusto (dati ad ettaro).
| Età (anni) |
Numero Ceppaie |
Numero polloni/ha |
Numero Poll/Cepp |
G ha-1 (m²) |
Dg (cm) |
Vol ha-1 (m³) |
|---|---|---|---|---|---|---|
| 1 | 1171 | 2031 | 1.7 | 21.95 | 11.9 | 110.8 |
| 2 | 1113 | 1619 | 1.4 | 16.74 | 11.7 | 84.8 |
| 4 | 1092 | 1574 | 1.4 | 21.54 | 13.4 | 110.6 |
| 14 | 1065 | 1410 | 1.3 | 27.91 | 16.1 | 146.1 |
| Variazione | Rispetto al 1° anno | Rispetto al 1° anno | - | Rispetto al 2° anno | Rispetto al 2° anno | Rispetto al 2° anno |
| -106 | -621 | - | 11.70 | 4.4 | 61.3 |
Tab. 6 - Parametri dendrometrici medi riferiti alla Tesi E - Testimone (dati ad ettaro).
| Età (anni) | Numero Ceppaie |
Numero polloni/ha |
Numero Poll/Cepp |
G ha-1 (m²) |
Dg (cm) |
Vol ha-1 (m³) |
|---|---|---|---|---|---|---|
| 1 | 1159 | 6790 | 5.9 | 48.69 | 9.6 | 241.9 |
| 2 | 1150 | 6594 | 5.8 | 46.40 | 9.5 | 230.3 |
| 4 | 1142 | 6540 | 5.8 | 53.41 | 10.2 | 267.1 |
| 14 | 1102 | 5138 | 4.7 | 54.97 | 11.7 | 278.7 |
| Variazione | Rispetto al 1° anno | Rispetto al 1° anno | - | Rispetto al 2° anno | Rispetto al 2° anno | Rispetto al 2° anno |
| -56 | -1652 | - | 8.57 | 2.2 | 48.4 |
Numero di ceppaie
Per quanto riguarda le aree sottoposte a Conversione ad alto fusto, vi è stata una diminuzione di 106 ceppaie ha-1. Su queste ceppaie, al rilievo del 2007, non era presente più nessun pollone vivo. Nelle aree lasciate ad evoluzione naturale il numero di ceppaie che rispetto al primo rilievo non avevano più nessun pollone vivo è stato di 56 ceppaie ha-1.
Il motivo per cui nelle aree sottoposte a Conversione ad alto fusto vi è stata una mortalità delle ceppaie quasi doppia rispetto alle aree lasciate ad evoluzione naturale, è spiegabile dai maggiori danni che si sono verificati nelle aree in conversione, dopo i due eventi meteorici a carattere eccezionale che vi sono stati nel 1994 e nel 1995 ([27]). La diminuzione del numero di ceppaie con polloni vivi è avvenuta proprio tra il primo e il secondo anno dall’avvio della sperimentazione, periodo in cui si sono verificati i due eventi meteorici a carattere eccezionale. Si ricorda infatti che i danni nelle aree in conversione hanno interessato circa il 20% delle piante in piedi, mentre nelle aree testimoni i danni hanno colpito il 3% delle piante in piedi ([27]).
Ai fini di una maggiore comprensione del fenomeno, sembra utile comunque ricordare che i danni maggiori verificatisi nelle aree in Conversione sono dovuti essenzialmente ad una sola delle 3 aree sottoposte a Conversione ad alto fusto. Dopo gli eventi meteorici in una di queste aree il numero di ceppaie ad ettaro con polloni vivi è diminuito di 169 unità, nelle altre due aree invece la diminuzione del numero di ceppaie ad ettaro con polloni vivi è stata rispettivamente di 6 unità per un’area e di 0 (zero) unità per l’altra.
Se si escludono i danni dovuti agli eventi meteorici, la diminuzione del numero di ceppaie non è stata significativa in nessuna delle due situazioni messe a confronto. La mortalità avutasi rientra nella evoluzione naturale dei popolamenti.
Numero di polloni ad ettaro e numero di polloni per ceppaia
Il numero di polloni morti dopo 14 anni dall’avvio della sperimentazione è stato di notevole entità per le aree lasciate ad evoluzione naturale (1652 polloni ha-1) e di media intensità per le aree sottoposte a Conversione ad alto fusto (621 polloni ha-1). Al quattordicesimo anno il numero di polloni ad ettaro è di 1410 unità per le conversioni e di 5138 unità per le aree lasciate ad evoluzione naturale. I polloni morti hanno interessato soprattutto le aree lasciate ad evoluzione naturale; col taglio di avviamento all’alto fusto, invece, vi è stata già una selezione dei migliori polloni e, conseguentemente, una maggiore stabilità.
Nelle aree lasciate ad evoluzione naturale la maggiore competizione tra le piante ha favorito di conseguenza una maggiore mortalità, con una notevole perdita di biomassa e un notevole accumulo di necromassa per terra, necromassa che se da un lato ha la sua innegabile valenza bio-ecologica, dall’altra può costituire anche un pericoloso veicolo di incendi soprattutto in queste zone del Gargano dove gli incendi sono tutt’altro che un evento raro.
A causa della mortalità dei polloni e della competizione tra polloni della stessa ceppaia, anche il numero di polloni per ceppaia col tempo è diminuito. Mediamente dopo 14 anni dall’avvio della sperimentazione nelle aree in conversione sono presenti circa 1.3 polloni per ceppaia (1.7 polloni per ceppaia agli inizi della sperimentazione), mentre nelle aree ad evoluzione naturale sono presenti mediamente 4.7 polloni per ceppaia (5.9 polloni per ceppaia agli inizi della sperimentazione).
Area basimetrica, diametro medio e volume
Nelle aree sottoposte a Conversione, l’area basimetrica ha subito una forte diminuzione nel secondo anno (a causa degli eventi meteorici), con differenze tra i valori prima e dopo gli eventi meteorici di 5.21 m2 ha-1. In seguito vi è stato un buon incremento in tutte le aree sottoposte a Conversione (Fig. 13). L’area basimetrica è passata da circa 17 m2 ha-1 del secondo anno a circa 28 m2 ha-1 a 14 anni dall’avvio della sperimentazione, con un incremento periodico pari a 11.70 m2 ha-1.
Fig. 13 - Sviluppo dell’area basimetrica ad ettaro nelle aree a conversione e nelle aree ad evoluzione naturale.
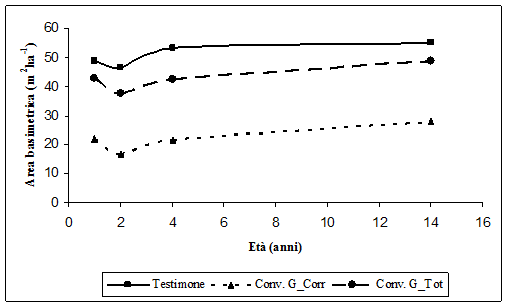
Nelle aree lasciate ad evoluzione naturale (Fig. 13) la diminuzione dei valori di area basimetrica tra il primo e secondo anno è stata minore, a causa dei minori danni che qui si sono verificati, rispetto alle aree in Conversione: qui infatti la diminuzione di area basimetrica è stata di 2.29 m2 ha-1. In seguito gli incrementi sono stati di media entità. L’area basimetrica è passata da circa 46 m2 ha-1 del secondo anno a circa 55 m2 ha-1 a 14 anni dall’avvio della sperimentazione, con un incremento periodico pari a 8.57 m2 ha-1.
In Fig. 13 è riportata anche la curva riferita alla massa totale in termini di area basimetrica nelle aree sottoposte a conversione ad alto fusto; tale curva è stata ottenuta dalla somma dell’area basimetrica corrente e dell’area basimetrica riferita alle piante asportate con l’intervento di conversione. Come si può osservare, i valori riferiti alla massa totale in termini di area basimetrica della tesi Conversione risultano inferiori ai valori della tesi Testimone. Questo perché già prima dell’asportazione di biomassa riferibile all’inizio della sperimentazione l’area basimetrica della tesi Conversione ad alto fusto era inferiore a quella registrata nella tesi Testimone.
I dati di incremento periodico di area basimetrica, sottoposti ad analisi della varianza, hanno evidenziato differenze significative tra le due tesi poste a confronto.
Il diametro medio (Tab. 5) delle piante nelle aree in conversione è passato da 11.9 cm al primo anno di sperimentazione a 16.1 cm al quattordicesimo anno, con un incremento diametrico pari a 4.2 cm. Nella aree ad evoluzione naturale nello stesso periodo di cui sopra il diametro medio è passato da 9.6 cm a 11.7 cm, con un incremento diametrico pari a 2.1 cm. Nelle aree in conversione l’aumento del diametro medio risulta più forte rispetto alle aree lasciate ad evoluzione naturale.
Anche in questo caso i dati, sottoposti ad analisi della varianza, hanno mostrato differenze molto significative tra le tesi poste a confronto.
Per quanto i riguarda i volumi ad ettaro (Fig. 14), nelle aree sottoposte a Conversione il volume ad ettaro ha subito una forte diminuzione nel secondo anno, con differenze tra i valori prima e dopo gli eventi meteorici di 26 m3 ha-1. Le aree hanno comunque risposto positivamente al trattamento impartito: il volume è passato da circa 85 m3 ha-1 del secondo anno a circa 146 m3 ha-1 a 14 anni dall’avvio della sperimentazione, con un incremento periodico pari a 61.3 m3 ha-1.
Fig. 14 - Sviluppo del volume ad ettaro nelle aree a conversione e nelle aree ad evoluzione naturale.
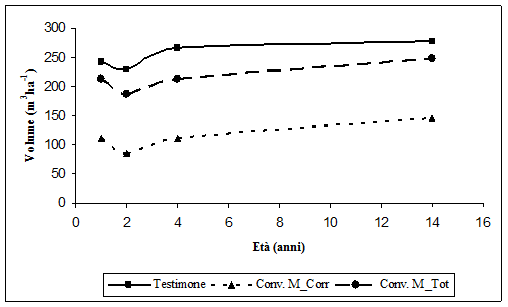
Nelle aree lasciate ad evoluzione naturale la diminuzione dei valori di volume tra il primo e secondo anno è stata di circa 12 m3 ha-1, sempre a causa degli avversi eventi meteorici. Dopo il secondo anno gli incrementi sono stati di modesta entità. Il volume è passato da circa 230 m3 ha-1 del secondo anno a circa 279 m3 ha-1 a 14 anni dall’avvio della sperimentazione, con un incremento periodico pari a 48.40 m3 ha-1.
In Fig. 14 è riportata anche la curva riferita alla massa totale in termini di volume della tesi Conversione ad alto fusto; come per l’area basimetrica tali valori sono stati ottenuti attraverso la somma del volume corrente e del volume asportato all’inizio della sperimentazione. La figura mostra come i valori di massa totale in termini di volume della tesi Conversione ad alto fusto risultino inferiori ai valori riferiti alla tesi Testimone, questo perché già prima del taglio di conversione le aree appartenenti alla tesi Conversione facevano registrare volumi inferiori alle aree appartenenti alla tesi Testimone.
Anche in questo caso i dati di incremento periodico di volume, sottoposti ad analisi della varianza, hanno evidenziato differenze significative tra le due tesi poste a confronto.
Conclusioni
Lo studio condotto in un ceduo di leccio sul Promontorio del Gargano ha consentito di rilevare gli effetti di diverse scelte selvicolturali: evoluzione naturale, conversione ad alto fusto, ceduo matricinato con 50 matricine ad ettaro, ceduo matricinato con 250 matricine ad ettaro e ceduo composto con 140 matricine ad ettaro.
I risultati ottenuti nelle aree sottoposte ad interventi di conversione e in quelle lasciate ad evoluzione naturale hanno messo in evidenza la maggiore convenienza della prima scelta colturale. Gli incrementi registrati nelle aree sottoposte a conversione risultano nettamente superiori rispetto ai valori calcolati per le aree lasciate ad evoluzione naturale. In base ai valori di incremento diametrico e al numero di polloni ad ettaro, si può ipotizzare una maggiore stabilità del soprassuolo sottoposto a conversione rispetto a quello lasciato ad evoluzione naturale. Inoltre la notevole quantità di necromassa presente nelle aree lasciate ad evoluzione naturale, dovuta agli effetti della selezione naturale, rende queste ultime maggiormente sottoposte al rischio di incendi, evento frequente in questa zona. La scelta di operare una selezione mirata sulle ceppaie, con l’intervento di conversione ad alto fusto, e conseguente asportazione del materiale legnoso diminuisce notevolmente questo rischio. Presi in considerazione gli elementi relativi all’incremento, alla stabilità e al rischio incendi, la scelta di operare una conversione guidata del ceduo verso la fustaia, risulta preferibile rispetto all’evoluzione naturale. Ciò nonostante permangono non poche perplessità sulle condizioni pedologiche della stazione e sulla possibilità di poter sostenere da un punto di vista ecofisiologico la futura fustaia.
L’alternativa alla conversione ad alto fusto è il mantenimento del governo a ceduo, considerando anche le condizioni socio-economiche del territorio in esame, dove tale scelta colturale offre un prodotto ancora richiesto dal mercato, oltre che rappresentare la forma di governo utilizzata tradizionalmente ([27]).
Con la presente sperimentazione si è cercato di definire quale possa essere il trattamento più idoneo al mantenimento di tale forma di governo.
Per ciascuna delle ipotesi di gestione proposte sono state fatte delle considerazioni sulla rinnovazione gamica, sulla componente agamica e sulla produttività totale.
L’analisi sulla componente gamica ha evidenziato che i semenzali nascono immediatamente prima e subito dopo il taglio di maturità: a distanza di 14 anni non sono stati rilevati nuovi semenzali sviluppati. Si sono però registrati semenzali allo stato di plantula. È da supporre che questi semenzali non riescano a svilupparsi; infatti tutti i semenzali allo stato di plantula trovati avevano tutti un anno di età o al massimo due anni di età. La mortalità dei semenzali è stata notevole. All’età di 14 anni il numero di semenzali ad ettaro oscilla tra 2500 e 3500. Subito dopo il taglio di utilizzazione il numero di semenzali ad ettaro era superiore a 24000. Le aree dove la percentuale di semenzali vivi rispetto al primo anno è maggiore appartengono alla tesi C (ceduo composto). Le dimensioni dei semenzali sono inferiori ai 10 mm per quanto riguarda i diametri e ai 50 cm per le altezze. Pertanto allo stato attuale non sussistono garanzie per quanto riguarda la loro sopravvivenza. È utile infine ribadire che nella Tesi A (50 matricine ad ettaro) si sono registrate le dimensioni ipsodiametriche più elevate, come anche il più alto numero di semenzali.
Per quanto riguarda la componente agamica del soprassuolo, il numero di polloni (totali e medio per ceppaia), diminuisce col tempo in maniera molto graduale. I polloni sembrano essere molto resistenti sia alle avversità che alla competizione intraindividuale. All’età di 14 anni il numero di polloni vivi rispetto al primo anno oscilla mediamente tra il 42% riferibile alla tesi B e il 51% nella tesi A, con differenze non significative tra i trattamenti messi a confronto. Attualmente il numero medio di polloni per ceppaia è di quasi 16, e i valori più elevati sono stati riscontrati dove il rilascio di matricine è stato minore. Quando il ceduo appartenuto al precedente ciclo fu utilizzato, il numero medio di polloni per ceppaia era di circa 5.5. C’è da ritenere dunque che il numero di polloni per ceppaia debba ulteriormente diminuire.
Gli incrementi ipso-diametrici hanno messo in evidenza la lentezza di accrescimento dei polloni di leccio. A 14 anni dal taglio il diametro medio dei polloni è pari a circa 3.3 cm e l’altezza media è poco meno di 4 metri. Pur non essendoci differenze statisticamente significative, la tesi A (ceduo matricinato con 50 matricine ad ettaro) ha fatto registrare i maggiori incrementi in diametro e in altezza. Si può supporre l’esistenza di una correlazione negativa tra l’aduggiamento provocato dalle matricine rilasciate a dote del bosco e l’accrescimento della componente agamica del soprassuolo. Analizzando i valori di area basimetrica e di volume dei polloni, il trattamento che presenta i valori più elevati è la tesi A, dovuto sia al maggior numero di polloni qui presenti che alle loro maggiori dimensioni diametriche.
L’analisi dei dati diametrici inerenti le piante portaseme ha evidenziato un accrescimento non omogeneo tra i trattamenti. Più in particolare le matricine della tesi C (ceduo composto) hanno fatto registrare incrementi unitari maggiori rispetto alle restanti tesi, sia in termini di area basimetrica che di volume.
Concludendo, la capacità produttiva dei soprassuoli (somma dei volumi derivanti dalla crescita delle matricine e dalla componente agamica del ceduo) sembra sia strettamente correlata all’intensità delle matricine rilasciate a dote del bosco. L’intensità di matricinatura infatti influisce in senso negativo sulla produzione legnosa del ceduo.
Considerazioni da un punto di vista produttivo, ma anche biologico, porterebbero ad accettare un grado di matricinatura relativamente basso, dove le matricine svolgono essenzialmente la funzione di sostituire le ceppaie che hanno esaurito la loro capacità pollonifera.
In alternativa grande interesse potrebbe avere anche il ceduo composto, che insieme ad una elevata produzione di ghianda adatta per l’alimentazione del bestiame, offre anche una significativa produzione legnosa. Il ceduo composto infine garantisce effetti positivi anche dal punto di vista paesaggistico-ambientale. Inoltre le percentuali dei volumi appartenenti alle matricine e ai polloni segnalano come nelle aree trattate a ceduo composto vi sia un maggior equilibrio tra le due componenti rispetto alle altre tesi ceduo considerate.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar