Growth dynamics and climatic sensitivity of Silver fir (Abies alba Mill.) in the European important site (SIC) at the Alpe della Luna - Bocca Trabaria (PU - Italy)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 6, Pages 85-99 (2009)
doi: https://doi.org/10.3832/efor0564-006
Published: Mar 25, 2009 - Copyright © 2009 SISEF
Research Articles
Abstract
A dendroecological and dendroclimatic analysis was carried out in silver fir stand of Fonte Abeti, in the Apennines of central Italy. This small forest is the only one present in the northern side of the Marche region and it is currently included as a priority habitat in the Site of Community Interest “ Luna-Bocca Trabaria”. The study aimed to assess the sensitivity of silver fir to the current climate variability and was carried out on 40 dominant trees, for which we reconstructed radial growth dynamics, pointer years and climate-growth relationships. These were computed for the period 1892-2003, using monthly mean temperatures and precipitation drawn from the ALP-IMP climate dataset using simple and moving correlations in order to verify response stationarity through time.
Keywords
Abies alba, Tree-ring growth, Climate-growth response, Moving correlation function, Habitat directive, Forest management
Introduzione
Secondo i dati dell’ultimo inventario forestale ([60]) le foreste di abete bianco (Abies alba Mill.) in Italia si estendono per circa 65.000 ha con una distribuzione prevalentemente alpina. In Italia centrale, il progressivo deperimento dell’abete bianco e la contrazione del suo areale, testimoniata ancora oggi da molti toponimi nei quali la specie non è più presente, sono ampiamente documentati in letteratura a partire dai primi decenni del 1900 ([40], [84], [49], [80], [50], [25]). È vero peraltro che l’elevato valore economico che ha avuto in passato il legname di abete, ne ha determinato la diffusione per via artificiale anche in stazioni non propriamente idonee.
L’abete bianco dell’Appennino presenta alcuni caratteri ecologici differenti rispetto a quello centro-europeo, che però non sembrano giustificare l’esistenza di una varietà appenninica, come proposto da Giacobbe ([51], [52]). Dalla seconda metà del secolo scorso sono stati intrapresi numerosi studi relativi a possibili differenze genetiche ([82], [81], [41], [68], [58]), morfologiche ed ecologiche ([82], [97], [74], [54], [72], [30], [53], [71], [67], [83]) tra provenienze ed al suo deperimento diffuso in Italia centrale ([75]) e soprattutto in Toscana ([31], [32], [64], [59], [65]).
Le analisi genetiche e polliniche raggruppano per somiglianza l’abete appenninico a quello del centro Europa, ipotizzando l’esistenza di una rotta migratoria post-glaciale dall’Appennino centro-settentrionale verso nord e l’isolamento delle popolazioni calabresi nel rifugio glaciale ([66], [14], [62], [81], [24], [99], [101], [69]). Queste ultime sono caratterizzate da un’elevata variabilità genetica che ha permesso l’evoluzione di ecotipi particolarmente adattatati alle condizioni ambientali del luogo. Le altre popolazioni invece sono andate incontro a fenomeni di deriva genetica e di perdita della variabilità come probabile risposta a condizioni climatiche avverse incontrate durante il processo di ricolonizzazione ([66], [14], [24], [41], [102]). Tale riduzione è considerata un fattore predisponente a forme di stress che hanno condotto a fenomeni di deperimento e, nei casi estremi, alla locale scomparsa dell’abete bianco, registrati dalla metà del XX secolo in gran parte dell’Europa centrale ([66]). In Italia il deperimento è stato messo in relazione principalmente ad annate siccitose consecutive soprattutto in popolamenti con criticità strutturali e funzionali ed ha contribuito a diminuire il valore economico del legname in Italia e all’estero ([84], [22], [37], [63], [75]). La ricerca sull’abete bianco in Italia ha subito un generale rallentamento e soltanto recentemente la necessità di conservare e valorizzare l’abete appenninico degli habitat prioritari della rete Natura 2000, ha rinnovato in parte l’interesse per questa specie ([16], [18], [17], [48]). Sono stati realizzati a tale proposito alcuni progetti LIFE per la salvaguardia e la reintroduzione dell’abete ([1], [2], [4]). Altri studi hanno indagato con approccio dendroecologico le popolazioni di maggiore importanza; Abetone e Campigna ([47]), Camaldoli e La Verna ([73]), Vallombrosa ([36]), Pescopennataro ([91], [93]), Rosello, Serra San Bruno ([92]) e Basilicata ([48]). Tali studi hanno messo in evidenza i trend di accrescimento dell’abete bianco, le loro relazioni con gli andamenti climatici e gli eventuali disturbi causati da interventi antropici. Molto poco si conosce ancora sui nuclei minori distribuiti fra le Marche e l’Abruzzo nei versanti orientali dell’Appennino, che potrebbero fungere da cerniera tra le estese abetine molisane e quelle tosco-emiliane.
In questo lavoro vengono presentati i risultati relativi ad uno dei siti (abetina di Fonte Abeti, PU) presi in esame nell’ambito di una ricerca in corso in Romagna, Marche, Abruzzo e Molise che ha come obiettivi: a) la caratterizzazione strutturale-compositiva delle cenosi di abete; b) la loro sensitività alla variabilità climatica in atto ed alla selvicoltura applicata; c) la definizione di indirizzi gestionali per la loro conservazione e/o valorizzazione. Per Fonte Abeti le analisi finora svolte avevano come obiettivi: 1) l’analisi dendrocronologica, al fine di evidenziare il trend di accrescimento e ipotizzare i fattori che hanno concorso a determinarlo, nonché di confrontarlo con quello di altre cronologie appenniniche omospecifiche; 2) l’analisi dendroclimatica, che permette di osservare le relazioni causali fra accrescimento radiale dell’abete e principali fattori climatici (precipitazioni e temperature medie) e di verificarne la stabilità nell’ultimo secolo.
Materiali e metodi
L’area di studio
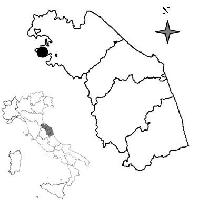
L’abetina di abete bianco oggetto dello studio si trova in località Fonte Abeti in comune di Borgo Pace (PU), nei pressi del passo di Bocca Trabaria, nell’Appennino tosco-umbro-marchigiano e costituisce uno dei nuclei di Abies alba più settentrionali delle Marche. (Fig. 1). La cenosi occupa una vallecola sul ripido versante Nord del Poggio del Romito, si estende per 15 ha interamente nel SIC “Alpe della Luna-Bocca Trabaria” (IT5310010) ed è compresa negli habitat prioritari “Faggeti degli Appennini con Taxus e Ilex” (9210) e “Faggeti degli Appennini con Abies alba e Faggeti con Abies nebrodensis” (9220).
Fig. 1 - Localizzazione dell’area di studio.
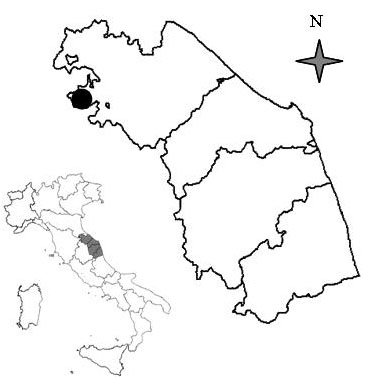
Secondo la classificazione bioclimatica di Rivas-Martinez, l’area appartiene al macrobioclima temperato a variante sub-mediterranea con termotipo mesotemperato superiore e ombrotipo umido inferiore. Il regime udometrico è subequinoziale-autunnale caratterizzato dalla presenza di un brevissimo periodo di aridità estiva tra la metà di luglio e la metà di agosto.
Il substrato geo-litologico è di tipo pelitico-calcarenitico in sequenze torbiditiche, il quale dà origine a pendii con pendenza superiore anche al 60%. I suoli sono subacidi, profondi (25-75 cm) e sciolti ([3]).
L’abetina ha un nucleo in purezza di probabile origine artificiale a 900-1000 m slm, che sfuma a quote superiori in una faggeta mesofila submontana in conversione a fustaia e, a quote inferiori, in una cerreta sub-mesofila submontana.
Sono state realizzate due aree di saggio circolari con raggio di 20 m, una a 960 m slm nel cuore dell’abetina (FAB1) ed una a circa 1050 m slm, dove la composizione è più eterogenea ed aumenta la componente di latifoglie (FAB2). Nel primo caso il soprassuolo ha struttura biplana con un piano dominante rappresentato dalla fustaia matura coetanea di abete bianco ed un piano dominato, poco consistente, costituito da acero a foglie ottuse (Acer obtusatum W. et K.), nocciolo (Corylus avellana L.) e faggio (Fagus sylvatica L.) di prevalente origine agamica. In FAB2 aumentano le latifoglie nel piano dominante e gli individui di abete sono di maggiori dimensioni. In entrambi i casi la vigoria del popolamento è elevata e la copertura è superiore all’80%, (di poco maggiore in FAB2). Semenzali e rinnovazione sono assenti o danneggiati per l’elevata densità di copertura e la brucatura da parte del capriolo ([55], [33]).
Raccolta ed elaborazione dei dati dendrometrici
Di ciascun individuo arboreo censito avente diametro superiore a 7.5 cm ad 1.30 m da terra, sono stati misurati il diametro a 1.30 m e l’altezza totale; inoltre per l’abete bianco sono state estratte anche due carote legnose (una a monte e l’altra a 90° da questa) con trivella di Pressler. Sono state prelevate altre carote da individui prossimi alle due aree per raggiungere un numero adeguato di campioni ai fini dell’analisi dendrocronologica. In totale sono state utilizzate per le analisi successive 85 carote da 44 individui (da 3 alberi è stato possibile prelevare solo un campione di qualità sufficiente). Le carote, montate su appositi supporti e levigate con carta abrasiva a grana via via più fina (80-400 grits), sono state sottoposte a misurazione utilizzando il sistema ottico semi-automatico LINTAB (Rinntech, Germany) costituito da un binoculare con crocefilo ed un piano mobile i cui spostamenti, corrispondenti alle ampiezze anulari espresse in 1/100 mm, sono stati registrati dal software collegato TSAPWin 0.55 (Rinntech, Germany). Con lo stesso software sono state eseguite le progressive sincronizzazioni tra serie elementari, per giungere alla costruzione delle cronologie individuali e medie di ogni area di saggio. La sincronizzazione è stata effettuata con metodo visivo e statistico mediante il CDI (Cross Date Index), un indice di sincronizzazione calcolato da TSAPWin e derivante dalla combinazione di un test non parametrico Glk (Gleichlaeufigkeit) ([42]) e del t di Student ([87]), ambedue ampiamente utilizzati per valutare la concordanza di andamento tra serie di ampiezze anulari. La sincronizzazione è considerata accettabile con CDI>10 ([87]).
Le serie individuali delle due aree di saggio sono state mediate per ottenere un’unica cronologia stazionale di Fonte Abeti (FAB), la quale è stata confrontata con altre cronologie di abete bianco appenninico provenienti dalle stazioni di: Camaldoli, La Verna e Poppi (AR), Sasso Fratino (FO), Abetone (PT), Cortino e Ceppo Martese (TE), messe a disposizione dai rispettivi autori.
La standardizzazione delle serie, finalizzata all’analisi dendroclimatica, è stata effettuata utilizzando il software ARSTAN ([34], [35]). Tale procedura rimuove la varianza a bassa frequenza determinata solitamente da perturbazioni non-climatiche (es. trattamenti selvicolturali) e produce serie indicizzate con valore medio pari a 1 e varianza omogenea per l’intero arco temporale della serie. Il metodo adottato consta di tre fasi: a) applicazione alle serie elementari grezze di una funzione esponenziale negativa che elimina il trend negativo dovuto alla crescente circonferenza dell’albero; b) calcolo di uno spline con risposta di frequenza del 50% su una finestra temporale di 10 anni; c) calcolo della cronologia media utilizzando una media robusta allo scopo di ridurre gli outliers, cioè i valori estremi positivi e negativi ([34], [28], [9]).
Raccolta dati climatici
Nello studio delle relazioni clima-accrescimento sono state utilizzate serie plurisecolari delle precipitazioni totali mensili (1813-2003) e degli scarti della temperatura media mensile dalla media del periodo (1774-2003) ottenute dal dataset ALP-IMP ([7], [8], [5], [6]). Questo raccoglie dati recenti e storici di stazioni meteorologiche presenti nell’area compresa tra 4° e 19° Est e tra 43° e 49° Nord omogeneizzati e spazializzati su una griglia con maglie di 1° x 1°. Gran parte della provincia di Pesaro-Urbino ricade al limite meridionale dell’area e per la stazione in esame sono stati ritenuti rappresentativi i dati del punto con coordinate 44°12°. L’impiego di queste serie garantisce maggiore significatività nelle relazioni clima-accrescimento per la maggiore completezza e continuità e soprattutto permette di valutare meglio la variabilità temporale delle risposte ottenute.
Relazioni clima-accrescimento
Anni caratteristici
Gli anelli caratteristici (pointer values) sono anelli particolarmente larghi o stretti, visibili nelle serie come picchi positivi o negativi. Questi sono stati individuati con il software Weiser ([56]) utilizzando una finestra temporale di 7 anni e tre differenti classi di intensità crescente. Gli anni in cui almeno il 75% dei campioni presenta un anello caratteristico, sono stati considerati anni caratteristici (pointer year) ([10], [94], [76], [78]). Si è quindi proceduto alla ricerca di una corrispondenza tra eventi climatici estremi ed anni caratteristici dopo aver messo in evidenza fenomeni climaticamente rilevanti attraverso il calcolo dei percentili (3°, 5°, 10°, 90°, 95°, 97°) nel dataset ALP-IMP. Secondo la definizione dell’IPCC un evento climatico è considerato estremo se supera il 10° o il 90° percentile ([61]). A questo scopo è stato possibile analizzare solo il periodo 1915-2006 poiché nel periodo precedente la cronologia è formata da meno di dieci campioni, soglia minima per considerare significativo un pointer year ([10]).
Correlazioni lineari
Le risposte incrementali dell’abete bianco ai fattori climatici sono state valutate con il calcolo della correlazione tra serie di accrescimento indicizzate e serie termo-pluviometriche relative al punto scelto della griglia ALP-IMP (P e Scarti Tmed). A tale scopo è stato utilizzato il software DENDROCLIM2002 ([21]) che, con metodo bootstrap (1000 replicazioni), restituisce i coefficienti di correlazione e le funzioni di risposta per ogni variabile climatica inserita. Non essendoci differenze significative nei profili ottenuti per le correlazioni e per le funzioni di risposta e dal momento che le correlazioni sono ritenute più conservative ([27]), più stabili e facilmente replicabili delle funzioni di risposta ([23], [79], [85]), si è scelto di considerare soltanto i valori delle correlazioni.
Poiché l’accrescimento annuale dell’abete bianco può essere condizionato da eventi climatici verificatisi anche prima dell’inizio del periodo vegetativo ([11], [89]), si è considerato un anno biologico a partire dal mese di maggio dell’anno precedente la formazione dell’anello (n-1) ad ottobre dell’anno di formazione (n) ([43], [44]). Per ciascuna variabile climatica sono stati calcolati i valori di correlazione medi per l’intero periodo di analisi (1892-2003).
Correlazioni mobili
L’andamento nel tempo delle risposte ai fattori climatici è stato valutato ancora con il software DENDROCLIM2002, utilizzando l’opzione delle moving correlation a scansione annuale con una finestra mobile di 57 anni, determinata dalla limitata lunghezza della cronologia media disponibile (116 anni). Ad ogni iterazione e ad ogni passaggio viene calcolato un coefficiente di correlazione, la cui media finale viene determinata con il metodo bootstrap ([20]). I coefficienti di correlazione prodotti hanno una soglia di significatività diversa per ogni variabile considerata, per cui sono stati standardizzati dividendoli per la loro deviazione standard (r/s) ottenendo così una soglia di significatività uguale a ± 1.96 cui corrisponde una probabilità pari a 0.05. Nel caso specifico l’analisi dei risultati si realizza fondamentalmente con le eventuali differenze di andamento all’interno dei periodi 1891-1947 e 1948-2003.
Risultati e discussione
La dinamica di accrescimento
Le due aree di saggio, sebbene con alcune differenze di composizione e struttura, sono caratterizzate dalla prevalente presenza di abete bianco, i cui caratteri dendrometrici (Tab. 1) sono molto simili ([55], [33]). Le cronologie di abete bianco delle due aree di saggio sono state costruite utilizzando 40 serie elementari relative a 21 alberi per FAB1 e da 37 serie elementari relative a 19 alberi per FAB2. Età massima (116 anni) e media sono maggiori in FAB1, ma non ci sono sostanziali differenze negli indicatori statistici (Tab. 2), ad eccezione dei valori di ampiezza anulare media (M), che risultano maggiori di circa il 17% in FAB1 rispetto a FAB2.
Tab. 1 - Principali parametri dendrometrici dell’abete bianco nelle due aree di saggio (FAB1 e FAB2).
| Parametro | FAB1 | FAB2 |
|---|---|---|
| N/ha | 184 | 96 |
| dg [cm] | 50.7 | 65.2 |
| d dom [cm] | 58.5 | 58 |
| G [m2/ha] | 36.9 | 31.9 |
| Hg [m] | 28.3 | 31.6 |
| H dom [m] | 30.2 | 28.9 |
| V [m³/ha] | 488.5 | 413.4 |
Tab. 2 - Principali parametri statistici delle cronologie relative alle due aree di saggio (FAB1 e FAB2) e alla cronologia stazionale (FAB). M: ampiezza anulare media; DS: deviazione standard; AC(1): autocorrelazione di primo ordine; SM: sensitività media.
| Area | N. campioni |
Periodo (anni) |
M (mm) |
DS (mm) |
AC(1) | SM |
|---|---|---|---|---|---|---|
| FAB1 | 40 | 1891-2006 (116) |
3.31 | 1.34 | 0.70 | 0.218 |
| FAB2 | 37 | 1919-2006 (88) |
2.77 | 1.49 | 0.82 | 0.221 |
| FAB | 77 | 1891-2006 (116) |
3.09 | 1.42 | 0.76 | 0.219 |
In Fig. 2 sono riportate le cronologie non indicizzate delle due aree di saggio e quella media stazionale (composta da 77 serie individuali), di cui si riportano anche i valori indicizzati (Fig. 3): la sincronizzazione tra FAB1 e FAB2 è molto elevata (CDI= 81) e gli accrescimenti annuali sono molto simili fino al 1970 circa, quando, in corrispondenza della prolungata riduzione di accrescimento, FAB1 riesce a mantenere accrescimenti più elevati.
Fig. 2 - Cronologie grezze di Abies alba relative alle due aree di saggio (FAB1 e FAB2) e all’intero popolamento (FAB). L’asse di destra rappresenta la numerosità dei campioni (N).
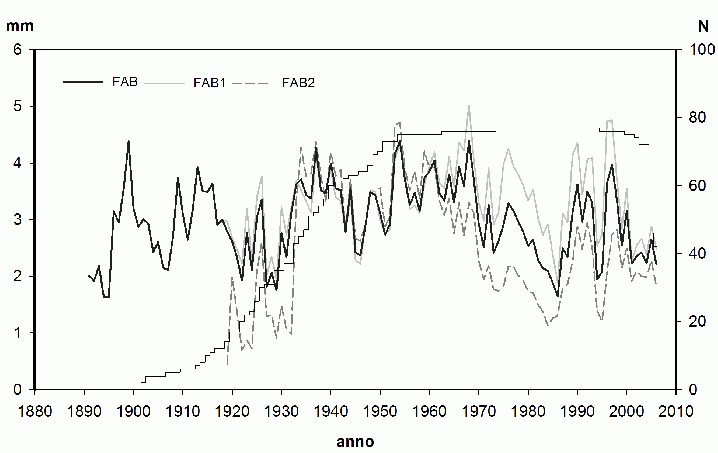
Fig. 3 - Cronologia media indicizzata di Fonte Abeti.
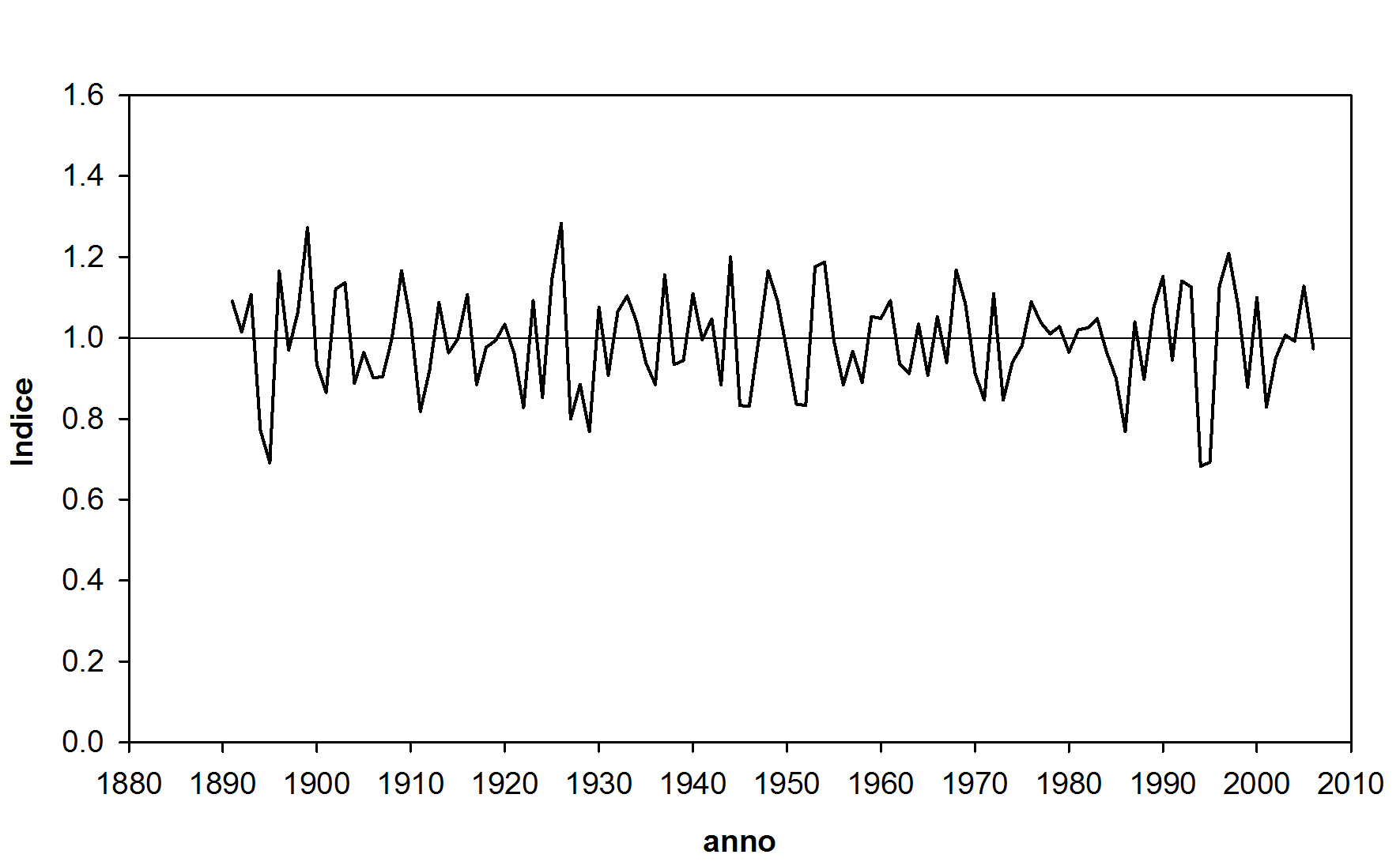
La curva media è relativamente stazionaria e caratterizzata nel primo tratto da fluttuazioni cicliche fino al 1945, che potrebbero essere attribuibili a tagli intercalari diventati più sporadici e di minore entità dopo la seconda guerra mondiale ([55]). Il periodo 1930-1970 è caratterizzato da valori di ampiezze anulari relativamente elevati, anche per la giovane età della maggior parte degli individui che compongono la cronologia in quell’intervallo temporale (Fig. 2). Dalla fine degli anni ’60 si osserva una progressiva riduzione di accrescimento che culmina negli anni 1985 e 1986; dopo una rapida ripresa alla fine degli anni ’80, l’andamento è caratterizzato da rilevanti oscillazioni e poi si assesta su valori inferiori alla media. Per valutare la natura della riduzione nel quindicennio 1970-1985, ne è stata verificata la presenza anche in altre cronologie di abete bianco appenninico. In Tab. 3 sono riportati i principali parametri statistici delle cronologie stazionali analizzate ed i rispettivi CDI con la curva media di Fonte Abeti.
Tab. 3 - Principali parametri statistici delle cronologie non standardizzate di abete bianco appenninico utilizzate per il confronto con la cronologia media di Fonte Abeti. CDI: Cross Date Index; M: ampiezza anulare media; DS: deviazione standard; AC(1): autocorrelazione di primo ordine; SM: sensitività media.
| Cronologia | CDI | Periodo (anni) |
M (mm) |
DS (mm) |
AC(1) | SM |
|---|---|---|---|---|---|---|
| Poppi (AR) | 34 | 1862-1998 (137) |
3.20 | 1.69 | 0.96 | 0.14 |
| Camaldoli (AR) | 28 | 1899-1996 (98) |
7.01 | 3.41 | 0.93 | 0.12 |
| Abeti Soprani (IS) | 23 | 1824-2006 (183) |
2.01 | 0.91 | 0.80 | 0.20 |
| Cortino (TE) | 17 | 1875-1999 (125) |
2.69 | 1.23 | 0.94 | 0.15 |
| La Verna (AR) | 16 | 1848-1996 (149) |
5.48 | 2.72 | 0.96 | 0.13 |
| Sasso Fratino (FC) | 12 | 1789-1998 (210) |
2.06 | 0.71 | 0.90 | 0.13 |
| Ceppo (TE) | 9 | 1595-1998 (404) |
0.85 | 0.44 | 0.93 | 0.16 |
| Abetone (PT) | 4 | 1845-1998 (154) |
2.09 | 0.68 | 0.82 | 0.15 |
I valori di CDI sono superiori alla soglia considerata significativa (10) per tutte le stazioni ad eccezione di Ceppo e dell’Abetone; quelli più elevati si registrano a Poppi e Camaldoli, ma la sincronizzazione visiva è buona anche per le altre stazioni. Nonostante la diversità dei caratteri stazionali, dell’estensione delle serie e della storia dei popolamenti, si evidenziano alcuni andamenti molto simili fra le curve. Un fattore di eterogeneità nella dinamica di accrescimento è individuabile nelle perturbazioni di carattere locale, come gli interventi selvicolturali, eseguiti nel secolo scorso in molte stazioni almeno fino agli anni ’60 e che producono una maggiore sincronizzazione fra le serie a partire dagli anni ’70, da quando verosimilmente in gran parte delle stazioni si sono ridotti o non si sono più effettuati significativi interventi colturali. Le cronologie di Camaldoli e La Verna ([73]) per esempio sono caratterizzate da andamenti diversi fino al 1970 che poi tendono ad assimilarsi nel periodo di crisi e di ripresa dell’ultimo ventennio.
La repentina e forte riduzione di accrescimento del periodo 1970-1986, presente in gran parte delle cronologie analizzate, è stata registrata in altre stazioni di abete bianco sia italiane che europee, non disponibili per questo lavoro. L’abete bianco piemontese nel periodo 1968-1989 presenta le più forti riduzioni di accrescimento dell’intera cronologia (1890-1990) ([86]). Anche nei Vosgi (Francia) la riduzione inizia nel 1973 e, dopo aver raggiunto il massimo nel 1976, tende a recuperare tra il 1983 e il 1987 ([11], [12]). Il fenomeno è stato associato a fenomeni di deperimento che hanno interessato l’abete bianco nel XX secolo e la cui origine climatica sembra essere avvalorata anche da ulteriori riduzioni repentine di accrescimento in corrispondenza di altri fenomeni di siccità prolungata nel corso del 1900 in Slovenia ([19]), nei Vosgi francesi ([13]), a Vallombrosa ([96], [37], [36]) ed in Casentino ([96]). Sia nei popolamenti italiani ([36], [86]) sia in quelli francesi sopra citati, il deperimento dell’abete e la riduzione di accrescimento sarebbero stati determinati (nei casi meno gravi) da severi e ripetuti fenomeni di deficit idrico. Questi avrebbero interessato maggiormente quei popolamenti già in condizioni critiche per densità eccessiva, struttura coetanea, monospecificità o per maturità eccessiva, che hanno reso l’abete più sensibile a fattori secondari di stress ([37], [57], [64], [12], [59], [45], [86], [15]). È stata osservata una minore incidenza del fenomeno in popolamenti disetanei e più equilibrati strutturalmente (La Verna e Sasso Fratino), come indicato anche da altri autori ([11], [86]). È quindi probabile che anche a Fonte Abeti le scarse precipitazioni nel periodo 1965-1977, verificatesi dopo la relativa siccità del decennio precedente (1946-56 circa) e la struttura non ottimale delle cenosi abbiano determinato una crisi incrementale dell’abete bianco. È importante sottolineare che ad Abeti Soprani (Molise) e a Serra San Bruno (Calabria) l’abete bianco non ha registrato tale riduzione, in linea con l’ipotesi che le popolazioni più meridionali sarebbero maggiormente adattate a condizioni più xeriche ([92], [91]).
L’accrescimento, dopo una rapida ripresa post-crisi, sembra essersi stabilizzato, anche se gran parte delle cronologie disponibili terminano proprio in questi anni. Nelle poche serie più lunghe non si osservano riduzioni riferibili a fenomeni di siccità recenti (es. 2003) ed è stato anche ipotizzato, per l’abete di Camaldoli e La Verna, che l’aumento delle temperature ed il conseguente allungamento della stagione vegetativa possano aver influito positivamente sull’accrescimento ([73]).
Le relazioni clima-accrescimento
Anni caratteristici
In Tab. 4 sono riportati gli anni caratteristici per l’abete di Fonte Abeti; alcuni coincidono con anni o intervalli caratteristici segnalati da altri autori per la stessa specie; la maggiore concordanza si ha per i pointer year negativi più forti (1922, 1929, 1956 e 1986), condivisi dalla maggior parte delle cronologie italiane e dell’Europa centrale. Al contrario i picchi positivi di accrescimento tipici di Fonte Abeti non trovano corrispondenza in altre stazioni.
Tab. 4 - Anni caratteristici individuati nella cronologia stazionale di Fonte Abeti; (a): se presenti nell’80% delle serie individuali, (b) nel 75%. A fianco di ogni anno sono citati i lavori in cui l’anno è considerato caratteristico.
| Pointer year negativi | Pointer year positivi | ||
|---|---|---|---|
| 1922a | [47], [86], [13], [89], [39], [90], [77] | 1926a | [47] |
| 1929a | [47], [89], [39], [77] | 1940b | - |
| 1945a | - | 1944b | - |
| 1946a | - | 1953a | - |
| 1951a | [95] | 1954a | - |
| 1952b | - | 1968b | - |
| 1956b | [73], [78], [89], [39], [77] | 1997a | - |
| 1986b | [47], [73], [78], [89], [39], [90] | - | - |
| 1994 | - | - | - |
| 1995 | - | - | - |
L’origine climatica delle riduzioni di accrescimento è riconducibile a due fenomeni diversi; per gli anni 1929, 1956 e 1986 sembra preponderante l’effetto delle temperature particolarmente basse registrate nei mesi invernali precedenti la stagione vegetativa (da gennaio ad aprile), in quanto l’abete bianco necessita di inverni abbastanza miti ([98], [15]). Nel caso invece del 1922, dei bienni 1945-46 e 1994-1995, l’azione combinata delle alte temperature primaverili ed estive e delle scarse precipitazioni, sia dell’anno che dell’anno precedente, ha influenzato negativamente la formazione dell’anello. Analogamente, all’origine delle annate positive si trovano estati fresche e piovose, talvolta accompagnate da inverni miti (come nel 1968 e nel 1997), con un effetto favorevole sull’accrescimento.
Quelle proposte sono ipotesi derivanti da un accurato confronto tra accrescimento ed eventi climatici estremi: è chiaro che, non trattandosi di un popolamento di treeline ed essendo stato soggetto ad interventi selvicolturali sporadici fino ad un trentennio fa, è possibile la presenza di altri fattori di disturbo nelle serie di accrescimento. La natura climatica della maggior parte dei pointer year è però sostenuta sia dal confronto con altre cronologie che dalla concordanza con l’andamento dei fattori climatici che sono maggiormente limitanti per l’abete bianco.
Correlazioni lineari
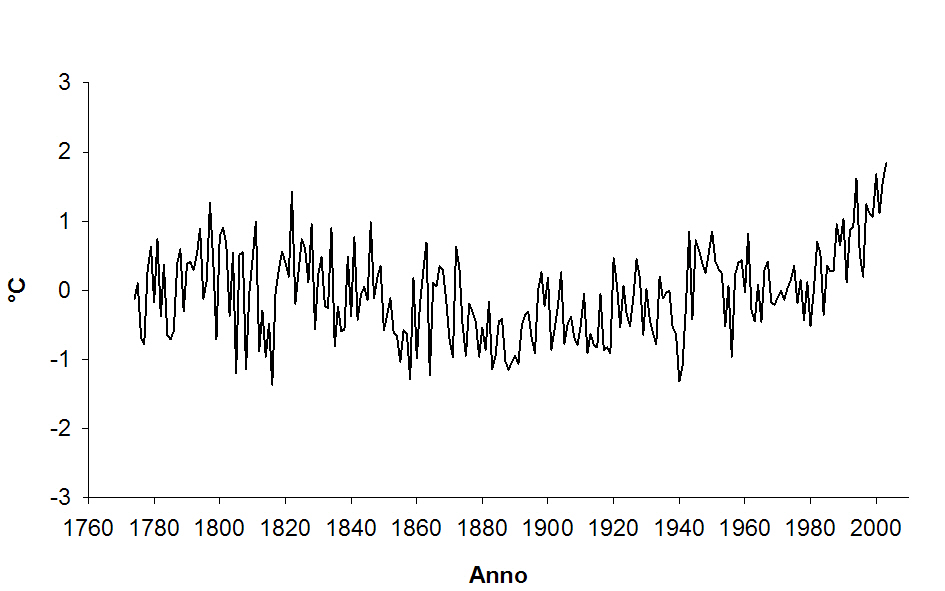
Se l’aumento delle temperature a livello planetario negli ultimi decenni è ormai una condizione verificata (IPCC, 2007), più incerti e variabili sono gli effetti che questo fenomeno produce sulla vegetazione forestale e la sua dinamica auxologica. Sia a scala globale che locale si osserva il notevole aumento delle temperature negli ultimi 25 anni (Fig. 4), culminato nel 2003, in cui la temperatura media dei mesi estivi (giugno-luglio-agosto) ha superato di 4.6°C la media degli ultimi due secoli ([5]). Minore instabilità caratterizza l’andamento delle precipitazioni, i cui valori oscillano intorno alla media, sebbene si osservi una diminuzione nell’ultimo ventennio, fenomeno che potrebbe rientrare anche in normali fluttuazioni cicliche ([5]).
Fig. 4 - Andamento delle temperature medie annue per il periodo 1774-2003 espresso come scostamenti dalla media relativa al periodo 1901-2000 per il punto 44°12° della griglia APL-IMP.
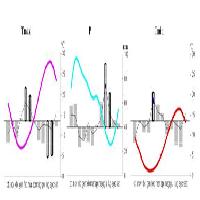
I profili delle correlazioni (Fig. 5) ottenute utilizzando i dati della griglia ALP-IMP evidenziano che i fattori che limitano maggiormente l’accrescimento dell’abete bianco sono le temperature medie estive, in particolare quelle di luglio e agosto e secondariamente quelle di giugno. Ciò significa che l’accrescimento dell’abete bianco diminuisce con l’aumento delle temperature medie estive.
Fig. 5 - Profilo delle correlazioni tra accrescimento annuale e temperatura media mensile e precipitazioni mensili. I valori maggiori di |1.96| sono significativi per P<0.05.
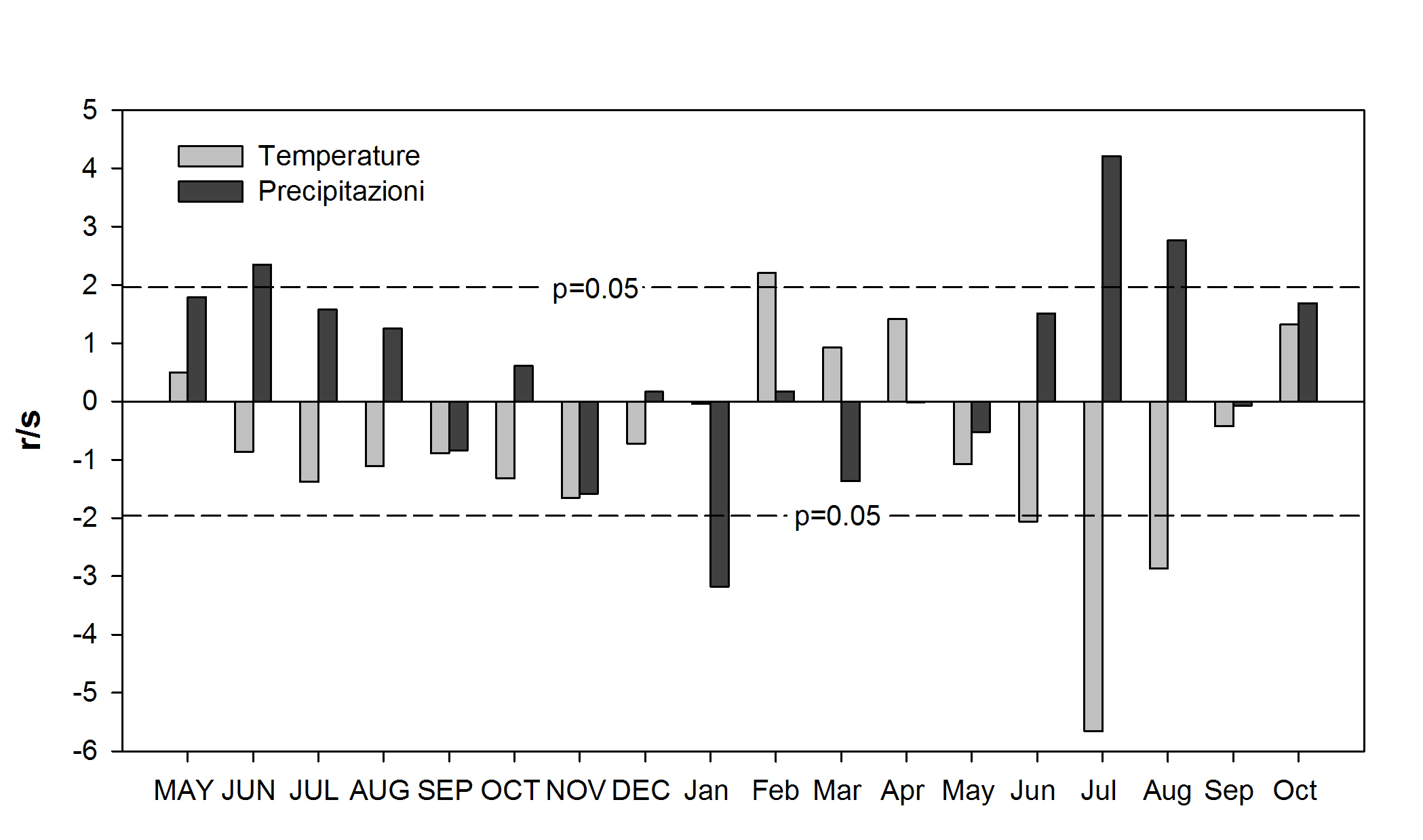
Relativamente alle precipitazioni, l’aumento di piovosità in luglio e agosto ha invece effetti incrementali positivi. La dinamica di accrescimento dell’abete di Fonte Abeti sembra quindi essere controllata prevalentemente da un effetto sinergico delle temperature e delle precipitazioni: mesi estivi molto caldi attenuano l’azione positiva determinata dalle precipitazioni primaverili ed estive analogamente a quanto osservato in altre stazioni dell’Appennino centrale ([36], [91], [47], [48]); in stazioni più piovose, come Camaldoli e La Verna, l’accrescimento è controllato in modo significativo soltanto dalle temperature di agosto ([73]). Il fattore udometrico diventa ancor più limitante per l’abete dell’Appennino centro-meridionale: infatti nelle abetine molisane la correlazione positiva è significativa solo con le precipitazioni di giugno e agosto, ma non con quelle di luglio, quando lo stress idrico è maggiore a causa delle più elevate temperature ([91]). Tale fenomeno si manifesta ancor più chiaramente nelle provenienze calabresi, dove la formazione dell’anello avviene principalmente a maggio e giugno, perché in luglio ed agosto la specie ha probabilmente evoluto un sistema adattativo di parziale chiusura degli stomi per ridurre la traspirazione ([92]). È interessante notare che la sensitività udometrica dell’abete bianco è presente anche in ambiente alpino, soprattutto nel settore occidentale, dove, nonostante le differenze nel regime delle precipitazioni e nella xilogenesi, sussistono dei periodi con rischio di stress idrico ([88], [38], [39], [89]).
Sebbene meno evidenti, sono risultate significative, in senso positivo, le temperature medie di febbraio. L’abete bianco appenninico sembra essere avvantaggiato da inverni non troppo freddi, sia nelle stazioni più settentrionali ([47], [73]) sia in quelle meridionali ([92], [91], [48]), evitando così il rischio di danni da gelo, importante fattore di stress per questa specie. Peraltro un aumento delle precipitazioni di gennaio, di probabile carattere nevoso, può determinare fenomeni di idromorfismo, con sofferenza a livello radicale e riduzione della sintesi di sostanze di riserva utili al momento della ripresa dell’attività vegetativa. Nella stazione in esame, caratterizzata anche da un’esposizione NE, questa possibilità potrebbe determinarsi in concomitanza con una prolungata permanenza del manto nevoso. In generale l’esigenza di inverni miti e asciutti è stata riscontrata per tutte le stazioni appenniniche sottoposte ad analisi dendroclimatiche ([92], [91], [47], [73]).
Anche l’abete bianco, come altre conifere, è sensibile all’influenza climatica dell’anno precedente la formazione dell’anello (n-1) ed in particolare dalle precipitazioni di giugno, correlate positivamente. Tale sensitività potrebbe essere messa in relazione alla possibilità di formare sostanze di riserva per l’anno successivo, anche se tale fenomeno avviene soprattutto nella fase conclusiva della stagione vegetativa (da agosto ad ottobre), quando la moltiplicazione cellulare è pressoché terminata, la lignificazione è in atto e quindi i fotosintetati prodotti possono essere immagazzinati, come riportato sia per le stazioni molisane ([91]) e calabresi ([92]), sia per quelle tosco-emiliane ([36], [47]) che in quelle delle Alpi francesi ([88], [38], [89]) e dei Pirenei ([100], [70]).
In sintesi si può affermare che l’abete bianco di Fonte Abeti ha un accrescimento che è sensibile soprattutto all’andamento climatico estivo, sia dell’anno di formazione dell’anello sia di quello precedente. È una specie che teme condizioni ricorrenti di elevate temperature e siccità estive, soprattutto a quote non elevate ([38], [47]). Beneficia di inverni relativamente miti, ma teme fenomeni anche locali di idromorfismo, che possono determinare l’insorgere di attacchi patogeni, di prevalente natura fungina. Le risposte climatiche lo collocano più vicino alle abetine dell’Appennino centro-settentrionale piuttosto che a quelle meridionali, dove l’abete può aver messo in atto, soprattutto al limite dell’areale (Calabria), adattamenti xerofitici ([82], [98], [54], [45], [92]) che hanno indotto alcuni autori ad ipotizzare la differenziazione di una vera e propria varietà ([51], [52]).
Correlazioni mobili
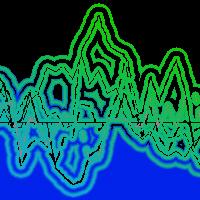
Il calcolo delle moving correlation consente di osservare meglio nel tempo l’andamento e la significatività delle singole variabili risultate fondamentali per la formazione dell’anello (Fig. 6). Relativamente alle temperature, si osserva che i dinamismi significativi (tratti della curva all’esterno della zona tratteggiata) sono limitati ai mesi estivi dell’anno di formazione dell’anello (giugno, luglio ed agosto), peraltro con alcune differenze sostanziali. Le risposte negative di giugno sono caratterizzate da un’elevata significatività iniziale, che tende a ridursi progressivamente fino a perdersi nel tratto finale della curva. Quella di luglio rimane sempre significativa, anche se nel secondo tratto subisce anch’essa una forte riduzione che tende a stabilizzarsi su valori più vicini alla soglia minima. Nel mese di agosto le risposte inizialmente non significative lo diventano progressivamente per poi invertire nuovamente l’andamento nell’ultimo periodo verso valori nuovamente vicini alla soglia minima. Ancora più interessante appare l’andamento delle precipitazioni estive, caratterizzato da una progressiva perdita e riduzione di significatività (nel settore positivo) rispettivamente dei mesi di giugno e luglio. In agosto la risposta è quasi stabilmente sopra il livello di significatività, con una lieve diminuzione nell’ultimissimo tratto della curva. Anche le precipitazioni di ottobre presentano un’interessante dinamismo caratterizzato da una relazione positiva progressivamente crescente nel tempo e che diventa significativa nel secondo intervallo temporale.
Fig. 6 - Andamento dell’effetto delle temperature medie mensili (A) e delle precipitazioni (B) sull’accrescimento dell’abete bianco a Fonte Abeti durante il periodo 1774-2003; correlazioni calcolate usando una finestra mobile di 56 anni. Le linee tratteggiate indicano la soglia di significatività pari a |1.96| per P<0.05.
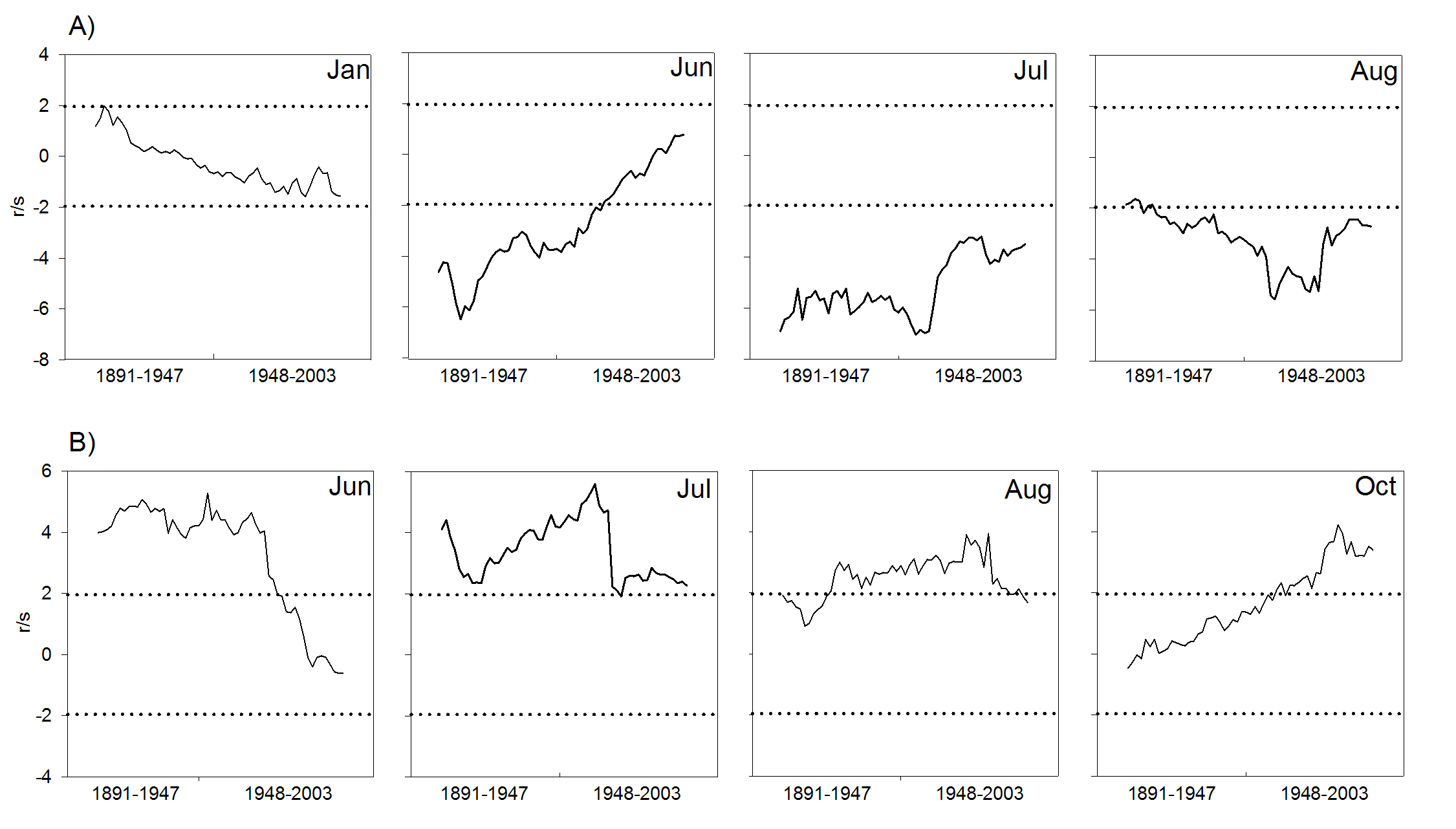
Nonostante tale metodo non consenta un controllo annuale dei dinamismi di risposta, è evidente che la sensitività al regime termo-pluviometrico estivo dell’abete bianco di Fonte Abeti si è modificata nel corso dell’ultimo secolo con un interessamento anche dei mesi autunnali. Tutte le variazioni osservate sono collocabili nel secondo cinquantennio di osservazione, evidenziando la possibilità di un rapido cambiamento di comportamento messo in atto dall’abete. La diminuzione e/o perdita di significatività delle temperature e delle precipitazioni in corrispondenza dei mesi cruciali per la formazione dell’anello farebbe ipotizzare una sorta di adattamento a condizioni più xero-termiche dell’abete, in risposta al riscaldamento climatico in atto a scala locale e globale. Se tale ipotesi è in linea con l’andamento della temperatura media dei mesi estivi, in evidente aumento anche nella zona d’indagine, molto meno lo è con quello della quantità di precipitazioni, che non evidenzia trend altrettanto evidenti.
Analoghe risposte non stazionarie delle variabili termo-pluviometriche maggiormente limitanti o incentivanti l’accrescimento radiale sono state ottenute su abete bianco nei Pirenei spagnoli ([100], [70]) e in Italia al limite superiore del bosco alpino su larice ([29], [46]) e pino cembro ([46], [26]); in tutti i casi gli autori ipotizzano possibili adattamenti alle mutevoli condizioni climatiche.
Conclusioni
I risultati dell’analisi dendrocronologica e dendroclimatica nell’abetina di Fonte Abeti (PU) sono un primo step di una ricerca in corso sull’abete bianco in Romagna, Marche, Abruzzo e Molise volta a valutarne la capacità di resilienza ai cambiamenti del clima ed ai regimi selvicolturali. Sebbene il soprassuolo indagato non sia interamente di origine naturale e sia stato sottoposto a interventi colturali irregolari, è presente nelle serie di accrescimento radiale un evidente segnale climatico che si esplicita con una buona sincronizzazione delle curve individuali ed una discreta teleconnessione ad altre cronologie italiane di abete bianco.
La cronologia media secolare (FAB) è per il momento l’unica disponibile per questa specie nelle Marche (un’altra è in costruzione ai Monti della Laga) e aggiunge ulteriori informazioni a quelle raccolte con il progetto FORMAT (ENV4-CT97-0641 - Forest Modelling Assessment from Tree Rings). Elemento caratteristico presente anche in altre cronologie italiane ed europee, è una fluttuazione negativa che inizia intorno al 1970 e che si risolve dopo il 1986. Tale evento è stato associato a condizioni perduranti di aridità che in alcuni casi avrebbero determinato situazioni di deperimento, talvolta molto accentuato, dell’abete bianco. Da confronti con altre cronologie sembra evidenziarsi una maggiore incidenza della crisi incrementale nelle formazioni pure e coetaniformi.
L’ipotesi avanzata per l’abete di Camaldoli e La Verna ([73]), secondo la quale la successiva ripresa sarebbe dovuta all’aumento delle temperature in corso da circa 30 anni, sembrerebbe poco probabile a Fonte Abeti, se si considerano le correlazioni inverse con le temperature estive. Peraltro, le moving correlation hanno evidenziato la non stazionarietà di tale comportamento ed anzi il tendenziale adattamento dell’abete bianco a condizioni maggiormente xerotermiche. Anche la concentrazione di anni caratteristici dal 1986 è minore rispetto al periodo precedente, aspetto che potrebbe avvalorare l’ipotesi precedente, se si considera che gran parte di quelli di segno negativo sembrano associati a periodi estivi caldi e siccitosi.
L’abete di Fonte Abeti manifesta un comportamento, sia nella dinamica di accrescimento sia nelle funzioni di risposta, assimilabile a quello dell’abete del Casentino, anche se la perdita di stazionarietà delle correlazioni climatiche sembrerebbe predisporre l’abete ad un comportamento più simile a quello delle abetine meridionali, molto più tolleranti ed adattate a condizioni più xerotermiche.
I risultati di questa analisi, oltre a colmare alcuni gap nel settore orientale dell’areale in Appennino centrale, forniscono indicazioni sulla potenzialità ecologica dell’abete nell’area d’indagine, utili anche per calibrare gli interventi di conservazione e valorizzazione che sono richiesti dalla direttiva Habitat. L’abetina di Fonte Abeti non è ad immediato rischio di scomparsa, ma l’eccessiva omogeneità strutturale di alcuni settori, la contiguità con nuclei di abete rosso e l’assenza di nuclei di pre-rinnovazione ([55]) potrebbero progressivamente minarne la stabilità e ridurre la resilienza che tale specie sembra dimostrare almeno in termini climatici. L’evoluzione naturale, opzione gestionale spesso adottata in aree protette o della rete Natura2000, non sembra quindi la risposta idonea ad un obiettivo di durevolezza e di sostenibilità della cenosi.
Note
Lavoro presentato al VI Congresso nazionale SISEF: “La gestione delle foreste tra cambiamenti globali e azioni locali”, Arezzo, settembre 2007.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar