Natural dynamics of holm oak-dominated forest stands in the National Park of Maremma, Tuscany
Forest@ - Journal of Silviculture and Forest Ecology, Volume 6, Pages 186-198 (2009)
doi: https://doi.org/10.3832/efor0580-006
Published: May 19, 2009 - Copyright © 2009 SISEF
Research Articles
Abstract
Mediterranean sclerophyllous forests are coenoses worthy of great care because of their inherent bio-ecological value. The suspension of harvesting following the crisis of fuel wood market ruled the evolutive pattern of maquis over the last decades. In this phase, site quality seems to be the major driver of the holm oak-dominated forest recovery as well as of the lasting permanence of the sclerophyllous shrub forest. In this connection, stand dynamics has been monitored in three plots located in the strictly protected area of the “Parco Regionale della Maremma” (Grosseto - Tuscany), where natural evolution is in progress since at least five decades. The monitored stands are representative of different evolutive phases from the holm oak-dominated coppices to the typical maquis, as a function of the original stand structure and composition but also of the intensity and length of the applied coppice management system. The analysis was carried out by permanent transects; two inventories were undertaken in 1991 and 2006 to evaluate both compositional and structural variability and the dynamics in progress. Results at now highlight the role played by the residual site quality and specific composition in driving the evolutive pattern of tree species community. Under the absence of any silvicultural practice and better site conditions, the progress and specific incidence of regular mortality and the related aboveground biomass redistribution, supported the vegetation of holm oak and the other subsidiary species typical of more mature stages. The overall forest dynamics seemed anyway to proceed slowly and resulted in a steady physiognomy over a long time since the suspension of the historical disturbance made of repeated harvesting on short rotations.
Keywords
Mediterranean macchia, Stand structure, Natural dynamics, Mortality, Growth
Introduzione
Le formazioni forestali di sclerofille mediterranee, cedui di leccio (Quercus ilex L.) e di arbusti sempreverdi- sono caratterizzate da alcuni elementi che contraddistinguono la specificità di tali soprassuoli ([18], [15], [19], [50], [1], [46], [16], [24]). In primo luogo, nonostante l’intenso impatto antropico (utilizzazioni frequenti, pascolo, incendi) protrattosi per secoli, rappresentano cenosi con un elevato grado di biodiversità e complessità della struttura arborea; in secondo luogo hanno sviluppato meccanismi adattativi (caratteri morfologici, fisiologici e fenologici) idonei a compensare gli effetti dello stress determinato dalle particolari condizioni ecologiche tipiche dell’ambiente mediterraneo.
Si tratta, inoltre, di soprassuoli che offrono attualmente limitate o nulle potenzialità di reddito pur essendo caratterizzati da un elevato valore naturalistico, storico, estetico-paesaggistico e anche protettivo nei riguardi del suolo. Infine, queste formazioni sono la più importante componente del paesaggio vegetale nelle zone litoranee e collinari del Mediterraneo. In Italia il 7% della superficie forestale nazionale è rappresentato da boschi di leccio e macchia mediterranea mentre tale percentuale sale al 16% (175 000 ha su 1 086 016 ha) in Toscana ([43]).
Le formazioni di sclerofille sempreverdi vengono interpretate come il risultato di un processo di degradazione di natura antropogenica delle originarie foreste di latifoglie decidue che, dall’età del ferro, si è protratto nel corso degli ultimi 2.000-3.000 anni ([38], [7], [42], [11]). L’elevato grado di antropizzazione ha raggiunto l’apice tra la fine dell’Ottocento e gli inizi del Novecento quando la quasi totalità delle leccete residue è stata convertita a ceduo e sottoposta a sfruttamento intensivo per l’esercizio del pascolo e per la produzione di legna da ardere e di carbone ([49], [29], [6]). Tali forme di uso, insieme al ripetuto verificarsi degli incendi, hanno innescato un processo di degradazione che, nelle fasi più avanzate, ha ulteriormente portato alla scomparsa della lecceta e all’insediamento delle formazioni a macchia, con arbusti sclerofilli dotati di elevata capacità di propagazione vegetativa e quindi in grado di rinnovarsi in modo efficace in seguito a eventi di disturbo.
Negli ultimi decenni l’interruzione della ceduazione, causata dalle difficili vicende del mercato dei combustibili vegetali, ha indirizzato diversamente il processo di evoluzione naturale della macchia mediterranea. In Toscana le utilizzazioni della macchia si sono quasi annullate tra gli anni Cinquanta e l’inizio degli anni Sessanta anche se in alcune zone, in relazione alla presenza di soddisfacente viabilità, sono state riprese verso la metà degli anni Settanta. In generale, nelle aree preservate dalle utilizzazioni si è verificato, in tempi più o meno lunghi in funzione della fertilità stazionale e delle caratteristiche microclimatiche, una lenta e graduale riduzione della partecipazione degli arbusti sclerofilli e un aumento del leccio, fino alla formazione di popolamenti quasi puri di quest’ultima specie. In definitiva si è manifestata una spontanea riduzione della complessità strutturale e della diversità delle specie arboree ed arbustive presenti ([40], [37], [26], [27], [41], [28], [14], [7], [30], [2], [23], [3], [42], [9]).
I boschi della Maremma hanno manifestato un decorso storico coerente con quanto fin qui illustrato; alla metà dell’Ottocento si presentavano molto degradati oltre che per le utilizzazioni eccessive a cui erano stati sottoposti anche per l’improvvisa importanza che avevano assunto sia per la fabbricazione della potassa sia per l’uso della corteccia di leccio nella concia delle pelli ([44]). Niccolini ([36]) descrive che ancora nel 1881 il tratto di pianura lungo la via Aurelia era coperto da una magnifica foresta di querce e di olmi secolari, che si estendeva dai forteti dell’Uccellina a quelli di Montiano.
Se, in termini generali, si può parlare di un processo di ricostituzione della lecceta nelle stazioni dove le condizioni ambientali sono più favorevoli, si percepisce anche una permanenza degli arbusteti di macchia nelle stazioni più sfavorevoli. Queste sono comunque osservazioni generiche prive di un riscontro quantitativo in termini di composizione specifica, densità, altezza e riferibili ad un dato periodo. Per questo motivo abbiamo ritenuto opportuno stabilire nel Parco Regionale della Maremma (d’ora in poi P.R.M.) aree di monitoraggio in tratti di macchia che, da oltre 50 anni, non erano state sottoposte a tagli di utilizzazione. Le informazioni attualmente disponibili inerenti l’evoluzione strutturale di tali formazioni sono infatti piuttosto limitate e interessano per lo più leccete mature o popolamenti vegetanti su substrati di origine vulcanica ([32], [48], [31]). In particolare, in questo contributo si forniscono dati quantitativi sui processi dinamici in atto nelle formazioni forestali a prevalenza di leccio presenti nel P.R.M. attraverso l’analisi della mortalità, dell’accrescimento e della variazione nel tempo del peso funzionale di ciascuna specie.
Ambiente di studio
Il Parco occupa un’ampia fascia del litorale grossetano e si estende per circa 21 km con una superficie complessiva di 9.009 ha (altri 7.993 ha rientrano nell’Area contigua al Parco) in cui ricadono i Monti dell’Uccellina e la parte della pianura grossetana solcata dall’ultimo tratto del fiume Ombrone. Sotto l’aspetto amministrativo il Parco rientra nei comuni di Grosseto, Magliano in Toscana e Orbetello; la Regione Toscana possiede il 44% dell’intera superficie boscata, il 46% appartiene a quattro grandi proprietari, mentre il rimanente 10% è frazionato tra numerosi piccoli proprietari.
I Monti dell’Uccellina sono disposti con il crinale principale parallelo alla linea di costa e raggiungono la quota massima in corrispondenza della cima di Poggio Lecci (417 m s.l.m.).
Il calcare massiccio che forma i maggiori rilievi della zona è un tipo litologico compatto; i suoli da esso derivati hanno subito un lungo processo di alterazione e di decarbonatazione e, in caso di erosione, si rigenerano molto lentamente; la vegetazione risente di questo processo di degradazione. Anche nelle zone in cui i suoli hanno avuto origine da substrati silicei attribuiti al Verrucano, le facies di degradazione della foresta di sclerofille sempreverdi, quali la macchia e la gariga, possono perdurare per tempi notevoli perché, anche se la capacità di ritenuta idrica e la profondità sono elevate, risulta insufficiente l’apporto di materiale organico e di elementi minerali atti a migliorare la fertilità chimica del suolo ([45]).
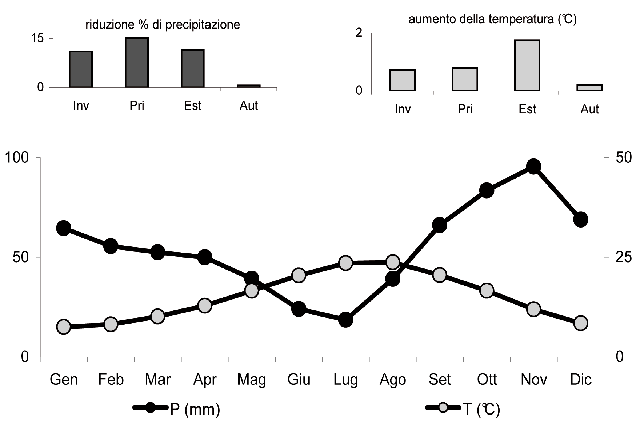
Il clima, contrassegnato da un’estrema variabilità, è di tipo mediterraneo con estati calde e asciutte e inverni miti e relativamente piovosi (Fig. 1). Il regime pluviometrico (657 mm in 62 giorni piovosi) è caratterizzato da un picco principale in autunno e da uno secondario in inverno, la stagione meno piovosa è l’estate con un prolungato periodo di aridità che va da giugno a metà settembre. La temperatura media annua è di 15.0°C, quella del mese più freddo (gennaio) di 7.5° C e quella del mese più caldo (agosto) di 23.7°C. Nel corso degli ultimi 15 anni le precipitazioni si sono ridotte in media del 10-15% mentre è aumentata la temperatura media, soprattutto in estate.
Fig. 1 - Diagramma termopluviometrico relativo alla stazione di Grosseto (lat. 42°45’ lon. 11°7’) per il periodo 1960-2000. Nei grafici in alto sono riportati la riduzione percentuale di precipitazione e l’aumento della temperatura media degli ultimi 15 anni. Dati da: Banca Dati Agrometeorologica Nazionale (Ucea).
Arrigoni et al. ([4]), in riferimento alla classificazione di Thornthwaite, collocano la stazione di Alberese “fra i climi mesotermici al limite tra il clima sub-umido-secco e quello semi-arido, nell’area cioè dei consorzi di sclerofille sempreverdi del Quercetum ilicis con possibilità di infiltrazione di latifoglie decidue nei terreni con maggior capacità idrica”.
Secondo la classificazione fitoclimatica del Pavari il territorio del Parco rientra nella sottozona media del Lauretum, secondo tipo, con siccità estiva.
Secondo l’inquadramento proposto da Mondino e Bernetti ([43]) i popolamenti oggetto di studio si inseriscono fra l’Orno-lecceta con roverella delle zone interne e la Macchia media mesomediterranea. Il primo tipo si caratterizza, dal punto di vista fisionomico e fitosociologico, per costituire chiazze di bosco sempreverde di varia statura e fertilità, che nei fondovalle vede prevalere la componente di latifoglie decidue, in prevalenza cerro e roverella, che sono state rilasciate come matricine e che determinano la fisionomia del bosco. Il secondo tipo comprende quelle formazioni di sclerofille sempreverdi mediterranee “che non fanno prevedere prospettive di sviluppo oltre i 7 m di altezza” ([25], [35]).
La storia del territorio
Le principali vicende storiche che hanno influito sulla copertura forestale sono state ricostruite sia sulla base di osservazioni dirette effettuate in diverse zone del Parco sia attraverso l’esame di carte topografiche, documenti e testimonianze storiche ([10], [39]).
La presenza del monastero di San Rabano, la segnalazione di alcuni insediamenti medievali di cui ora non rimane traccia, la vicinanza della Via romana Aurelia e la presenza di varie torri di guardia distribuite lungo la costa fanno supporre una non trascurabile presenza di coltivi e di boschi a pascolo (ne sono un ricordo le rare querce ceduate a capitozza rinvenute in zone diverse del Parco) laddove ora è presente la foresta. Ancor più significativi sono i luoghi in cui nella macchia sono presenti ulivi, chiara testimonianza di una diversa forma di uso del suolo praticata in passato.
La disciplina delle utilizzazioni legnose è documentata solo a partire dal 1934-36, anni in cui si può considerare terminata la bonifica di Alberese, in un piano dei tagli rimasto in vigore fino al 1975, anno di istituzione del Parco Naturale della Maremma. La ceduazione era effettuata ogni 15-20 anni, era previsto il rilascio di 70-120 matricine ad ettaro di specie diverse, di 3-4 turni e il pascolo con bovini maremmani era ammesso 5 anni dopo il taglio ([17], [5]). Annualmente veniva utilizzata una superficie variabile da 53 a 183 ettari (in media 102 ha).
Le ceduazioni furono effettuate regolarmente fino al 1942 - anno in cui si interruppero per cause belliche, ripresero poi nel 1946 ma cessarono progressivamente nelle varie aziende tra la fine degli anni Cinquanta e l’inizio degli anni Sessanta.
Il “Parco Naturale della Maremma”, dal 1994 “Parco Regionale della Maremma”, fu istituito nel 1975, allo scopo di “tutelare le caratteristiche naturali, ambientali e storiche della Maremma, in funzione dell’uso sociale di tali valori, nonché di promuovere la ricerca scientifica e la didattica naturalistica” (Legge 5 giugno 1975, n° 65 della Regione Toscana). L’attività forestale all’interno del Parco è pianificata dal 1980 considerando tre principali forme di uso del suolo: (i) aree a ceduo per la produzione di legna da ardere, (ii) aree a protezione integrale per scopi di ricerca e conservazione e (iii) aree in conversione all’alto fusto per esaltare le qualità estetico - ricreative e per il pascolo ([8]).
Recentemente, in ottemperanza alla legge quadro sulle aree protette (6 dicembre 1991 n° 394), è stato realizzato il nuovo Piano per il Parco ([51]), la cui zonizzazione prevede quattro aree differenziate per finalità gestionale: (i) riserve integrali; (ii) riserve orientate, in cui le attività antropiche sono finalizzate al mantenimento e alla conservazione dei sistemi; (iii) aree di protezione dove le tradizionali attività di uso del suolo sono consentite e regolamentate da piani di utilizzazione specifici; (iv) aree di promozione, finalizzate al miglioramento della vita socio-culturale delle popolazioni locali. Le superfici forestali sono comprese per la quasi totalità (96%) nella categoria aree di protezione mentre mancano completamente in quelle di promozione.
La documentazione sugli incendi è scarsa ma sembra che l’ultimo grosso incendio si sia verificato nel 1912 e che abbia interessato, oltre che tratti di pineta, anche buona parte dei monti dell’Uccellina ([37]).
Tenuto conto della sensibile pressione antropica che ha gravato per secoli su queste formazioni, è impossibile definire un processo evolutivo, progressivo o regressivo, di tipo lineare; si possono invece individuare alcuni fenomeni di disturbo della vegetazione e del suolo, di origine direttamente o indirettamente antropica, che hanno agito sulla dinamica della vegetazione con conseguenze di lungo periodo sulla composizione specifica, sulla struttura del soprassuolo e sulla fertilità stazionale.
Materiali e metodi
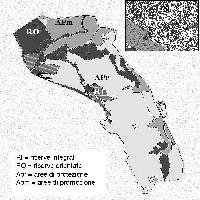
Nel 1991 furono delimitate, all’interno delle zone allora designate a riserva integrale, 3 aree di monitoraggio di 1000 m2 ciascuna, rappresentative di formazioni cedue di leccio e macchia mediterranea non sottoposte ad utilizzazioni da almeno 50 anni. L’ultima redazione del Piano del Parco ([51]) ha scorporato dalla categoria riserva integrale la zona dove ricadevano due delle tre aree permanenti (Fig. 2). Attualmente, mentre l’area denominata Poggio Lecci risulta ancora all’interno delle riserve integrali, quelle di Albatraia e Cupellaio rientrano invece nella categoria aree di protezione, “dove le tradizionali attività di uso del suolo sono consentite e regolamentate da piani di utilizzazione specifici”, situazione non confacente con le finalità della ricerca.
Fig. 2 - Zonizzazione del Parco Regionale della Maremma secondo l’ultimo piano di gestione e ubicazione delle aree permanenti.
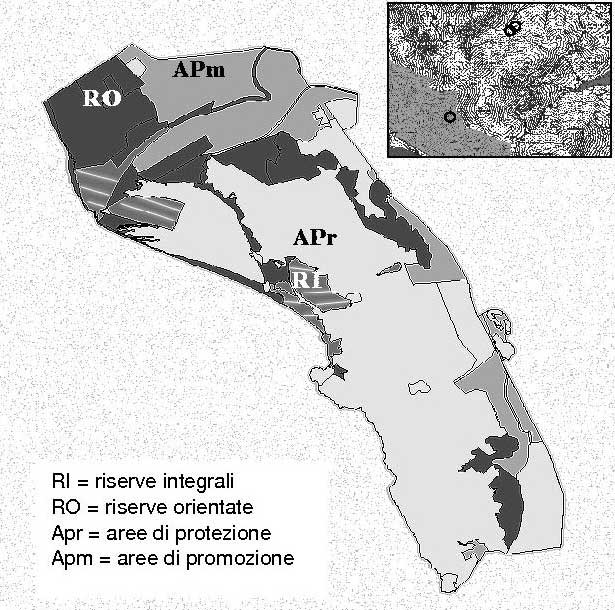
Le principali caratteristiche stazionali delle tre aree, tutte situate sul versante orientale dai monti dell’Uccellina e quindi al riparo dei venti marini, sono riportate in Tab. 1.
Tab. 1 - Principali caratteristiche stazionali ed età delle tre aree di monitoraggio.
| Parametro | Albatraia | Poggio Lecci | Cupellaio |
|---|---|---|---|
| Età (anni) | 56 | 59 | 56 |
| Quota (m s.l.m.) |
130 | 280 | 140 |
| Esposizione | nord-est | est | nord |
| Pendenza | 15% | 18% | 25% |
| Suolo | Mediamente profondo, poco pietroso su verrucano | Superficiale, roccioso su calcare | Superficiale, roccioso e pietroso su verrucano |
Albatraia, ceduo matricinato a prevalenza di leccio con presenza importante di orniello (Fraxinus ornus L.) e elementi residui della macchia mediterranea;
Poggio Lecci, ceduo matricinato di leccio con presenza importante di fillirea (Phillyrea angustifolia L.);
Cupellaio, macchia mediterranea a prevalenza di corbezzolo (Arbutus unedo L.), erica (Erica arborea L.) e fillirea con un piano superiore di matricine di leccio, roverella (Quercus pubescens L.) e sughera (Quercus suber L.).
All’interno delle aree tutti gli individui sono stati numerati progressivamente e sono stati eseguiti rilievi dendrometrici e strutturali, numero di ceppaie, polloni e matricine ad ettaro; definizione della specie, diametro a 1.30 m, altezza totale; posizione sociale individuale e delle ceppaie secondo tre classi (dominante, intermedia, dominata, si definisce ceppaia dominante quella con almeno 1 pollone dominante); collocazione spaziale delle ceppaie e delle matricine, nel 1991, al momento della istituzione delle aree di monitoraggio e nel 2006.
L’elaborazione dei dati ha permesso la determinazione dell’area basimetrica (G) e dell’incremento periodico sia monitorato (Ipm), ovvero la differenza tra i valori di G rilevati nel 1991 e quelli registrati nel 2006, sia effettivo (Ipe), vale a dire l’incremento dei soli polloni che sono sopravvissuti durante il quindicennio di monitoraggio.
Inoltre, per verificare il dato di età desunto dai registri aziendali attraverso la data dell’ultima ceduazione effettuata, in ciascuna area sono state prelevate 10-15 carotine delle varie specie presenti (leccio, orniello, corbezzolo, fillirea, erica, alloro, carpino).
L’analisi dei dati (Tab. 2) ha permesso di evidenziare le differenze di fertilità e le variazioni di altezza, densità, accrescimento e ruolo sociale verificatesi nel periodo di tempo compreso tra i due inventari. Il peso funzionale di ciascuna specie è stato determinato calcolando l’indice di importanza come definito da Chapman et al. ([12]), mentre la diversità specifica dello strato arboreo ed arbustivo è stata valutata sia in numero (SHn) che in area basimetrica (SHg) attraverso l’indice di Shannon ([47]).
Tab. 2 - Parametri utilizzati per quantificare e qualificare i processi dinamici in atto nelle aree monitorate.
| Parametro | Indicatore | Simbolo | Significato funzionale |
|---|---|---|---|
| Altezza dominante |
Altezza media dei 100 individui di diametro maggiore ad ettaro |
Hd (m) | Definisce la fertilità della stazione |
| n di specie | Variazione nel tempo del numero di specie presenti |
N specie | Valutano la diversità specifica dello strato arboreo e arbustivo |
| Indice di Shannon |
SH = Σ(pi · log pi) | SHn e SHg | |
| pi = densità relativa o area basimetrica relativa della specie i | |||
| Densità | Numero di polloni ad ettaro, totale e per specie | N (n ha-1) | Quantificano, in assenza di trattamento, la capacità portante della stazione |
| Accrescimento | Area basimetrica ad ettaro, totale e per specie | G (m2 ha-1) | |
| Mortalità | Variazione del numero di polloni nel tempo | M (%) | Definiscono i processi di selezione e competizione in atto nel popolamento |
| Tasso di accrescimento |
Variazione dell’area basimetrica nel tempo |
Ip (m2 ha-1) | |
| Indice di importanza |
I = 100 · (ni/N + gi/G)/2 | I (%) | Stima il peso funzionale di ciascuna specie |
| ni/N e gi/G =densità e area basimetrica relativa della specie i | |||
| Ruolo sociale | Variazione della classe sociale nel tempo | (%) | Definiscono la struttura verticale del soprassuolo |
| Indice di diversità strutturale |
VE = Σ(pi · ln pi) | VE | |
| pi = densità relativa nel piano di altezza i-esimo (3% < i < 50% di Hmax; 50% < i < 80% di Hmax; > 80% di Hmax) |
Risultati e discussione
Età dei soprassuoli e fertilità stazionale
La lettura degli anelli al 2006 ha confermato che l’età dei popolamenti studiati è simile nelle tre aree di ricerca e corrisponde a quella desunta in base all’ultimo taglio effettuato: 56 anni per le aree di Cupellaio ed Albatraia (ultima ceduazione nel 1950) e 59 anni per quella di Poggio Lecci (ultima ceduazione nel 1947 - [39]).
La differenza di età tra le tre aree é molto modesta così che possiamo, in prima approssimazione, ritenere le differenze di altezza dominante (Fig. 3) come una buona espressione della fertilità. Le altezze sono nettamente maggiori ad Albatraia, area scarsamente acclive con suolo profondo e poco pietroso, rispetto alle altre due aree dove alla maggior pendenza del terreno si associano suoli superficiali e rocciosi. Sembrerebbe quindi che la fertilità stazionale sia condizionata non tanto dal tipo di substrato, calcare a Poggio Lecci e verrucano ad Albatraia e Cupellaio, ma da una diversa giacitura, profondità, rocciosità e pietrosità ([37]).
Fig. 3 - Valori di altezza dominante (Hd) nelle tre aree di monitoraggio ai due inventari e incremento periodico dell’altezza media (IpHm) per le diverse specie.
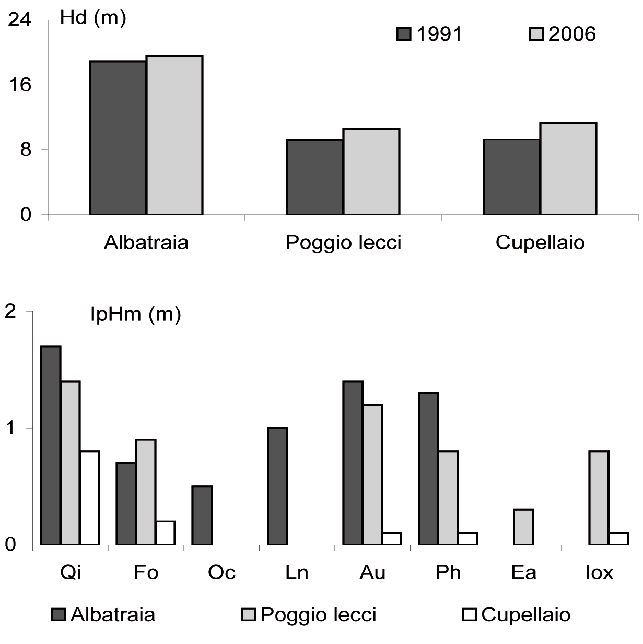
La variazione nel tempo dei valori di altezza media (IpHm) delle varie specie, anche se di scarsa entità in tutte e tre le aree, è risultata dipendente dalla fertilità stazionale. Ad Albatraia sono stati registrati i valori incrementali maggiori per tutte le specie censite, al contrario di Cupellaio i cui incrementi, significativamente minori rispetto alle altre due aree (p < 0.01), indicano un processo di accrescimento longitudinale estremamente lento. Il leccio mostra il maggior incremento periodico in tutte e tre le aree (1.7, 1.4 e 0.8 m rispettivamente ad Albatraia, Poggio Lecci e Cupellaio) anche se non sono state registrate differenze significative tra le singole specie.
Biodiversità
La diversa fertilità stazionale ha ovviamente condizionato sia la composizione specifica sia l’evoluzione delle strutture nel tempo ([24]). Complessivamente nei tre popolamenti analizzati sono state rilevate 11 specie arboree ed arbustive, anche se all’interno di ogni singola area non compaiono mai più di 8 specie. Alcune di esse caratterizzano tutte le aree e tra queste il leccio, l’orniello, il corbezzolo e la fillirea; altre, quali sughera, roverella, ginepro, alloro e carpino sono invece esclusive di ciascuna area (Tab. 3).
Tab. 3 - Variazione nel tempo delle specie presenti e dell’indice di Shannon, calcolato in numero - SHn - e in area basimetrica - SHg - nelle tre aree di studio.
| Specie | Albatraia | Poggio Lecci | Cupellaio | |||
|---|---|---|---|---|---|---|
| 1991 | 2006 | 1991 | 2006 | 1991 | 2006 | |
| Leccio | X | X | X | X | X | X |
| Roverella | - | - | - | - | X | X |
| Sughera | - | - | - | - | X | X |
| Orniello | X | X | X | X | X | X |
| Carpino nero | X | X | - | - | - | - |
| Alloro | X | X | - | - | - | - |
| Corbezzolo | X | X | X | X | X | X |
| Fillirea | X | X | X | X | X | X |
| Mirto | X | - | - | - | X | X |
| Erica arborea | - | - | X | X | X | X |
| Ginepro | - | - | X | X | - | - |
| SHn | 1.04 | 1.01 | 1.01 | 1.00 | 1.04 | 1.05 |
| SHg | 0.09 | 0.07 | 0.08 | 0.07 | 1.06 | 1.07 |
Nelle aree di Albatraia e Poggio Lecci la biodiversità espressa in area basimetrica (Shg - [13], [22]) è risultata inferiore rispetto a quella definita dal numero di individui (SHn) per l’elevato peso relativo in area basimetrica delle matricine, tutte di un’unica specie, ossia di leccio. Al contrario, nell’area di Cupellaio, la biodiversità in area basimetrica (SHg) è risultata maggiore poiché tra le matricine sono state censite anche specie diverse dal leccio quali roverella e sughera. Non sono state registrate sostanziali variazioni dell’indice nel tempo, anche se si può osservare una tendenza alla riduzione della biodiversità nell’area di Albatraia e un leggero aumento in quella di Cupellaio.
In relazione alla presenza della sughera, occorre sottolineare che in passato la specie era molto più diffusa; i registri aziendali del 1936 segnalano nelle sezioni di Cupellaio e di Albatraia, che si trovano su rocce acide del verrucano, un totale di 1050 sughere su 103.37 ha, di cui circa la metà da demaschiare e quindi presumibilmente giovani. Nel 1961 ne erano rimaste solo 400 e lo stesso numero è stato rilevato anche nel 1972. Alla data del primo rilievo di monitoraggio, ossia nel 1991, la sughera era assente nell’area di studio di Albatraia, non sappiamo se per la piccola dimensione dell’area rispetto alla scarsa densità delle sughere o se per la scomparsa di questa specie, nettamente eliofila, a causa della concorrenza.
Mortalità e accrescimento in area basimetrica
Le potenzialità stazionali espresse dai tre popolamenti si riflettono sui parametri dendrometrici considerati (Tab. 4). A fronte di un’area basimetrica totale praticamente simile, il numero di ceppaie e di polloni ad ettaro varia in modo considerevole. Ad Albatraia, ceduo matricinato a prevalenza di leccio, l’elevata mortalità (47% dei polloni), indice di una forte competizione, si è concretizzata in un valore di incremento periodico monitorato estremamente ridotto (IPm = 1.1 m2 ha-1); in realtà il buon livello di accrescimento individuale dei polloni e delle matricine e le dinamiche positive in atto nel soprassuolo sono espresse dal valore dell’incremento periodico effettivo (IPe = 9.5 m2 ha-1 - [21]). Il popolamento di Poggio Lecci (ceduo matricinato di leccio) mostra una tendenza dinamica simile a quella già evidenziata per Albatraia: mortalità elevata dei polloni (42%) e incremento periodico effettivo simile (IPe = 9.1 m2 ha-1); si mantiene tuttavia la maggiore densità iniziale rispetto all’area precedente. Il popolamento di Cupellaio (soprassuolo a macchia mediterranea) è caratterizzato da una dinamica più lenta, tipica di queste formazioni allorché occupano stazioni con scarsa fertilità. Il numero di polloni rimane molto alto; la mortalitàè ridotta (29%) e l’accrescimento più limitato (IPe = 5.6 m2 ha-1).
Tab. 4 - Area basimetrica totale (G) e numero ad ettaro di ceppaie (Cep), polloni (Pol) e matricine (Mat) nelle tre aree di studio, ai due inventari (M % = mortalità nel periodo; Ipe = incremento periodico effettivo = G2006-(G1991-Gmorti 1991-2006); Ipm = incremento periodico monitorato = G2006-G1991).
| Inventario | Albatraia | Poggio Lecci | Cupellaio | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G m2 ha-1 |
Cep n ha-1 |
Pol n ha-1 |
Mat n ha-1 |
G m2 ha-1 |
Cep n ha-1 |
Pol n ha-1 |
Mat n ha-1 |
G m2 ha-1 |
Cep n ha-1 |
Pol n ha-1 |
Mat n ha-1 |
|
| 1991 | 37.4 | 1150 | 3180 | 80 | 34.4 | 3033 | 10234 | 83 | 31.9 | 5733 | 14666 | 67 |
| 2006 | 38.5 | 730 | 1670 | 70 | 38.4 | 2251 | 5933 | 83 | 34.7 | 4816 | 10417 | 67 |
| M (%) | - | 37 | 47 | 13 | - | 26 | 42 | 0 | - | 16 | 29 | 0 |
| IPm | 1.1 | - | - | - | 4.0 | - | - | - | 2.8 | - | - | - |
| IPe | 9.5 | - | - | - | 9.1 | - | - | - | 5.6 | - | - | - |
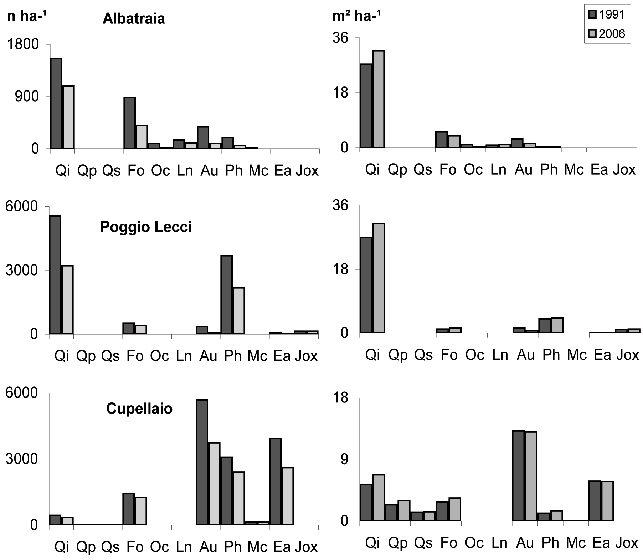
La variazione nel periodo di monitoraggio (1991-2006) del numero di individui e dell’area basimetrica per specie (Fig. 4) ha permesso di apprezzare l’importanza relativa di ciascuna di esse ed in particolare il ruolo del leccio. Questa specie risulta, in entrambi gli inventari, la principale sia ad Albatraia che a Poggio Lecci; tuttavia, mentre ad Albatraia le specie della macchia sono una componente limitata, a Poggio Lecci è la fillirea a mantenere una importanza numerica elevata. Al contrario, nell’area di Cupellaio, l’85% della composizione specifica è costituito da arbusti di macchia; tra questi, quelli più rappresentati sia in numero che in area basimetrica, sono il corbezzolo e l’erica, mentre il leccio è presente principalmente come matricina.
Fig. 4 - Numero di individui ed area basimetrica ad ettaro per specie ai due inventari, nelle tre aree. Tenere presenti le differenze di scala in ordinata.
Le analisi della mortalità e del tasso di accrescimento (incremento percentuale di area basimetrica), effettuati separatamente per le varie specie, hanno evidenziato che i due parametri, indicativi del dinamismo in atto nelle tre aree, risultano inversamente correlati: a mortalità superiori al 50%, corrisponde in genere una notevole riduzione dell’area basimetrica. Ad Albatraia (Fig. 5), nel periodo tra i due inventari, la mortalità del leccio è stata pari al 30%, assai inferiore a quella delle altre specie arboree e di macchia. L’elevata mortalità si è concretizzata nel decremento dell’area basimetrica per tutte le specie ad eccezione del leccio e dell’alloro, che hanno invece espresso un tasso di accrescimento positivo.
Fig. 5 - Mortalità e tasso di accrescimento registrato nel periodo di monitoraggio per specie, nelle tre aree.
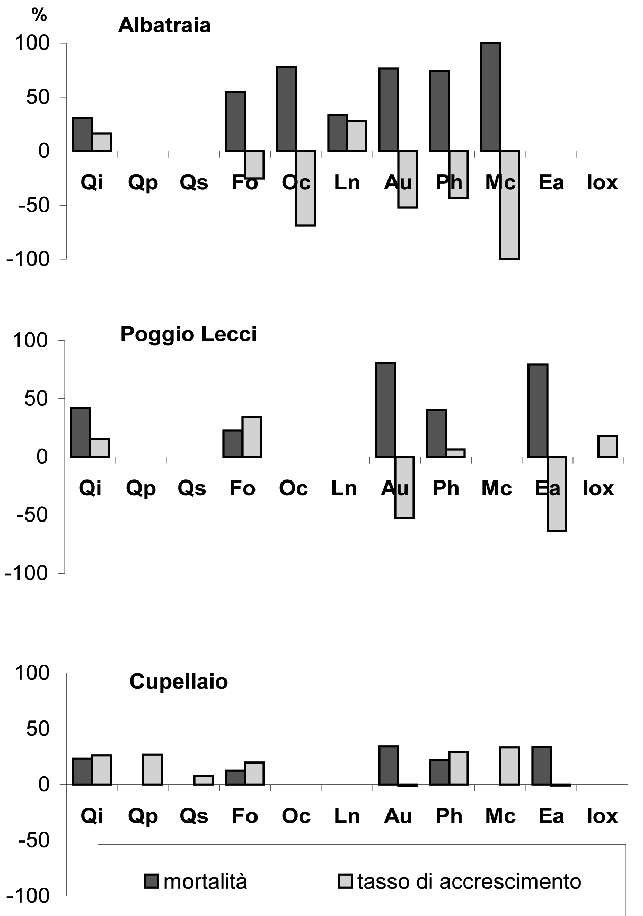
A Poggio Lecci, valori negativi del tasso di accrescimento sono stati registrati solo per il corbezzolo e per l’erica, specie caratterizzate da mortalità elevata (80% circa). Il leccio, nonostante una mortalità del 42%, ha mantenuto invece un accrescimento positivo.
L’area di Cupellaio, a differenza delle due precedenti, non registra casi di accrescimento negativo per alcuna delle specie componenti. La mortalità si mantiene infatti sempre su livelli molto inferiori al 50%, lontana quindi dalla soglia che abbiamo visto determinare un decremento in area basimetrica. In questa area, i saggi di accrescimento sono relativamente uniformi tra le specie arboree (compreso il leccio) e quelle della macchia. La minore fertilità e la tipologia compositivo-strutturale diversa producono quindi una dinamica differente nel periodo osservato. Questa si concretizza in un maggiore equilibrio tra mortalità e accrescimento sia a livello di specie che di popolamento.
Importanza relativa
Il peso numerico e funzionale delle singole specie presenti in ciascuna area è stato riassunto nel valore dell’indice di importanza relativa (Fig. 6), attraverso il quale sono state valutate le variazioni di presenza e consistenza nel tempo. L’andamento dell’indice conferma quanto già osservato attraverso l’analisi della mortalità e dell’accrescimento.
Fig. 6 - Indice di importanza relativo delle diverse specie ai due inventari, nelle tre aree.
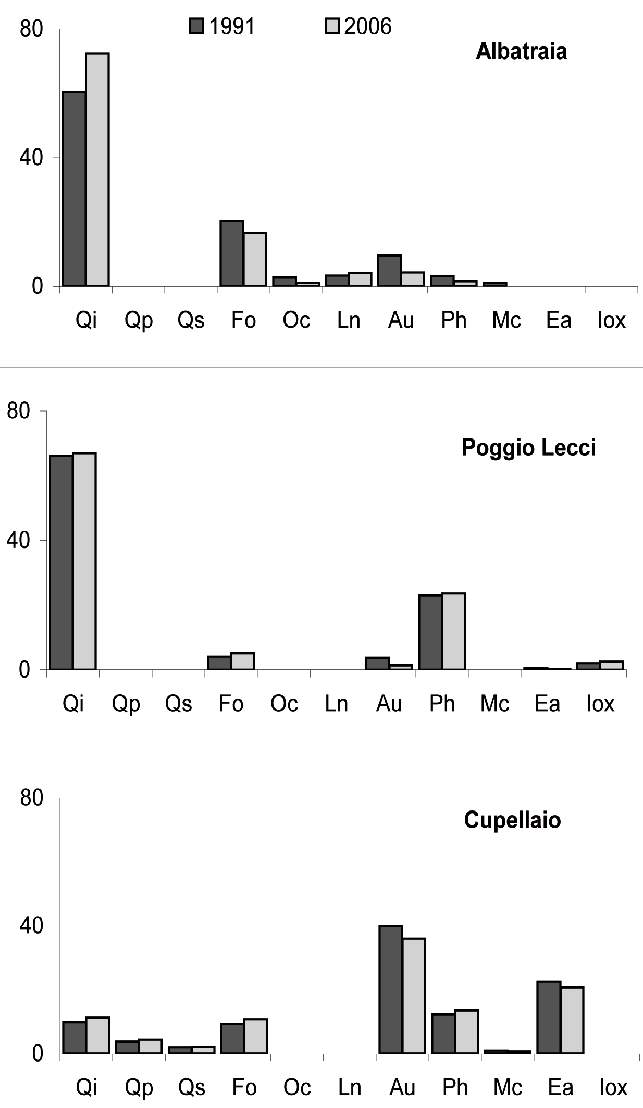
Ad Albatraia l’indice di importanza del leccio è aumentato nel periodo dal 60 al 72%, mentre si è ridotto quello delle altre due specie arboree (orniello e carpino nero) e di quelle della macchia, ad eccezione dell’alloro. Da segnalare inoltre la scomparsa del mirto (Myrtus communis L.). Al contrario, nelle aree di Poggio Lecci e Cupellaio, l’indice è rimasto pressoché invariato per il leccio come per le altre specie. A Poggio Lecci, l’importanza del leccio è comunque consistente (67% nel 2006) e la fillirea segue come seconda specie (24% nel 2006). A Cupellaio il leccio mantiene un peso numerico e funzionale pressoché costante, pari a circa il 10%.
Ruolo sociale e struttura verticale
L’esame della variazione nel tempo della componente dominante del soprassuolo espressa come numero di ceppaie dominanti e distinta per specie (Tab. 5), ha evidenziato l’aumento di questa categoria sociale nell’area di Albatraia (da 29 a 36%), la presenza stabile in quella di Poggio Lecci (da 29 a 30%) e un consolidamento della scarsa differenziazione sociale a Cupellaio (da 22 a 10%). Nelle aree di Albatraia e Poggio Lecci, già al primo inventario, la maggior parte delle ceppaie dominanti era di leccio, mentre a Cupellaio prevaleva il corbezzolo. La presenza nel piano dominante del leccio aumenta nei 15 anni di osservazione in tutte le aree monitorate (da 46 a 69% ad Albatraia; da 67 a 79% a Poggio Lecci e da 10 a 32% a Cupellaio).
Tab. 5 - Percentuale delle ceppaie dominanti ai due inventari, nelle tre aree di studio.
| Specie | Albatraia | Poggio Lecci | Cupellaio | |||
|---|---|---|---|---|---|---|
| 1991 | 2006 | 1991 | 2006 | 1991 | 2006 | |
| Totale | 29 | 36 | 29 | 30 | 22 | 10 |
| Leccio | 46 | 69 | 67 | 79 | 10 | 32 |
| Roverella | - | - | - | - | 1 | 4 |
| Sughera | - | - | 1 | 4 | ||
| - | - | |||||
| Orniello | 37 | 23 | 8 | 7 | 15 | 12 |
| Carpino nero | 9 | 4 | - | - | - | - |
| Alloro | 6 | 4 | - | - | - | - |
| Corbezzolo | 3 | 0 | 4 | 5 | 57 | 48 |
| Fillirea | - | - | 13 | 10 | 1 | 0 |
| Mirto | - | - | - | - | - | - |
| Erica arborea | - | - | - | - | 15 | 0 |
| Ginepro | - | - | 8 | 0 | - | - |
Le differenti modalità di strutturarsi nel piano verticale dei tre popolamenti (Tab. 6) risultano condizionate sia dalle caratteristiche ecologiche delle specie prevalenti (specie più o meno tolleranti dell’ombra) sia dalla diversa fertilità stazionale che ha inciso sull’inizio e sulla velocità di risoluzione dei fenomeni competitivi ([20], [23]).
Tab. 6 - Indice di diversità della struttura verticale ai due inventari, nelle tre aree di studio.
| Area | 1991 | 2006 |
|---|---|---|
| Albatraia | 0.97 | 1.02 |
| Poggio Lecci | 1.07 | 1.08 |
| Cupellaio | 0.23 | 0.53 |
Dove predomina il leccio (aree di Albatraia e Poggio Lecci) l’indice di diversità verticale definisce popolamenti pluristratificati in cui ciascun piano di altezza risulta ugualmente rappresentato anche se diversamente composto ([22]). Nell’area di Albatraia le migliori potenzialità stazionali hanno determinato una forte riduzione delle specie più eliofile della macchia mediterranea e permesso la permanenza di quelle più tolleranti dell’ombra, quali leccio, corbezzolo, alloro, nei piani inferiore e intermedio. Il leccio, peraltro, risulta equamente rappresentato in tutti e tre i piani mentre l’orniello è concentrato essenzialmente in quello superiore. Nell’area di Poggio Lecci la divisione tra i piani è invece molto netta: nel piano inferiore si ritrovano esclusivamente le specie della macchia mentre il leccio è concentrato unicamente nei piani superiore ed intermedio.
Al contrario nell’area di Cupellaio, dove predominano le specie della macchia a temperamento più eliofilo, i bassi valori dell’indice identificano una struttura notevolmente semplificata in cui la maggior parte degli individui è concentrata nei piani inferiore ed intermedio mentre in quello superiore sono presenti, oltre alle matricine di quercia, alcuni esemplari di orniello.
Conclusioni
Le tre popolazioni esaminate presentano differenze quali-quantitative originate:
- dalle forme di uso pregresse che hanno inciso più o meno profondamente sulla struttura e la composizione specifica originale;
- dalla auto-ecologia delle specie le quali, a seguito di questi disturbi, hanno modificato la propria presenza in termini di abbondanza e dominanza;
- dalle diverse condizioni di fertilità stazionale.
A partire da queste differenze, la sospensione di qualsiasi forma di trattamento (taglio) e di uso (pascolo) negli ultimi cinquanta anni, e ancora prima l’assenza di disturbi importanti documentati (incendi), ha consentito comunque uno sviluppo non condizionato da fattori esterni ai caratteri della stazione ([33]).
Questa è la situazione osservata nei 15 anni di monitoraggio, fino all’attualità. Un periodo così limitato ha consentito tuttavia di misurare variazioni relative nella composizione specifica e nel tasso di accrescimento.
Un tasso di mortalità elevato è comune alle due aree di ceduo, mentre è più ridotto nell’area di macchia. Se si considera il dato a livello di specie, nelle prime due aree (Albatraia e Poggio Lecci) la mortalitàè molto elevata per le specie arboree secondarie e per tutte le specie di macchia presenti, mentre per il leccio è minore. Una dinamica molto diversa avviene nell’area di macchia (Cupellaio), dove la riduzione numerica delle singole specie riflette il dato complessivo e appare bene bilanciata tra quasi tutte quelle presenti. L’accrescimento monitorato nelle due aree a ceduo risente, evidentemente, della elevata mortalità e registra valori molto bassi; quello reale è viceversa molto superiore e concentrato sulle specie arboree (leccio soprattutto). Un minore accrescimento (pari a circa la metà) si registra nell’area di macchia, dove però ancora tutte le specie contribuiscono similmente al valore complessivo. L’indice di importanza relativa conferma quanto definito in termini di mortalità ed accrescimento.
Le variazioni di tipo dendrometrico si traducono in modo diverso nel peso della componente dominante: questa mantiene valori all’incirca stabili nel ceduo matricinato di leccio (Poggio Lecci), aumenta nel ceduo matricinato in cui il leccio è prevalente ma condivide il piano arboreo con una seconda specie (orniello), recessiva rispetto alla quercia (Albatraia), e diminuisce sensibilmente nell’area di macchia (Cupellaio). Se però si qualifica il carattere di dominanza a livello di specie, si osserva come le ceppaie dominanti di leccio aumentino sensibilmente la loro presenza relativa in tutte le aree, indipendentemente dalla consistenza numerica iniziale.
I processi in atto nelle aree monitorate hanno permesso di definire non solo il ruolo prioritario della fertilità stazionale e della composizione specifica nella dinamica della comunità di specie arboree ma anche di interpretare la dinamica di lungo periodo ([34]). Nelle aree di Poggio Lecci e Albatraia, il gradiente di fertilità del suolo, associato a pendenza ed esposizione simili, trova riscontro nella maggiore presenza di specie di macchia dove la fertilitàè presumibilmente minore data la elevata rocciosità (Poggio Lecci) senza condizionare, almeno nella fase osservata, la dinamica positiva del leccio. Nel lungo periodo ci si può attendere un’evoluzione verso la lecceta con riduzione progressiva della complessità strutturale e della biodiversità. Nell’area di Cupellaio, composta prevalentemente da specie di macchia, sono state al contrario osservate dinamiche lente che mantengono nel tempo una configurazione fisionomica, tassonomica e strutturale stabile.
La diversa composizione di Cupellaio potrebbe testimoniare già in origine una struttura arborea rada, creata e mantenuta tale per il pascolo. Questo disturbo può avere ridotto molto la capacità di germinazione e di ricaccio del leccio e favorito la diffusione delle specie di macchia, una volta diminuita e poi cessata la pressione del pascolo. La differenza evolutiva con l’area contigua di Albatraia, pure pascolata, potrebbe essere dovuta alla maggiore pendenza del terreno, associata a suoli superficiali, pietrosi e con affioramenti di roccia che risulta sfavorevole al leccio ma più adatta all’insediamento di una macchia secondaria. Si conferma così come la variazione microstazionale, anche di una sola componente, possa dare origine a soluzioni evolutive differenti in ambienti difficili per la vegetazione forestale, come quello mediterraneo.
Già questi primi risultati confermano come boschi, sottoposti a forme di utilizzo multiplo e intensivo ed in condizioni ambientali limitanti, siano caratterizzati da una lenta e diversificata dinamica in termini di struttura, composizione specifica e accrescimento. Si conferma l’utilità del monitoraggio di lungo periodo su aree permanenti, sia per approfondire le conoscenze sulle dinamiche evolutive spontanee sia per valutare le strategie di conservazione e di gestione di questi soprassuoli.
Ringraziamenti
Si ringrazia sentitamente il personale del CRA-SEL, Maurizio Piovosi, Claudia Becagli e Orazio Ivan Gugliotta e il dottore forestale Andrea Ciofini per il prezioso aiuto nella fase di rilievo dei dati sperimentali. Si ringrazia altresì il Parco Regionale della Maremma e l’Azienda di Alberese per aver reso possibile la realizzazione della presente ricerca. Lo studio è stato in parte finanziato dal progetto RISELVITALIA, sottoprogetto 3.2.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar