Structural characteristics of young plantations with pedunculate oak and hornbeam in the Veneto region
Forest@ - Journal of Silviculture and Forest Ecology, Volume 6, Pages 4-18 (2009)
doi: https://doi.org/10.3832/efor0554-006
Published: Jan 29, 2009 - Copyright © 2009 SISEF
Research Articles
Abstract
The Asparago tenuifolii-Quercetum roboris [22] is the forest climax association of Po Valley. Nowadays these forests, located on a few patches in Veneto, that totally cover about 60 hectares, show serious problems in stands structure and functions. They are very important for the nature conservation even if they are fragmented in small stands. Foresters manage them as an open museum, adopting particular forestry technique to preserve and maintain the overstorey vegetation, even through new plantations near the ancient woods. Since 1988, thanks to the CAP’s strategy of set-aside, hundreds hectares of agricultural land were converted to forest in order to restore the ancient plain forest. These new forests have a lot of problems caused by errors in the projecting phase and by cultural lacks so they need a model for the management of the stands. The relationships between age and the most important tree variables, natural regeneration and herbs vegetation, were studied by a chronosequence approach over 10 forest sites. This article presents the results of the study about the diameter at breast height (henceforth: dbh) and the other tree variables relationships analyzed both at species and social class level. The authors suggested a new planting scheme aimed at promoting the natural restoration.
Keywords
Asparago tenuifolii-Quercetum roboris, Plantations, Ancient woods, Chronosequence, Tree variables, Social class
Premessa
L’Asparago tenuifolii-Quercetum roboris [22], costituisce l’associazione forestale climax della Pianura Padana. Tale formazione, peraltro oggi estremamente limitata in termini di estensione (circa 60 ha in Veneto), è dominata nel piano arboreo da Quercus robur L. e Carpinus betulus L. con Acer campestre L., Fraxinus angustifolia Vahl e Ulmus minor Mill. come specie secondarie ([12]). Solamente a partire dagli anni ’80, in cui prese avvio il movimento ecologista, si è constatato un crescente interesse del pubblico verso la tutela di questi boschi relitti. Grazie anche all’impulso di tale spinta sociale, a livello europeo sono stati definiti appositi strumenti normativi (reg. CEE n. 797/85, reg. CEE n. 2080/92, Piani di Sviluppo Rurale, ecc.) che hanno consentito di finanziare interventi di riconversione di estese superfici agricole in boschi, sia da parte degli agricoltori che di enti pubblici.
Tuttavia questi nuovi boschi, realizzati soprattutto per gli incentivi offerti o per questioni di visibilità“politica”, oltre a numerosi problemi derivanti da errori di progettazione e da carenze culturali dovute alla scarsità di esperienze pregresse e di conoscenze nel settore, soffrono anche della mancanza di modelli di gestione forestale. Per tale motivo, è stata effettuata una indagine volta a fare il punto della situazione ed a porre le basi per un monitoraggio di lungo periodo sui principali impianti realizzati nella Regione Veneto, al fine di valutare le variazioni di accrescimento a scala individuale, a livello di classe sociale e di specie. Inoltre si è voluto valutare la dinamica auxometrica e strutturale del complesso dei popolamenti studiati in funzione della mortalità naturale e dei modelli di coltivazione, dei diversi ritmi di accrescimento per classe sociale e dei caratteri bio-ecologici delle specie componenti.
Materiali e metodi
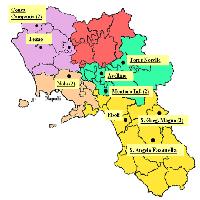
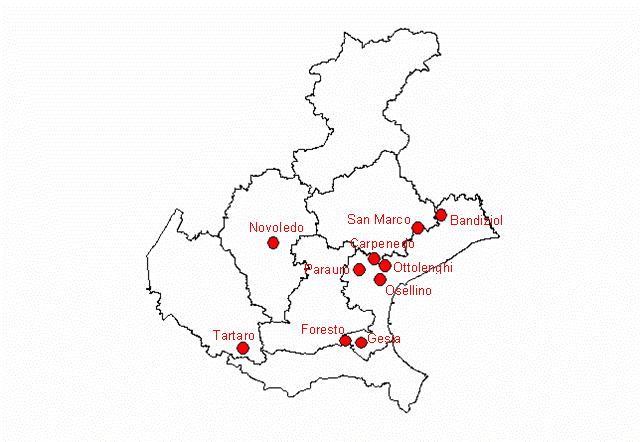
Sul territorio regionale del Veneto sono stati selezionati 10 siti (Fig. 1 e Tab. 1) nei quali sono presenti degli impianti artificiali a prevalenza di farnia e carpino bianco con età variabile tra 7 e 18 anni su una superficie complessiva di 235 ettari. Si tratta di stazioni ubicate principalmente nella provincia di Venezia e di Treviso e due stazioni, rispettivamente, in provincia di Vicenza e Verona, su terreni della bassa pianura (fa eccezione il sito di Novoledo, che si trova al confine tra alta e bassa pianura), situati generalmente ad un’altitudine di pochi metri sul livello del mare. L’area presenta una notevole omogeneità sia nelle caratteristiche climatiche che in quelle pedologiche. In particolare, i suoli che ospitano le formazioni a querco-carpineto sono generalmente profondi, con tessitura tendenzialmente fine, con drenaggio mediocre, falda profonda, regime idrico udico, a reazione alcalina, moderatamente calcarei (Regione Veneto - [2]). Negli impianti si sono individuate complessivamente 24 diverse situazioni differenziate per età e/o per composizione specifica: al loro interno sono stati rilevati 10.815 soggetti arborei ed arbustivi in 197 aree di saggio lineari (transect), costituite da porzioni di filare di 100 metri di lunghezza. Al fine di valutare le variazioni intercorse tra l’epoca di messa a dimora e quella del rilievo, in base al disegno d’impianto sono inoltre stati censiti anche i soggetti morti (o ritenuti tali) che complessivamente ammontano a 1327 unità. Per ciascun soggetto arboreo ed arbustivo, sono stati rilevati i seguenti parametri: specie, diametro a 1.3 m, altezza dendrometrica, diametro della chioma, altezza di inserzione della chioma, posizione sociale (dominante, codominante, dominata) e coordinate relative.
Fig. 1 - Localizzazione delle aree campione nel Veneto.
Tab. 1 - Caratteristiche generali degli impianti.
| Stazione | Anno impianto |
Età al rilievo | Disegno d’impianto |
Distanza intrafilare (m) | Distanza interfilare (m) | Uso di specie pronto effetto |
Uso di pacciamatura |
|---|---|---|---|---|---|---|---|
| Carpenedo (VE) | 1990-1998 1999 | 7 | sinusoidale, a gruppi | 2.0 | 4.0 | no | si |
| 8 | sinusoidale, a gruppi | 1.8 | 3.7 | no | si | ||
| 9 | sinusoidale, a gruppi | 1.7 | 3.3 | no | si | ||
| 16 | leggermente sinusoidale, a gruppi | 2.4 | 3.9 | no | si | ||
| 17 | leggermente sinusoidale, a gruppi | 2.3 | 3.9 | no | si | ||
| Foresto (VE) | 1991 | 15 | sinusoidale | 1.5 | 4.0 | no | si |
| Gesia (VE) | 1991 | 15 | sinusoidale | 1.5 | 4.0 | no | si |
| Novoledo (VI) | 1988-1991 | 10 | sinusoidale | 1.0 | 2.5 | no | plastica e vegetale |
| 18 | sinusoidale | 1.5 | 3.0 | no | vegetale | ||
| Osellino (VE) | 1994 | 9 | sinusoidale, a gruppi | 2.3 | 3.2 | si | si |
| 11 | sinusoidale, a gruppi | 2.1 | 3.2 | si | si | ||
| Ottolenghi (VE) | 1997-1999 | 7 | sinusoidale | 2.5 | 3.0 | no | si |
| 8 | sinusoidale | 2.3 | 3.1 | no | si | ||
| 9 | sinusoidale | 2.1 | 3.2 | no | si | ||
| 10 | sinusoidale | 2.5 | 3.2 | no | si | ||
| Parauro (VE) | 1992 | 12 | sinusoidale, a file alterne alberi arbusti | 1.7 | 3.1 | no | si anche sperimentale |
| San Marco (TV) | 1995-1997 | 9 | sinusoidale | 1.6 | 3.3 | no | si |
| 11 | sinusoidale | 2.0 | 2.9 | no | vegetale | ||
| Bandiziol e Prassaccon (VE) |
1995-1996 1997-1998 1999 | 7 | sinusoidale | 1.7 | 3.5 | no | si |
| 8 | sinusoidale | 1.9 | 3.2 | no | si | ||
| 9 | sinusoidale | 1.4 | 3.1 | no | si | ||
| 10 | sinusoidale | 1.9 | 3.5 | no | si | ||
| 11 | sinusoidale | 2.1 | 3.0 | no | si | ||
| Tartaro (VR) | 1991 | 14 | sinusoidale | 1.2 | 3.0 | no | si |
Procedure statistiche e di elaborazione dei dati
Al fine di definire delle relazioni generali tra il diametro a 1.3 m e le altre variabili oggetto di rilievo, per ciascuna specie e posizione sociale è stata eseguita una regressione non lineare (PROC NPAR1WAY SAS Institute Inc. Cary, NC, 2001) attraverso una procedura bootstrap sui dati grezzi da cui sono stati estratti 1000 campioni con un tasso di campionamento corrispondente al 50% della popolazione totale. Tale metodo costituisce una tecnica statistica di ricampionamento che consente di approssimare media e varianza di uno stimatore, costruire intervalli di confidenza e calcolare p-values di test quando, in particolare, non si conosca la distribuzione della statistica di interesse. I valori dei parametri di regressione sono stati quindi mediati, calcolando i rispettivi limiti di confidenza ed il valore di R2 di ciascuna funzione usata per l’interpolazione dei dati.
Per modellizzare le relazioni ipsometriche e le relazioni diametro-altezza di inserzione della chioma, sono state testate diverse funzioni matematiche (Gompertz, Chapman-Richards e logistica) ampiamente usate in campo forestale. Tra queste, è stata scelta quella di Gompertz che ha dato i migliori risultati in base al valore di R2, ai limiti di confidenza calcolati con una probabilità statistica del 95% ed alla convergenza della regressione. La formula di riferimento è (eqn. 1):
in cui y è l’altezza dendrometrica (m) oppure l’altezza di inserzione della chioma (m); dbh è il diametro a 1.3 m (cm); a, b, c sono i parametri da stimare con la regressione.
I parametri della formula vengono stimati attraverso un processo iterativo ed ogni nuova iterazione serve a minimizzare la dispersione dovuta ai residui e migliorare il valore di R2. Per lo studio della relazione diametro fusto-diametro chioma, è stata invece utilizzata una funzione logaritmica che impiega come variabili dipendenti il diametro a 1.3 m assieme all’altezza della pianta ([32], [26]). La formula di riferimento è (eqn. 2):
in cui dch è il diametro medio della chioma (m); dbh è il diametro del fusto a 1.3 m (cm); h è l’altezza dendrometrica (m); a, b, c sono i parametri da stimare con la regressione.
Risultati e discussione
Composizione specifica e numero di piante ad ettaro
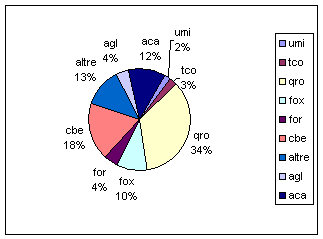
Nel dataset esaminato (Fig. 2), le specie arboree principali (farnia e carpino bianco) costituiscono circa il 52% del totale e riflettono in gran parte il risultato dei criteri e della composizione adottati in fase di progettazione degli impianti. Analoghe considerazioni si possono fare anche per le specie arbustive, fra cui prevale il biancospino, seguito dalle altre specie che sono rappresentate nella stessa misura (nocciolo, frangola, prugnolo, pallon di maggio).
Fig. 2 - Composizione del dataset specie arboree (Legenda: agl = Alnus glutinosa; aca = Acer campestre; umi = Ulmus minor; tco = Tilia cordata; qro = Quercus robur; fox = Fraxinus angustifolia; for = Fraxinus ornus; cbe = Carpinus betulus; altre = altre specie).
In 4 impianti sono stati effettuati dei diradamenti con un prelievo medio del 23.5% degli alberi; nei rimanenti 20 impianti le fallanze ammontano all’11.5% delle piante messe a dimora.
In base alla posizione sociale (Tab. 2), mediamente sono presenti circa 1005 soggetti arborei ha-1, di cui quasi tre quarti occupano i piani superiori (dominante e codominante) e i rimanenti invece rappresentano il piano dominato. Lo strato dominante è costituito soprattutto da specie eliofile dotate di buoni ritmi di accrescimento (farnia, frassino ossifillo, ontano nero, olmo campestre ed altre specie) che rappresentano circa la metà del soprassuolo arboreo. Le specie arboree accessorie (acero campestre, orniello e tiglio selvatico) sono invece fortemente rappresentate nel piano dominato mentre il carpino bianco è equamente distribuito in tutti i piani di vegetazione. Lo strato arbustivo è mediamente composto da 406 soggetti ha-1, rappresentati quasi esclusivamente nel piano dominato.
Tab. 2 - Densità media a ettaro per specie e posizione sociale dello strato arboreo.
| Specie | Pos. dominante |
Pos. codominante |
Pos. dominata |
densità ha-1 |
|---|---|---|---|---|
| Quercus robur | 201 | 78 | 76 | 355 |
| Carpinus betulus | 57 | 66 | 66 | 190 |
| Acer campestre | 23 | 40 | 60 | 123 |
| Fraxinus angustifolia | 84 | 11 | 6 | 101 |
| Fraxinus ornus | 1 | 8 | 33 | 42 |
| Alnus glutinosa | 25 | 8 | 6 | 40 |
| Tilia cordata | 6 | 4 | 16 | 26 |
| Ulmus minor | 19 | 2 | 1 | 22 |
| Altre specie | 62 | 22 | 22 | 106 |
| Totale | 480 | 240 | 285 | 1005 |
| % | 48% | 24% | 28% | 100% |
Distribuzione diametrica
La distribuzione diametrica totale degli alberi può essere suddivisa in sottodistribuzioni di ciascuna classe sociale ([21], [4], [34]). In ogni caso, Assmann ([3]), nella descrizione di una tipica struttura di un popolamento in funzione della classe sociale e del piano di vegetazione, ha notato che non è raggiungibile una completa corrispondenza tra le classi della distribuzione diametrica e le classi bioecologiche o sociali delle piante, in quanto normalmente vi è una certa sovrapposizione nei diametri tra le varie classi sociali. Questa sovrapposizione è in parte dovuta ad errori nella classificazione e in parte causata dalla relazione stocastica tra le altezze delle piante e i diametri e la variabilità nello spazio delle dimensioni delle piante nel popolamento. La curva della distribuzione diametrica per posizione sociale, oltre ad evidenziare i ritmi di accrescimento, ben si presta a indicare le esigenze ecologiche delle diverse specie arboree.
In particolare, per quanto riguarda la farnia (Fig. 3), si può notare che fin dalle prime fasi di sviluppo (diametri inferiori) vi è una forte differenziazione tra gli accrescimenti dei soggetti appartenenti ai diversi piani. La marcata eliofilia della specie e la rapidità di accrescimento consentono alla maggioranza dei soggetti di farnia di svilupparsi nel piano dominante raggiungendo un massimo di frequenza in corrispondenza di 8 cm di diametro. I soggetti del piano codominante e dominato raggiungono invece il massimo a valori di diametro rispettivamente pari a 4 cm e 2 cm, manifestando così la scarsa tolleranza della specie alla riduzione di luminosità e di spazio che tende a comprometterne, generalmente in modo irreversibile, lo sviluppo. Al contrario, l’accentuata tolleranza dell’ombra per il carpino bianco consente uno sviluppo regolare anche ai soggetti che occupano il piano codominante e dominato (Fig. 4), in quanto non si osserva una netta separazione tra i soggetti dei diversi piani né per quanto riguarda le distribuzioni di frequenza né per il diametro massimo. Simili considerazioni si possono fare anche per l’acero campestre, specie ben adattata a tollerare ridotti livelli di luminosità. Per quanto concerne le altre specie, si possono identificare i comportamenti simili del frassino ossifillo, dell’olmo campestre e dell’ontano nero, specie eliofile ed a rapido accrescimento, con un massimo della curva di distribuzione diametrica a 10 cm. Analogia di comportamento viene dimostrata anche per orniello e tiglio selvatico, che rappresentano le specie arboree caratterizzate dallo sviluppo iniziale più lento, con individui presenti quasi esclusivamente nel piano dominato.
Fig. 3 - Distribuzione diametrica della farnia per posizione sociale.
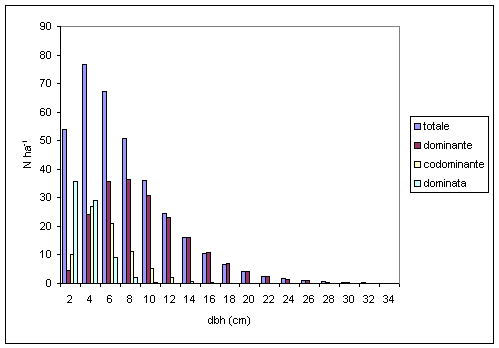
Fig. 4 - Distribuzione diametrica del carpino bianco per posizione sociale.
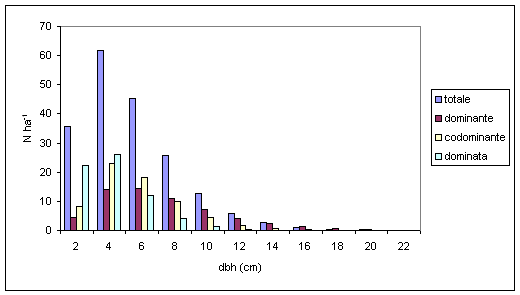
Nel confronto tra specie aventi la medesima posizione sociale si conferma l’esistenza di differenti velocità di accrescimento diametrico. Per il piano dominante (Fig. 5), farnia, frassino ossifillo, olmo campestre e ontano nero presentano un massimo centrato su valori di 8-10 cm di diametro, mentre per le altre specie arboree secondarie (acero campestre, carpino bianco) ed accessorie (tiglio selvatico ed orniello) tale valore si riduce rispettivamente a 6 cm ed a 2 cm. Per quanto riguarda il piano codominante (Fig. 6), la farnia dimostra un rallentamento dell’accrescimento che la pone allo stesso livello dell’acero campestre e del carpino bianco. Tale andamento sembrerebbe confermare la “sofferenza” dei soggetti appartenenti a queste specie soprattutto nei confronti dello spazio e dalla luce. Per quanto concerne il frassino ossifillo la scarsità degli individui presenti farebbe invece supporre che non vi sia una effettiva separazione tra il piano dominante e codominante. Nel piano dominato (Fig. 7), l’analogia di comportamento tra farnia, acero campestre e carpino bianco tende ad accentuarsi ancora di più, arrivando quasi alla sovrapposizione tra le distribuzioni diametriche. L’orniello ed il tiglio selvatico continuano a presentare comportamenti differenziati che tenderebbero a confermare uno sviluppo diametrico più “tardivo” rispetto alle altre specie arboree.
Fig. 5 - Distribuzione diametrica del piano dominante per specie (Legenda: qro = Quercus robur; cbe = Carpinus betulus; fox = Fraxinus angustifolia; aca = Acer campestre; umi = Ulmus minor; agl = Alnus glutinosa).
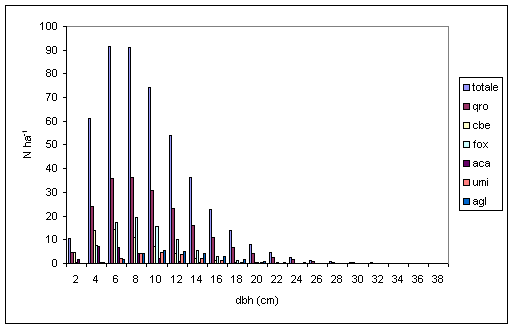
Fig. 6 - Distribuzione diametrica del piano codominante per specie (Legenda: qro = Quercus robur; cbe = Carpinus betulus; fox = Fraxinus angustifolia; aca = Acer campestre).
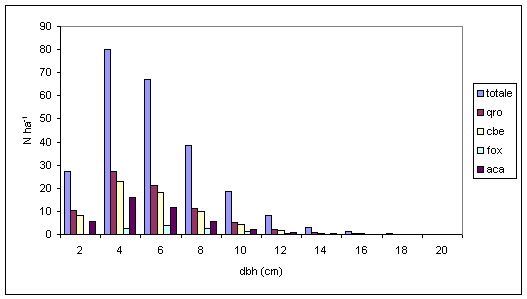
Fig. 7 - Distribuzione diametrica del piano dominato per specie (Legenda: qro = Quercus robur; cbe = Carpinus betulus; aca = Acer campestre; tco = Tilia cordata; for = Fraxinus ornus).
Area basimetrica
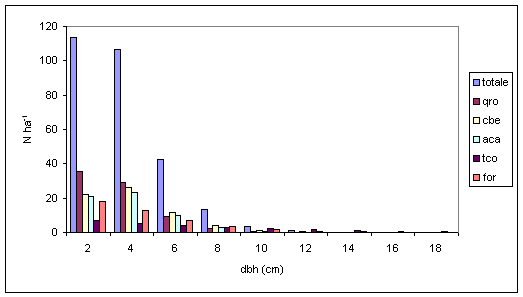
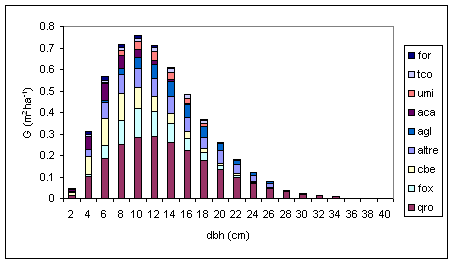
L’area basimetrica è indicativa delle relazioni esistenti tra la densità e la crescita delle piante ([4]). Inoltre, in associazione con il numero di piante, l’area basimetrica può essere usata per definire il tipo e l’intensità dei diradamenti ([15], [29]). In linea generale, la curva dell’area basimetrica è sempre caratterizzata da una distribuzione che si approssima molto a quella normale. La farnia si conferma come specie predominante (Tab. 3 e Fig. 8), oltre che in termini di numerosità degli individui, anche in termini di area basimetrica, mentre il carpino bianco, pur essendo numericamente superiore, a causa dello sviluppo più lento viene uguagliato in termini di area basimetrica dal frassino ossifillo.
Tab. 3 - Area basimetrica media per specie arborea.
| Specie | G (m2 ha-1) | G (%) |
|---|---|---|
| Quercus robur | 2.22 | 42% |
| Fraxinus angustifolia | 0.65 | 12% |
| Carpinus betulus | 0.63 | 12% |
| Altre specie | 0.61 | 11% |
| Alnus glutinosa | 0.46 | 9% |
| Acer campestre | 0.28 | 5% |
| Tilia cordata | 0.20 | 4% |
| Ulmus minor | 0.19 | 4% |
| Fraxinus ornus | 0.09 | 2% |
| Totale | 5.33 | 100% |
Fig. 8 - Distribuzione dell’area basimetrica per specie arborea (Legenda: for = Fraxinus ornus; tco = Tilia cordata; umi = Ulmus minor; aca = Acer campestre; agl = Alnus glutinosa; altre = altre specie; cbe = Carpinus betulus; fox = Fraxinus angustifolia; qro =Quercus robur).
Relazioni ipsometriche
La previsione dell’altezza totale basata sul diametro a 1.3 m della pianta è uno dei dati comunemente richiesti sia nella gestione pratica che nei lavori di ricerca in campo selvicolturale ([23]). La stima del volume delle piante e la descrizione del popolamento e del loro sviluppo nel tempo, richiedono la definizione di accurate funzioni che leghino l’altezza al diametro ([10]). Molti modelli di accrescimento richiedono come variabili di base l’impiego dei dati di diametro ed altezza, in cui le altezze di tutti o di parte dei soggetti siano derivate da valori diametrici misurati ([8]). Nel caso in cui non siano disponibili valori attuali di crescita in altezza, le funzioni ipsometriche possono essere indirettamente usate per predire la crescita in altezza ([20]).
A fini gestionali, è molto importante avere informazioni attendibili sull’altezza dei giovani popolamenti forestali in quanto la programmazione temporale di alcune operazioni selvicolturali, come i diradamenti, dipende in primo luogo dall’altezza degli impianti.
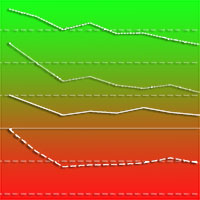
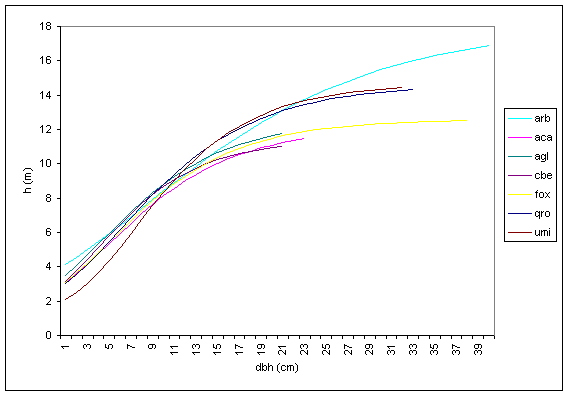
I risultati delle regressioni effettuate hanno prodotto dei valori del coefficiente di determinazione R2 compresi tra un minimo di 0.453 (carpino bianco dominato) ed un massimo di 0.805 (farnia dominante). Il valore di R2 è più elevato per le specie del piano dominante e tende a diminuire passando alle altre posizioni sociali. La curva ipsometrica del piano dominante (Fig. 9) denota un primo tratto di crescita molto simile e senza particolari differenze tra le diverse specie arboree esaminate. Questa fase iniziale dello sviluppo del bosco è caratterizzata da un accrescimento dei soggetti che dipende soprattutto dai fattori stazionali e dalle cure colturali in quanto non si è ancora innescata la competizione per lo spazio e la luce. La differenziazione tra le specie arboree considerate rispettivamente principali (farnia, olmo campestre e frassino ossifillo) ed accessorie (acero campestre, carpino bianco, ontano nero) si registra in corrispondenza di un diametro di circa 11-12 cm e ad un’altezza variabile tra 8 e 10 m, a sua volta corrispondente ad una età media compresa tra 10 e 15 anni in funzione della velocità di accrescimento. A titolo di confronto è stata ricavata anche la curva ipsometrica complessiva del piano arboreo dominante che rappresenta pertanto la media di tutte le specie arboree, con esclusione del genere Populus e di specie non completamente coerenti con la composizione del querco-carpineto (ad es., Ulmus laevis Pallas, Populus x euramericana (Dode) Guinier, Ostrya carpinifolia Scop.). Tale curva risulta fortemente influenzata soprattutto dalla farnia che costituisce la specie maggiormente rappresentata negli impianti indagati.
Fig. 9 - Curva ipsometrica per specie del piano dominante (Legenda: arb = arboree totale; aca = Acer campestre; agl = Alnus glutinosa; cbe = Carpinus betulus; fox = Fraxinus angustifolia; qro = Quercus robur; umi = Ulmus minor).
Le specie arboree principali sono anche quelle che dimostrano un più rapido accrescimento (Tab. 4) e, di conseguenza, raggiungono il diametro soglia ad una età media inferiore rispetto alle specie arboree secondarie, ad accrescimento più lento. A tale proposito si deve osservare che il frassino ossifillo, nella prima fase di crescita, non dimostra un ritmo di accrescimento così elevato come la farnia e l’olmo campestre, tanto che si sarebbe portati a classificarlo tra le specie arboree secondarie. Un altro dato interessante che conferma l’inizio della competizione è il valore del diametro medio della chioma che si attesta per tutte le specie intorno a 5 m: se si considerano i valori medi del sesto d’impianto e si confrontano con le dimensioni delle chiome, si comprende come queste ultime, dopo una prima fase di crescita libera, siano ormai parzialmente sovrapposte e compenetrate, con la conseguenza di limitazioni per quanto concerne la luce e lo spazio.
Tab. 4 - Valori medi di altezza, diametro chioma, età ed altezza inserzione chioma dei soggetti arborei per specie e posizione sociale dominante in corrispondenza di dbh cm 11-12 (n.= numerosità soggetti nel dataset con dbh 11-12 cm; h= altezza in m; dch= diametro chioma in m; hins= altezza inserzione chioma in m).
| specie | n. | età | h | dch | hins |
|---|---|---|---|---|---|
| Acer campestre | 11 | 15 | 8.0 | 5.0 | 0.8 |
| Alnus glutinosa | 41 | 15 | 8.6 | 4.0 | 4.0 |
| Arboree totale | 358 | 11 | 9.5 | 5.0 | 1.0 |
| Carpinus betulus | 23 | 14 | 8.6 | 5.0 | 0.0 |
| Fraxinus angustifolia | 52 | 10 | 10.0 | 5.0 | 1.0 |
| Quercus robur | 153 | 10 | 9.6 | 5.0 | 1.0 |
| Ulmus minor | 24 | 10 | 9.0 | 5.0 | 0.5 |
Relazioni ipsometriche generali del piano arboreo
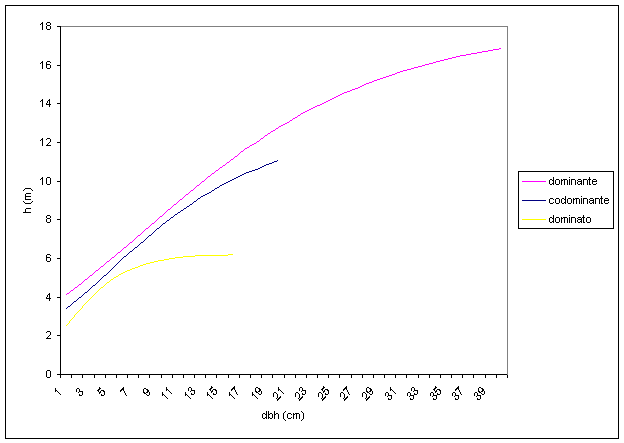
Le curve ipsometriche generali del piano arboreo (Fig. 10) hanno un andamento parallelo fino ad un diametro di 6 cm, dove inizia il rapido declino di quella riferibile al piano dominato, mentre le altre due continuano con lo stesso andamento fino a circa 13-14 cm, soglia limite a cui avviene la differenziazione. Il piano dominato sembrerebbe essere costituito da individui che presentano un accrescimento iniziale in diametro ed altezza decisamente inferiore rispetto agli altri. Tale accrescimento ridotto tende a protrarsi anche nel futuro, destinandoli o a permanere nello stesso piano, qualora siano in grado di sopportare bassi livelli di luminosità, o a soccombere alla concorrenza. Tra il piano dominante e codominante esistono generalmente solo delle differenze legate all’età media, mentre il piano dominato è nettamente differenziato in tutti i valori dei parametri arborei, dimostrando un rallentamento nella crescita. Le analisi dei valori medi del dataset (Tab. 5) riportano come in corrispondenza di un diametro di 13-14 cm sembri avvenire la differenziazione, sia in termini di età che di altezza, tra gli individui del piano codominante e dominante. La concentrazione massiccia degli individui nel piano dominante sembra peraltro indicare che la selezione naturale non stia ancora agendo in maniera incisiva, e quindi la composizione iniziale degli impianti tenda a mantenersi abbastanza inalterata nel tempo, creando la necessità di un intervento selvicolturale per consentire un armonico sviluppo del popolamento.
Fig. 10 - Curva ipsometrica generale del piano arboreo per posizione sociale.
Tab. 5 - Valori medi di altezza, diametro chioma, età ed altezza inserzione chioma dei soggetti arborei in corrispondenza di dbh 13-14 cm (n.= numerosità soggetti nel dataset con dbh 13-14 cm; h= altezza in m; dch= diametro chioma in m; hins= altezza inserzione chioma in m).
| Pos. sociale | n. | età | h | dch | hins |
|---|---|---|---|---|---|
| dominante | 222 | 11 | 10.5 | 5 | 1 |
| codominante | 9 | 14 | 9 | 5 | 2 |
| dominata | 3 | 14 | 8 | 5 | 0 |
Relazioni tra diametro ed altezza di inserzione della chioma
L’ampiezza della chioma e l’area di proiezione sul terreno ([9]) sono misure importanti per descrivere:
- l’efficienza nell’accrescimento ([5], [3], [30], [19], [13], [14]);
- la competizione a livello di popolamento ([18], [11], [31], [1], [28], [24]);
- per l’impiego in modelli di accrescimento individuale su base spaziale ([6], [7], [16], [26]).
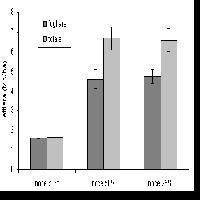
L’ampiezza della chioma è una importante misura di alcuni fattori chiave nella gestione forestale ([26]). Su base individuale, essa aiuta a descrivere la competizione esistente fra soggetti arborei ed essendo correlata alla dimensione dei rami, indirettamente esprime anche la qualità del legname prodotto ([33]), e quindi il valore economico di una pianta. Su base di popolamento, serve ad esprimere il grado di copertura delle chiome, che d’altra parte è sia un indicatore della competizione generale esistente sia un’importante misura della qualità dell’habitat forestale. In analogia con la linea di indagine precedente, nell’ambito della ricerca si è ritenuto di analizzare anche le relazioni esistenti tra il diametro del fusto e il diametro della chioma a livello di posizione sociale e specie arborea. I risultati delle regressioni effettuate hanno prodotto dei valori del coefficiente di determinazione R2 compresi tra un minimo di 0.814 (farnia e orniello dominati) ed un massimo di 0.977 (olmo campestre dominante). Anche in questo caso, il valore di R2 è più elevato per le specie del piano dominante e tende a diminuire passando alle altre posizioni sociali.
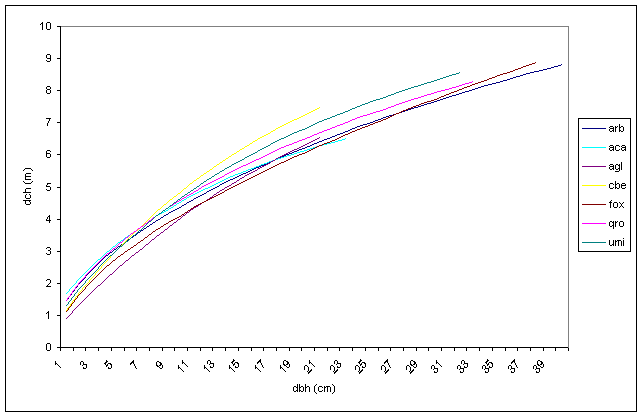
Ad un diametro del fusto di 11-12 cm, corrispondente al livello di differenziazione ipsometrico tra le specie arboree principali ed accessorie, le chiome dei soggetti dominanti raggiungono un diametro medio compreso tra 4 e 5.3 m. Se si considerano le distanze d’impianto utilizzate, al raggiungimento di questo diametro del fusto le chiome hanno ormai cominciato a compenetrarsi in modo più o meno consistente soprattutto qualora nella progettazione siano state privilegiate le specie arboree rispetto alle arbustive. In linea generale, si può affermare che con i sesti d’impianto e la composizione impiegati la copertura tende a chiudersi piuttosto precocemente e comunque entro i primi 10-12 anni. Per quanto riguarda il piano dominante (Fig. 11), è interessante notare come tutte le specie presentino un accrescimento analogo della chioma. Infatti, a parità di diametro, la massima variazione riscontrata tra le specie per l’ampiezza della chioma è di circa un metro. Le specie eliofile (farnia ed olmo campestre) sono anche quelle dotate di chioma più ampia e di accrescimenti iniziali superiori alla media. Il carpino bianco, dopo una crescita iniziale simile alle altre specie, a partire da un diametro del fusto di circa 8 cm diventa la specie che si accresce più velocemente. L’acero campestre sembra essere l’unica specie che, dopo un buon accrescimento iniziale, tende a rallentare ed a declinare lo sviluppo della chioma. Questo comportamento è probabilmente dovuto al fatto che, nella fase iniziale, la luminosità avvantaggia maggiormente le specie eliofile. Le specie tolleranti l’ombra diventano più efficienti nell’utilizzo della radiazione solare quando lo strato arboreo tende a saturare lo spazio.
Fig. 11 - Relazione diametro fusto-diametro chioma per specie del piano dominante (Legenda: arb= arboree totale; aca= Acer campestre; agl= Alnus glutinosa; cbe= Carpinus betulus; fox= Fraxinus angustifolia; qro= Quercus robur; umi= Ulmus minor).
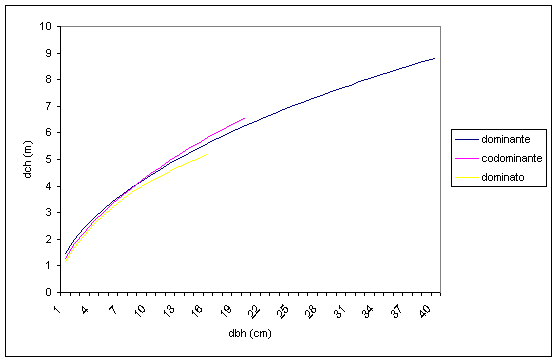
L’indagine è poi proseguita anche nell’analisi delle relazioni esistenti tra le diverse posizioni sociali all’interno di ciascuna specie arborea. Le specie che compongono lo strato arboreo tendono a sviluppare la chioma in modo complementare tra di loro, in misura quasi uguale tra i diversi piani sociali (Fig. 12). Solamente per il piano dominato, a partire da un diametro di circa 10-12 cm, si inizia ad osservare qualche segno di riduzione della chioma, probabilmente a causa del declino di qualche soggetto appartenente a specie eliofile. L’assenza di differenziazione tra i piani è confermata anche dal punto di vista dello sviluppo verticale della struttura dei popolamenti studiati, che generalmente si presenta monoplana.
Fig. 12 - Relazione diametro fusto-diametro chioma dello strato arboreo per posizione sociale.
Parallelamente allo sviluppo della copertura arborea, si assiste ad una progressiva riduzione della quantità di radiazione luminosa in grado di raggiungere le parti inferiori della chioma, che tende pertanto ad un progressivo disseccamento. La velocità del fenomeno di autopotatura dei rami è connessa all’ecologia della specie ed è direttamente proporzionale alle sue esigenze di luce.
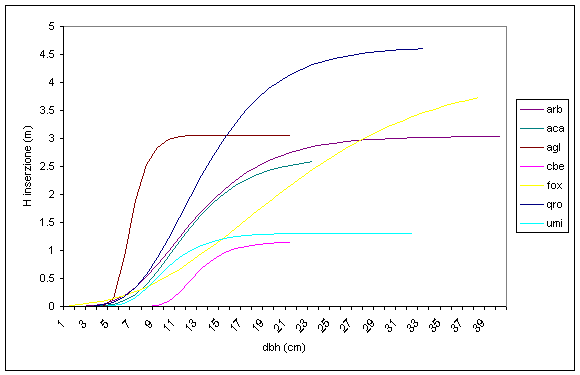
Tra le variabili indagate, l’altezza di inserzione della chioma si contraddistingue per i valori più bassi del coefficiente di determinazione R2, che variano tra un minimo di 0.086 (acero campestre dominato) ed un massimo di 0.457 (farnia dominante). Anche in questo caso, il valore di R2 è più elevato per le specie del piano dominante e tende a diminuire passando alle altre posizioni sociali. In tutte le specie si registra un andamento analogo, più o meno accentuato a seconda delle esigenze ecologiche, soprattutto nei confronti della luce. Per quanto concerne il piano dominante (Fig. 13), possiamo notare che l’inizio dei fenomeni di autopotatura si inizia a percepire (0.2-0.5 m di altezza) per quasi tutte le specie quando gli individui raggiungono un diametro del fusto di 8 cm. Fa eccezione in carpino bianco che, grazie alla sua sciafilia, mantiene i rami verdi fino a diametri superiori a 10 cm. Il disseccamento della parte bassa della chioma procede con velocità diverse: in linea generale è maggiore per le specie eliofile mentre è di minore intensità per le specie mesofile o sciafile.
Fig. 13 - Relazione diametro fusto-altezza di inserzione chioma per specie del piano dominante (Legenda: arb = arboree totale; aca = Acer campestre; agl = Alnus glutinosa; cbe = Carpinus betulus; fox = Fraxinus angustifolia; qro = Quercus robur; umi = Ulmus minor).
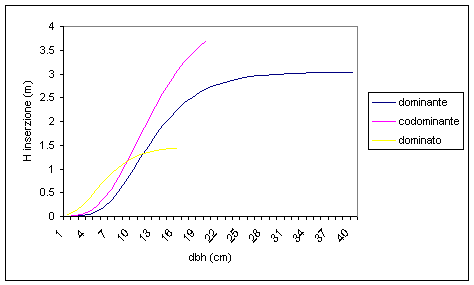
Passando all’analisi per specie in base alla posizione sociale, emerge un comportamento analogo per tutte le specie a prescindere dalle differenti esigenze ecologiche (Fig. 14): gli individui del piano dominante sono quelli che possiedono la chioma più sviluppata per estensione verticale e che risentono in misura minore dei fenomeni di autopotatura. Al contrario, gli individui del piano dominato sono quelli con la chioma più stentata e che manifestano precocemente ed in misura accentuata i fenomeni di autopotatura.
Fig. 14 - Relazione diametro fusto-altezza di inserzione chioma dello strato arboreo per posizione sociale.
Considerazioni di tipo gestionale
Per gli impianti oggetto di indagine si può osservare come, grazie agli accorgimenti agronomici ed alle cure colturali adottate, la mortalità sia estremamente contenuta. Sarebbe pertanto auspicabile riconsiderare, in sede di progettazione, sia la composizione specifica da utilizzare che la densità ed il disegno d’impianto, anche in funzione delle caratteristiche del suolo. Questo consentirebbe di ridurre da un lato, i costi sostenuti per l’impianto e dall’altro, le spese derivanti dai successivi interventi selvicolturali e di manutenzione, evitando gli errori commessi nel recente passato a causa della scarsa esperienza esistente nel settore. A tale proposito, per quanto riguarda la Regione del Veneto, si evidenzia che per l’imboschimento dei terreni agricoli - sia nelle prescrizioni adottate per l’erogazione degli incentivi della L.R. 13/2003, sia per gli aiuti recati dal Programma di Sviluppo Rurale 2007-2013 - pur considerando le caratteristiche dei suoli nella scelta delle specie, vengano ancora consigliate delle densità d’impianto estremamente elevate (da 1904 a 3690 piante ha-1) con un minimo obbligatorio di 1300 soggetti ha-1 ([27]). L’adozione di tali livelli di densità va esattamente nella direzione opposta all’obiettivo della rinaturalizzazione: infatti la rinnovazione naturale, che potrebbe insediarsi già nei primissimi anni dall’impianto, viene del tutto compromessa a causa della precoce chiusura delle chiome.
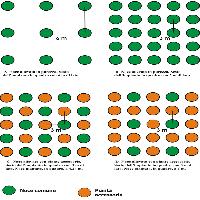
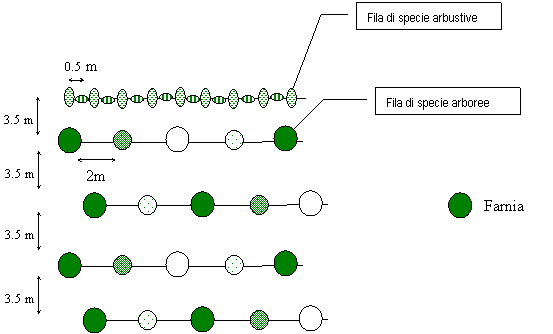
Le osservazioni in campo mostrano come le specie arboree a rapido accrescimento (pioppi, salici, farnia, olmo, frassino ossifillo), fungendo da posatoi privilegiati per l’avifauna, possano costituire dei nuclei di disseminazione soprattutto per le specie arboree ed arbustive legate alla diffusione zoocora ([17]). Per tale motivo, risultano scarsamente motivate le densità d’impianto consigliate, soprattutto nel caso delle specie arbustive, variabili tra 714 e 1667 piante ha-1. Si propone pertanto una variante al modulo d’impianto n. 1 (Fig. 15) suggerito dalla Regione Veneto ([27]). Tale modulo prevede una struttura costituita da quattro file di piante di specie arboree, alternate tra loro in maniera irregolare lungo la fila; ogni 4 file viene inserita una fila di sole specie arbustive alternate tra loro in maniera irregolare. La densità teorica è di 1964 piante ha-1 di cui 1250 di specie arboree e 714 di specie arbustive. L’impianto per file di sole specie arbustive dovrebbe consentire di creare delle siepi all’interno dell’imboschimento: gli arbusti subiranno una spontanea diffusione negli anni a venire ad opera di uccelli e polloni radicali. L’assenza di arbusti lungo le file di specie arboree giustifica la elevata densità di impianto di queste.
Fig. 15 - Modulo d’impianto n. 1 della Regione Veneto ([27]).
Pur prescindendo dalle caratteristiche pedologiche, il modulo che viene proposto come variante adotta invece una densità minore sia nelle specie arboree che arbustive. La struttura (Fig. 16) è costituita complessivamente da sei file, di cui cinque di specie arboree, alternate tra loro in maniera irregolare lungo la fila; ogni cinque file, viene inserito un filare di sole specie arbustive, alternate tra loro in maniera irregolare. La densità teorica del modulo corrisponde a 756 piante ha-1, di cui 576 ha-1 di specie arboree e 180 ha-1 di specie arbustive.
Fig. 16 - Modulo d’impianto proposto.

La distanza interfilare di 4 metri consente ai soggetti posti a dimora di svilupparsi in maniera ottimale senza che si instauri precocemente la competizione per la luce e lo spazio. In questo modo, le chiome dei soggetti arborei tenderanno a compenetrarsi verso i 12 anni. La luce che arriva al suolo sarà sufficiente a consentire lo sviluppo della rinnovazione naturale. Nelle file di specie arboree, ogni 3 soggetti potrebbe essere messo a dimora un soggetto isolato di specie a rapido accrescimento (pioppo nero e bianco, salice bianco, olmo campestre, frassino ossifillo) che, andando a costituire il posatoio preferenziale per l’avifauna, favorirà la formazione un nucleo di rinnovazione naturale in grado di svilupparsi ed affrancarsi.
L’impianto per file di sole specie arbustive con distanza intrafilare di 3 m, riducendo al minimo la concorrenza, permette uno sviluppo ottimale dei soggetti. Nel contempo, tale soluzione consente di creare dei nuclei ai margini ed all’interno dell’imboschimento dai quali si potrà avere una spontanea diffusione ad opera dell’avifauna e per riproduzione agamica per polloni radicali, accelerando, in tal modo, i processi di rinaturalizzazione del territorio.
Conclusioni
Lo studio dei giovani impianti planiziali a prevalenza di farnia e carpino bianco del Veneto, ha dimostrato l’utilità di ricorrere alla classificazione in base alla posizione sociale ed all’analisi per specie arborea ai fini dell’interpretazione ecologica delle dinamiche interspecifiche ed intraspecifiche in atto tra le diverse componenti. La differenziazione tra le diverse posizioni sociali sembra avvenire molto precocemente e ciò, soprattutto nel caso di specie eliofile (farnia, orniello), condiziona in maniera determinante lo sviluppo futuro degli individui. Inoltre la densità, il rapido sviluppo ed il precoce instaurarsi della competizione che si sono riscontrati in tutti gli impianti oggetto dell’indagine, caratterizzati da una composizione ed una struttura articolata con mescolanza di numerose specie aventi diverse esigenze ecologiche, evidenziano come sia necessario fin dai primi anni guidare l’evoluzione del soprassuolo mediante diradamenti al fine di avvicinarsi ad una composizione ottimale per una cenosi planiziale, correggendo eventuali valutazioni erronee effettuate in fase di progettazione.
I ritmi di accrescimento osservati dimostrano che, dopo 10-12 anni dall’impianto, i soggetti del piano dominante raggiungono un diametro medio del fusto di 11-12 cm ed un’altezza compresa tra 8 e 10 m, mentre lo sviluppo delle chiome determina la copertura totale del suolo. Il forte dinamismo e la notevole capacità di colonizzazione, evidenziata dalla componente arbustiva, rende sicuramente più economico, ma ugualmente valido, l’impianto degli arbusti secondo filari o gruppi ed in percentuali più ridotte di quanto effettuato finora.
È necessario infine ricordare che questi impianti sono un insieme artificiale di alberi ed arbusti, che non possono essere considerati un vero bosco, pur presentando una composizione e, dopo pochi decenni, anche una struttura articolata che si ispira a quella riscontrabile nelle foreste planiziali. Per ricostituire un bosco sono necessari tempi molto lunghi indispensabili per arricchire la biocenosi della miriade di componenti minori (funghi, batteri, invertebrati, erbe ecc) ora assenti e difficilmente introducibili in modo artificiale, ma componenti fondamentali di un ecosistema forestale ([25]).
Se l’obiettivo principale di questi impianti è la rinaturalizzazione del territorio, è necessario massimizzarne la stabilità ecologica nel minor tempo possibile promuovendo i meccanismi di rinnovazione naturale, anche attraverso l’adozione di idonei disegni d’impianto.
Ringraziamenti
Il lavoro è stato parzialmente svolto nell’ambito della convenzione tra l’Università degli Studi di Padova - Facoltà di Agraria - Dipartimento Territorio e Sistemi Agro-forestali ed il Comune di Venezia - Direzione Centrale Sviluppo del Territorio e Mobilità, Trasformazioni Urbane Progetti e Piani Ambientali, relativa allo studio degli aspetti economici ed ecologici del Bosco di Mestre. Si ringraziano il dr. Giustino Mezzalira ed il dr. Francesco Pelleri per aver messo gentilmente a disposizione i dati relativi al sito di Novoledo. Un particolare ringraziamento va agli anonimi Revisori per i preziosi suggerimenti e le indicazioni fornite che hanno consentito di apportare notevoli miglioramenti all’articolo.
References
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Online | Google Scholar
Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Online | Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar