Gap size effects on tree regeneration in afforestations of Black pine (Pinus nigra Arn.)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 6, Pages 312-319 (2009)
doi: https://doi.org/10.3832/efor0591-006
Published: Sep 18, 2009 - Copyright © 2009 SISEF
Research Articles
Abstract
The authors deal with the problem of the re-naturalization of afforestations of Black pine in the Apennines (Central Italy). Gap-cutting of two different sizes (the gap diameter/stand height ratios were respectively 0.75 and 1.0) on age classes stands (respectively 50 and 90 years old) have been carried out. The results of the dynamics of the tree regeneration established in the gaps 7 years after the beginning of the experiment are given. The Black pine represents the best regenerated tree species and its presence is increasing. Some broadleaves, Pubescent oak (Quercus pubescens Willd.) and Holm oak (Quercus ilex L.) particularly, and Hop-hornbeam (Ostrya carpinifolia Scop.) and Manna-ash (Fraxinus ornus L.) on a subordinate way, are also present in the gaps. These results confirm that the gap-cutting system represents an effective instrument for the natural regeneration priming and a low impact environmental treatment which is particularly fit for the re-naturalization of the conifers’ monocultures. According to this experience the gaps’ optimal sizes are 150-250 m2. These gaps should be distributed appropriately in space and time.
Keywords
Gap-size, Black pine, Stand management, Gap-cutting system, Re-naturalization
Introduzione
In Italia sono stati realizzati molti rimboschimenti per scopi protettivi a prevalenza di conifere e in particolare con pino nero.
All’opera di rimboschimento non hanno fatto seguito adeguate cure colturali. In particolare, l’assenza di diradamenti ha provocato una eccessiva densità delle pinete determinando: fragilità strutturale, forte rallentamento dei processi dinamici, problemi fitosanitari. Inoltre questi rimboschimenti hanno raggiunto età comprese tra 60 e 100 anni, prossime alla fase della maturità biologica (il tempo di permanenza della pineta non dovrebbe superare i 100 anni) per cui si rende necessaria e indilazionabile una azione colturale mirata ad assicurare ai popolamenti un maggior livello di stabilità, indirizzando i soprassuoli verso una maggiore complessità, sotto il profilo sia della struttura spaziale che della composizione specifica.
La rinaturalizzazione delle monocolture di conifere (qui intesa come trasformazione in popolamenti con una maggiore diversità compositiva e strutturale) rappresenta uno dei principali obiettivi selvicolturali anche in molti Paesi europei dove in passato sono state eseguite vaste piantagioni sia su terreni nudi che in sostituzione di formazioni naturali di latifoglie ([21], [29], [27], [23], [12], [44], [32], [11]). Tali azioni di lunga durata sono giustificate sul piano ecologico e economico per ottenere popolamenti con maggiore stabilità in termini strutturali e funzionali ([26], [39]).
Tra le varie opzioni selvicolturali proposte per la rinaturalizzazione delle monocolture di conifere ([28]), vengono prese in considerazione quelle basate sul principio che l’evoluzione naturale delle foreste è legata all’apertura di vuoti (buche, gap) nella copertura arborea per la morte di uno o più individui ([42], [33], [43], [30]).
La creazione di un sistema di buche è diventato un punto di riferimento per coloro che si occupano di gestione forestale e che cercano di emulare il regime dei disturbi naturali ([35], [8], [14]), in quanto il sistema in questione consente, tra l’altro, di incrementare la eterogeneità degli habitat e la complessità strutturale e compositiva dei popolamenti.
Il taglio a buche è stato proposto per i rimboschimenti di pino nero sia per rinnovare lo stesso pino che altre specie. Preto ([34]) prefigura per i rimboschimenti dell’Appennino romagnolo l’apertura di buche di dimensioni non superiori a 300 m2. Cucchi ([9]) individua nelle pinete di pino nero dell’Appennino ligure la possibilità di utilizzare il taglio a buche (di 200-400 m2) per favorire la rinnovazione di faggio e abete bianco. Bernetti ([2]) ritiene che il taglio a buche si possa applicare laddove si vuole conservare il pino. Secondo Malcolm et al. ([27]) le dimensioni ottimali delle buche per favorire la rinnovazione del pino laricio sono quelle dove il rapporto D/H (D = diametro della buca / H = altezza delle piante più alte) oscilla tra 1 e 2. Anche Bianchi et al. ([3]) ritengono che il taglio a buche sia un trattamento particolarmente adatto alla rinnovazione delle pinete di pino nero e laricio.
Dalle ricerche di Gugliotta et al. ([20]), eseguite in rimboschimenti di pino laricio, emerge che le buche grandi (1500 m2) favoriscono l’affermazione del pino laricio; diversamente la rinnovazione delle specie autoctone tardo-successionali sembra essere favorita dalle buche di dimensioni più piccole (380 m2). Ciò potrebbe far ritenere più appropriate le buche di piccole dimensioni se l’obiettivo fosse la rinaturalizzazione dei rimboschimenti.
Ai fini della rinaturalizzazione delle monocolture di conifere il taglio a buche può dunque assumere il duplice significato di:
- creare condizioni favorevoli all’insediamento e allo sviluppo della rinnovazione naturale;
- agevolare i processi dinamici di affermazione di altre specie che si sono insediate sotto copertura.
Il ruolo delle dimensioni delle buche sulla rinnovazione naturale è stato studiato nelle foreste gestite ([17], [40], [6], [7], [27], [12], [45], [13]).
In questo lavoro vengono analizzati gli effetti della dimensione di tagli a buche sulla rinnovazione naturale in pinete di pino nero. I risultati dopo 2 anni dall’esecuzione dei tagli a buche erano stati illustrati in un precedente lavoro ([19]), mentre in questa sede si riportano i risultati al termine di 7 anni di osservazioni.
Materiali e Metodi
Le aree sperimentali sono state eseguite a Monte Plaia, Comune di Introdacqua (AQ - Lat. 42° 01’ N; Long. 13° 54’ E) nel versante adriatico dell’Appennino abruzzese. In questa località furono eseguiti rimboschimenti in un arco di tempo compreso dal 1900 al 1955, il sistema di preparazione del suolo fu generalmente a gradoni, con densità d’impianto superiori a 3000 piante ad ettaro.
I dati climatici della zona sono riferiti alle stazioni termopluviometriche di Sulmona (420 m) e Goriano Sicoli (705 m): la precipitazione media annua varia da 746 a 1018 mm, quella estiva da 110 a 167 mm, la temperatura media annua da 9.7 a 11.8° C. Il substrato è costituito da calcari bianchi. I suoli appartengono ai Lithic Hapludoll ([36]), sono a profilo A-R con orizzonti superficiali dominati da materiale organico: poco decomposto nella parte superiore (Oi) e con un processo di decomposizione più avanzato in quello inferiore (Oe); l’orizzonte A è in genere profondo 30-40 cm con abbondanza di scheletro. La vegetazione naturale potenziale è rappresentata dai querceti decidui di roverella. In base alle indagini eseguite al momento dell’inizio dell’esperienza (Tiberi R, comunicazione personale), le diverse specie di insetti rilevate (Haematoloma dorsatum, Matsucoccus pini, Leucaspis pusilla, Pissodes notatus, Tomicus piniperda, Traumatocampa pityocampa) indicavano un precario stato vegetativo di queste pinete e quindi suggerivano la necessità urgente di interventi colturali.
Nella primavera 2000 furono eseguiti 4 tagli a buche, che comprendevano:
- 2 interventi in popolamenti di circa 50 anni, con rapporto tra il diametro della buca (D) e l’altezza delle piante più alte (H) rispettivamente di 0.75 e di 1;
- 2 interventi in popolamenti di circa 90 anni, con rapporto tra D e H, anche in questo caso, rispettivamente di 0.75 e di 1 (Tab. 1).
Tab. 1 - Monte Plaia. Caratteri dei tagli a buche.
| Codice area | Altitudine (m) |
Esp. | Pendenza (°) |
Età del popolamento (anni) | Altezza media (m) |
D/H |
|---|---|---|---|---|---|---|
| 1 | 830 | O | 17 | 50 | 14.1 | 1 |
| 2 | 830 | O | 17 | 50 | 13.9 | 0.75 |
| 3 | 840 | O | 25 | 90 | 18.2 | 1 |
| 4 | 850 | O | 24 | 90 | 17.3 | 0.75 |
Lo strascico del legname durante le operazioni di concentramento ed esbosco ha consentito la sostanziale rimozione degli orizzonti organici superficiali (lettiera di aghi, tappeti di muschi), dello strato erbaceo e della eventuale pre-rinnovazione esistente.
All’interno di ciascuna buca sono state delimitate 13 sub-aree circolari di 3.14 m2 (raggio 100 cm) per lo studio della rinnovazione naturale. Le sub-aree sono state posizionate: una al centro della buca e 3 lungo ogni direzione cardinale, con distanza dal centro pari rispettivamente a 1/3, 2/3, 3/3 il raggio della buca.
Di ogni semenzale ricadente nella sub-area è stata determinata la specie e misurata l’altezza in cm, stimata l’età per mezzo della conta dei flussi di accrescimento (dal conteggio sono stati esclusi i semenzali dell’anno) e l’eventuale presenza di danni da selvaggina. Non sono stati considerati gli eventuali polloni che potevano essersi originati da soggetti presenti prima della esecuzione dei tagli.
I rilievi sulla rinnovazione sono stati eseguiti nell’estate 2002 e 2007.
Per l’analisi statistica ci si è avvalsi del test non parametrico U di Mann-Whitney (Statistica, Statsoft®, Inc. Tulsa, OK, USA).
Nel 2007 sono stati eseguiti anche rilievi pedologici; il campionamento è stato eseguito con una trivella pedologica fino ad incontrare la roccia madre e ha interessato 3 prelievi all’interno di ogni buca e 3 all’interno nel bosco circostante.
I campioni di suolo sono stati asciugati all’aria e setacciati (< 2 mm) prima di effettuare le analisi chimico-fisiche. La granulometria è stata determinata con un idrometro Bouyoucos ([4]); il pH è stato determinato in sospensione, in rapporto suolo:acqua 1:2.5 e suolo:KCl (1M) 1:2.5; il carbonio organico (C.O.) è stato determinato mediante ossidazione col bicromato; la percentuale di sostanza organica (S.O.) è stata ottenuta moltiplicando la percentuale di C.O. per il fattore 1.72 ([41]). L’azoto totale è stato determinato utilizzando il metodo Kjeldahl ([24]), il fosforo disponibile con il metodo Bray II ([5]) e il potassio scambiabile mediante fotometro a fiamma.
I dati relativi all’analisi del suolo sono stati elaborati statisticamente utilizzando l’analisi della varianza (ANOVA). Le differenze tra i trattamenti sono state determinate utilizzando il Tukey’s test ([37]).
Risultati e Discussione
Caratteristiche della rinnovazione naturale dopo 7 anni
Dopo 7 anni dall’inizio dell’esperienza in tutte le buche prevale il pino nero in termini di densità, anche se le latifoglie si sono ben affermate, come mostrano i valori superiori di altezza media (Tab. 2). Tra le latifoglie le specie maggiormente rappresentate sono la roverella e il leccio, mentre sono sporadiche l’orniello e il carpino nero. La prevalenza di pino nero conferma, tra l’altro, che sussistono ancora nelle buche condizioni di elevata illuminazione favorevoli al suo sviluppo ([1], [22], [31], [3]). I valori di altezza media del pino sono superiori, in tutte le buche, nella parte centrale delle buche (sub-aree poste al centro e alla distanza di 1/3 dal centro della buca), anziché sugli orli delle buche (sub-aree poste alla distanza di 2/3 e 3/3 dal centro della buca - 32.0 vs. 12.3 cm), mentre i valori di densità sono sostanzialmente simili nel centro delle buche e negli orli (4.7 vs. 4.4 n/m2). Per le latifoglie i maggiori valori di altezza media si riscontrano nel centro delle buche (75.9 vs. 48.3 cm), mentre i valori di densità sono sostanzialmente simili nel centro delle buche e negli orli (0.44 vs. 0.49 n/m2 - dati non mostrati).
Tab. 2 - Monte Plaia. Rinnovazione naturale, 2007.
| Specie | Parametro | 1 | 2 | 3 | 4 |
|---|---|---|---|---|---|
| Pino nero | Altezza media (cm) | 25.5 ± 16.2 | 17.6 ± 10.2 | 34.8 ± 28.0 | 36 ± 24.6 |
| Densità (n. semenzali/m2) | 21.5 | 6.4 | 5.8 | 9 | |
| Età media (anni) | 4.4 | 4.1 | 4.4 | 4.4 | |
| Roverella | Altezza media (cm) | 26.6 ± 12.2 | 23.8 ± 9.6 | 75.5 ± 47.9 | 46.8 ± 31.8 |
| Densità (n. semenzali/m2) | 0.8 | 0.6 | 0.6 | 0.5 | |
| Età media (anni) | 3.8 | 3.6 | 4.5 | 3.9 | |
| Leccio | Altezza media (m) | 58.8 ± 45.7 | 39.7 ± 29.3 | 81.7 ± 44.9 | 87.5 ± 26.3 |
| Densità (n. semenzali/m2) | 0.5 | 0.6 | 0.2 | 0.1 | |
| Età media (anni) | 4.1 | 3.5 | 4.4 | 4.7 | |
| Orniello | Altezza media (cm) | 149 ± 0.0 | - | 80 ± 0.0 | - |
| Densità (n. semenzali/m2) | 0.02 | - | 0.02 | - | |
| Età media (anni) | 3 | - | 4 | - | |
| Carpino nero | Altezza media (cm) | - | - | 81 ± 12.7 | 121.4 ± 39.8 |
| Densità (n. semenzali/m2) | - | - | 0.04 | 0.17 | |
| Età media (anni) | - | - | 4 | 4.6 |
La roverella è la latifoglia che si è meglio rinnovata nelle buche, confermando le osservazioni fatte nelle pinete dell’Alta Valle dell’Aterno (Appennino abruzzese) a quote comprese tra 800 e 900 m, in esposizioni meridionali, su substrati calcarei ([16], [15]). L’orniello e il carpino nero, specie che dimostrano un carattere pioniero con ampie capacità colonizzatrici dei detriti calcarei, non hanno raggiunto una presenza significativa. Ciò potrebbe essere messo in relazione all’ambito bioclimatico non ottimale per le due specie: la tendenza dinamica è piuttosto in direzione dei querceti a dominanza di roverella.
Dal confronto dei valori di densità nelle buche con rapporto D/H = 1, emerge che la buca 1 presenta valori (soprattutto per il pino) superiori alla buca 3, dove i valori di densità dei semenzali risultano i più bassi, sia nei rilievi del 2002 che del 2007: il fenomeno è verosimilmente da attribuire al fatto che, nella buca 3, le operazioni di strascico del legname durante le operazioni di utilizzazione non asportarono completamente gli orizzonti superficiali, con la conseguenza che permangono settori con spessi strati di aghi e di muschi che impediscono la germinazione dei semi. Tuttavia l’altezza media dei semenzali di pino registrata nella buca 3 è maggiore rispetto a quella della buca 1. Le buche con rapporto D/H = 0.75 sono quelle dove sono stati rilevati i valori di densità minore (anche se leggermente superiori a quelli della buca 3): l’altezza media è in genere inferiore sia per il pino nero che per le latifoglie (Tab. 2). Anche se le differenze di densità dei semenzali tra le buche con rapporto D/H =1 e con rapporto D/H = 0.75 non sono significative in base all’analisi statistica, va osservato che comunque si sono registrate delle differenze.
L’età dei semenzali è rappresentata da diverse classi cronologiche (2-7 anni): ciò dimostra che la fase di insediamento della rinnovazione non si è esaurita nel primo anno di apertura della buca, ma che è continuata anche negli anni successivi.
La copertura erbacea e arbustiva è sostanzialmente assente e non pregiudica l’insediamento e l’affermazione della rinnovazione. Si è visto tuttavia che i tappeti di muschi, presenti soprattutto a margine delle buche per la mancata rimozione con lo strascico dei tronchi, rappresentano un ostacolo all’insediamento del novellame.
Non sono stati registrati danni prodotti da selvaggina sulla rinnovazione.
Nelle piante circostanti le buche, rese più vulnerabili in seguito al brusco isolamento, non si sono verificati danni meccanici da neve e/o vento (ad eccezione di 2 piante stroncate nella buca 2 e di una pianta stroncata nella buca 1 nel 2003).
Variazione dei valori (2002-2007) di altezza media e di densità della rinnovazione naturale
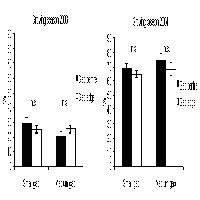
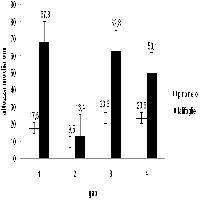
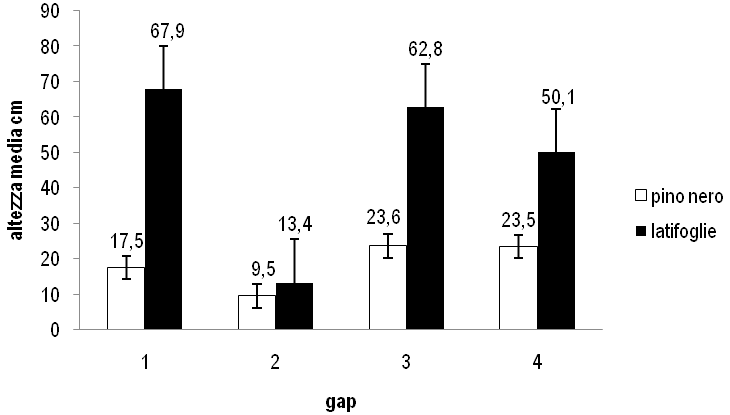
Gli incrementi di altezza media sono maggiori per le latifoglie rispetto al pino, soprattutto nelle buche con rapporto D/H = 1 (Fig. 1).
Fig. 1 - Variazione dei valori di altezza media nel periodo 2002-2007. Le barre indicano ES.
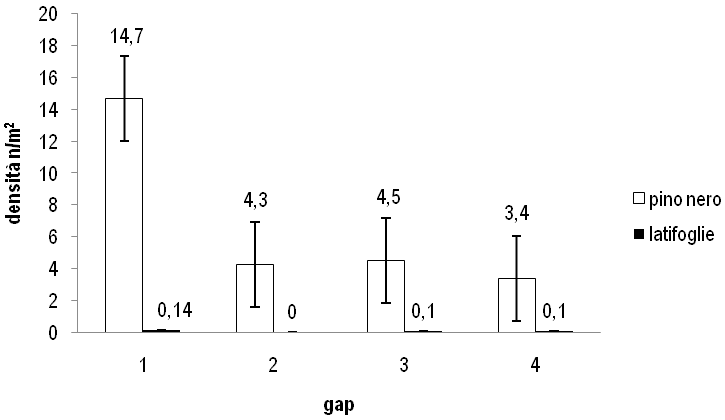
Si ha un incremento generalizzato di densità dei semenzali nel pino (in maniera più sensibile nella buca 1), a indicare che nel periodo di osservazione è proseguita la fase di insediamento del novellame della conifera.
La densità delle latifoglie non subisce in nessuna buca alcun incremento significativo, quindi la dinamica sembra al momento essersi stabilizzata (Fig. 2). Le differenze di incremento di densità tra pino e latifoglie non sono tuttavia significative all’analisi statistica.
Fig. 2 - Variazione dei valori di densità del novellame nel periodo 2002-2007. Le barre indicano ES.
Confronto tra le dimensioni delle buche (1 e 2) in popolamenti di 50 anni
I valori delle variazioni di altezza media e di densità del pino sono maggiori nella buca 1 rispetto alla buca 2. La percentuale di semenzali morti di pino è assai inferiore nella buca 1 (6.3%) rispetto alla buca 2 (51.3%). Anche le latifoglie mostrano nella buca 1 una variazione di altezza media nettamente superiore rispetto alla 2, mentre non si ha incremento di densità (Fig. 1 e Fig. 2). Tutto ciò può essere attribuito al fatto che la buca 1 è più grande della buca 2.
Confronto tra le dimensioni delle buche (3 e 4) in popolamenti di 90 anni.
Le variazioni dei valori di altezza media sono sostanzialmente simili nel pino, mentre sono maggiori nelle latifoglie nella buca 3 rispetto alla buca 4. Le variazioni di densità per il pino sono superiori nella buca 3, mentre sono simili e impercettibili per le latifoglie (Fig. 1 e Fig. 2). La percentuale di semenzali morti di pino è superiore nella buca 3 (2.9%) rispetto alla buca 4 (1.6%). Anche in questo caso si può attribuire alla maggiore dimensione della buca un effetto favorevole sulla rinnovazione.
Caratteristiche pedologiche dopo 7 anni
Nella Tab. 3 sono riportati i dati relativi alle analisi effettuate sul suolo prelevato nelle buche e nelle zone all’interno del bosco ad esse adiacenti, a 7 anni dall’inizio dello studio. Le buche rappresentano i campioni indicati con i numeri 1, 2, 3, 4, mentre per le zone all’interno del bosco limitrofe ad ogni buca i campioni sono indicati con i numeri 5, 6, 7, 8.
Tab. 3 - Caratteristiche chimiche del suolo determinate nelle buche (1-4) e nelle zone all’interno del bosco ad esse adiacenti (5-8). (*): i valori nella stessa colonna seguiti dalle stesse lettere non sono significativamente differenti per p≤0.05.
| Codice area | pH (H20) | pH (KCl) | C.O. (%) |
S.O. (%) |
CaCO3 (ppm) |
K2O | N (%) |
P2O5 (ppm) |
C/N | Classe Tessiturale |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 7.1b* | 6.0a | 3.18e | 5.46e | 0 | 397a | 4.2c | 7.2a | 7.5d | SF |
| 2 | 6.9d | 5.9a | 3.21e | 5.52e | 0 | 310e | 4.0d | 4.7b | 8.0c | SF |
| 3 | 7.0c | 5.9a | 3.18e | 5.29f | 0 | 382b | 4.6a | 6.8a | 6.6e | SF |
| 4 | 7.3a | 6.2a | 3.16e | 5.44e | 0 | 350c | 4.2c | 4.5c | 7.5d | SF |
| 5 | 6.9d | 6.1a | 3.45d | 5.96d | 0 | 259f | 3.9d | 4.2d | 8.8b | SF |
| 6 | 6.8d | 6.1a | 3.79c | 6.52c | 0 | 336d | 4.5a | 4.5c | 8.4c | SF |
| 7 | 6.9d | 6.1a | 3.94b | 6.78b | 0 | 225g | 4.3b | 4.4c | 9.1b | SF |
| 8 | 6.9d | 5.9a | 4.25a | 7.90a | 0 | 199h | 4.4b | 4.6b | 9.6a | SF |
I campioni rilevati all’interno del bosco sono caratterizzati da un maggior contenuto di sostanza organica rispetto ai campioni nelle buche (Tab. 3), con differenze significative. In particolare i campioni 7 e 8 (dei popolamenti di 90 anni) hanno un più elevato contenuto di sostanza organica. L’evoluzione della sostanza organica è indicata dal rapporto C/N, che è significativamente più elevato nei campioni 7 e 8. Questo indica che nei suoli sotto la copertura di popolamenti di 90 anni prevale il processo di umificazione. Il rapporto C/N è strettamente relazionato ai processi di trasformazione della sostanza organica e ai tempi di rilascio degli elementi nutritivi contenuti ([38]). Più bassi sono i valori del rapporto C/N maggiore sarà il rilascio di nutrienti e minore l’umificazione della sostanza organica. I più bassi valori di nutrienti (P e K) trovati nei campioni 7 e 8 suggeriscono una più lenta mineralizzazione della sostanza organica, avvalorando i risultati precedenti (Tab. 3). Il pH del suolo è neutro in tutti i siti studiati e la differenza tra pH determinato in acqua e quello in KCl si accentua nelle buche, quasi ad evidenziare differenze nel trend e nella costituzione della sostanza organica stessa. Durante il processo di mineralizzazione vi è una modificazione graduale della composizione elementare dei residui organici, la variazione di composizione riguarda soprattutto la diminuzione del contenuto di ossigeno e azoto e l’aumento del contenuto di idrogeno e carbonio. La differenza rilevata attraverso la determinazione del pH in H20 e in KCl può essere ascritta quindi alla quantità di ioni H+ rilasciati durante il processo di mineralizzazione della sostanza organica e presenti sui siti di scambio dei colloidi del suolo. Per quanto riguarda i caratteri pedologici delle buche, le migliori condizioni sono riferibili alla buca 1, dove è presente un buon contenuto di sostanza organica e un alto contenuto di nutrienti: questo suggerisce che il processo di mineralizzazione e umificazione sono ben bilanciati. Non si osservano significative differenze tra la buca 2 e 4, mentre nella buca 3 vi è il più basso valore di C/N insieme ad un elevato contenuto di nutrienti: ciò indica il prevalere del processo di mineralizzazione rispetto a quello di umificazione. I risultati ottenuti consentono di effettuare delle ipotesi di collegamento tra le caratteristiche pedologiche e quelle della rinnovazione naturale così, nella buca 1 (con rapporto D/H = 1), l’elevato contenuto di nutrienti insieme ad un buon processo di mineralizzazione-umificazione potrebbe favorire un’alta densità di semenzali di pino. La minore quantità di sostanza organica osservata nelle buche rispetto a quella all’interno del bosco è dovuta ad un diminuito apporto di lettiera e ad una più spinta mineralizzazione della sostanza organica stessa, probabilmente dovuta ad un maggiore apporto di radiazione solare e ad una più alta temperatura del suolo. Molti autori ([25], [18]) hanno infatti trovato una stretta relazione tra diametro dei tagli a buche e intensità della luce dimostrando che la quantità di radiazioni che arriva al suolo è direttamente proporzionale alla grandezza delle buche. Ciò comporta un aumento della temperatura del suolo, e di conseguenza del processo di mineralizzazione della sostanza organica. Analoghe osservazioni sono state fatte da Cutini et al. ([10]) in precedenti esperienze su tagli a buche nelle abetine delle Foreste Casentinesi (Appennino centro-settentrionale).
Conclusioni e implicazioni per la gestione
I risultati della presente indagine, soprattutto se confrontati con quelli del 2003, confermano che, in linea con quanto avviene quando si studiano i processi successionali, è possibile fornire risultati solo provvisori a causa del dinamismo cui è soggetta la vegetazione. In particolare è opportuno sottolineare che il pino è ancora in fase di espansione, fenomeno che al momento prevale sull’ingresso di latifoglie. Tuttavia quanto emerso dalle recenti osservazioni non sembra far discostare dalle ipotesi precedentemente avanzate ([19]) circa la tendenza dinamica della rinnovazione naturale all’interno delle buche, indirizzata verso la formazione di popolamenti a prevalenza di pino nero e roverella. Mentre la presenza della roverella, specie indice di un maggior stato di equilibrio, può far pensare ad una successiva definitiva affermazione, forse accompagnata da altre latifoglie, il ruolo del pino, per le sue caratteristiche di specie pioniera e preparatoria, dovrà essere ulteriormente analizzato e valutato nel corso del tempo, in quanto la rinnovazione di pino potrebbe diventare progressivamente sempre più subordinata.
La rinaturalizzazione significa un dinamismo verso la vegetazione potenziale della zona, rappresentata, in questo caso, dal querceto misto a dominanza di roverella.
Sul piano gestionale è prevedibile che in casi simili a quelli analizzati potrà essere applicato con successo, per avviare e velocizzare il processo di rinaturalizzazione di rimboschimenti di pino nero, un sistema di tagli a buche delle dimensioni di 150-250 m2 opportunamente distribuiti nello spazio e nel tempo. Tali dimensioni sono riferite, rispettivamente, ai popolamenti con altezza media di 14 m e a quelli con altezza media di 18 m: ovviamente buche più grandi dovrebbero favorire il pionierismo del pino.
Il taglio a buche induce complessi cambiamenti ecologici e biogeochimici nel suolo alterando il trend e la velocità di decomposizione della sostanza organica. Esso crea una dinamica ridistribuzione e ristrutturazione del pool organico accelerandone la decomposizione. La determinazione della sostanza organica e delle sue frazioni può essere un sensibile indicatore con effetti nella gestione forestale a breve e medio termine.
Inoltre, nonostante i vantaggi colturali ed ecologici di questo trattamento e l’apertura del mercato del cippato utilizzato per le caldaie a biomasse, gli interventi in questa area dell’Appennino abruzzese, avendo scarsa rimuneratività, richiedono il supporto del finanziamento pubblico.
Ringraziamenti
Gli Autori ringraziano il Dott. Francesco Mercurio per i rilievi e l’elaborazione dei dati.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar