Weak basis for “systemic” silviculture
Forest@ - Journal of Silviculture and Forest Ecology, Volume 6, Pages 274-276 (2009)
doi: https://doi.org/10.3832/efor0595-0006
Published: Sep 18, 2009 - Copyright © 2009 SISEF
Commentaries & Perspectives
Abstract
An analysis is presented of the ecological basis of the so-called “selvicoltura sistemica” (systemic silviculture). Systemic silviculture implies an “a-structured forest”: the meaning of this particular condition is discussed in the paper, exploring the different possible interpretations and thus demonstrating that an “a-structured” forest can not be observed in nature. The silvicultural method proposed therefore seems to have a fragile ecological basis moving it away from a close-to-nature forest management approach, which is currently believed to be the most promising one for achieving forests that provide multiple goods and services.
Keywords
Close-to-nature silviculture, forest ecology, ecosystem, forest structure, self-thinning law
La gestione di un ecosistema non può prescindere dalla conoscenza delle caratteristiche del sistema stesso. Relativamente agli ecosistemi forestali, significa che l’attività di gestione selvicolturale si debba basare sulla conoscenza della foresta, conoscenza che deriva, in primis, da una disciplina specifica: l’ecologia forestale. Essa è, quindi, il fondamento di ogni intervento e, per questa ragione, gli ecologi forestali si devono sentire chiamati in causa quando nuove modalità di gestione vengo discusse e proposte per applicazioni generalizzate.
Vorrei presentare una breve analisi su quella che viene definita “selvicoltura sistemica” ([1]) riguardo non, naturalmente, gli aspetti più specificatamente applicativi, su cui dichiaro la mia ignoranza, ma su quelli fondativi del metodo di gestione. Il mio scopo è dimostrare la debolezza e le contraddizioni della selvicoltura sistemica riguardo le sue basi ecologiche come evidenziato anche da altri autori ([7]). Tale debolezza pone seri interrogativi, quindi, anche sulla sua potenziale applicazione che è stata proposta al recente III Congresso Nazionale di Selvicoltura di Taormina.
Testualmente “la selvicoltura sistemica presuppone un bosco astrutturato, disomogeneo, autopoietico” ([1]). È la caratteristica della struttura “non-struttura” delle foreste sottoposte alla selvicoltura sistemica che voglio discutere nel dettaglio.
Ogni ecosistema si caratterizza da più attributi che, di norma, vengono riassunti come ([3]): struttura, funzione, interdipendenza, complessità dinamica temporale. Concentriamoci sui primi due attributi: la struttura (descrivibile delle caratteristiche fisiche del sistema) implica che le singole parti del sistema (individui popolazioni, comunità) abbiano un significato solo perché parti del tutto. Proprio perché le parti sono intimamente legate che si esplica una funzione complessiva, ossia si generano specifiche modalità di trasferimento di energia e materia tra le componenti. Struttura e funzione sono, quindi, strettamente interdipendenti ed intimamente connesse. Ad esempio, le caratteristiche della foresta (densità, area fogliare, frazione di strutture di sostegno, ecc.) determinano quanta energia la foresta possa mettere a disposizione per i consumatori e per i decompositori ed, in sintesi, il numero dei livelli trofici dell’intero ecosistema.
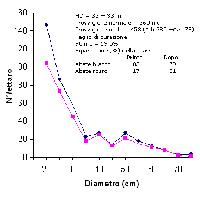
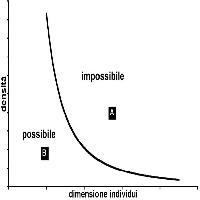
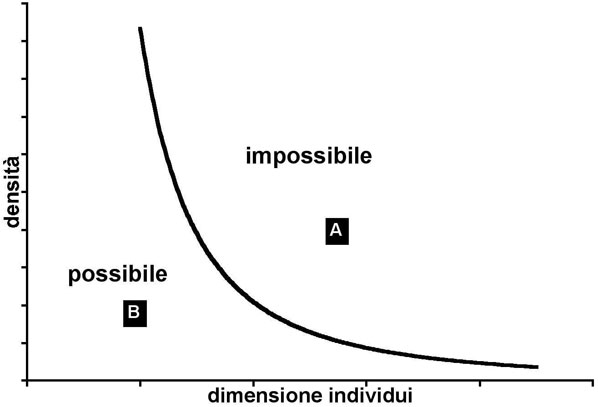
In tutti gli ecosistemi vi sono delle regole (a diverse scale): ad esempio, non è possibile che le componenti eterotrofe consumino più energia di quanta ne producano le componenti autotrofe. È noto, anche, che per garantire la sopravvivenza di una popolazione ci debba essere certa struttura cronologica ed uno specifico rapporto tra i sessi. Così come per le popolazioni animali una certa struttura cronologica è necessaria, così per la foresta, si potrebbe ipotizzare che una certa struttura debba essere presente per garantirne la sopravvivenza. Dato che per la foresta conta più la dimensione che l’età, ogni foresta dovrebbe caratterizzarsi per la presenza di un determinato numero di individui in ogni classe dimensionale. La quantificazione della numerosità delle coorti di diversa dimensione in una certa area viene utilizzata efficacemente dai forestali come rappresentazione della struttura della foresta (la curva di distribuzione diametrica). Anche riguardo la struttura ci sono delle regole: ad esempio non è possibile che 1000 individui piccoli occupino lo stesso spazio di 1000 individui molto grandi: necessariamente, questi ultimi, dovranno occupare un’area molto più ampia, ossia la densità deve diminuire in relazione alla dimensione media degli alberi. Questo processo detto di autodiradamento o self-thinning è molto noto ai forestali. Vi sono quindi, nella foresta, delle strutture possibili (Fig. 1), che si osservano in natura, e delle strutture impossibili (sopra la curva) che, infatti, non si osservano mai. La curva indica quale sia la massima numerosità per classe dimensionale in relazione alla massima compenetrazione delle chiome compatibile con la sopravvivenza degli individui.
Fig. 1 - Variazione della densità (N/ha) degli individui in funzione della loro dimensione (diametro o massa). Si osservano delle condizioni di struttura “impossibili” (condizione A ad esempio) e delle condizioni “possibili” (condizione B). Siccome la dimensione della chioma aumenta con l’aumentare del diametro, la curva rappresenta la condizione di massima compenetrazione delle chiome tra individui (che in genere è la minima possibile). Compenetrazioni troppo elevate non sono compatibili con la sopravvivenza degli individui che, quindi, vengono sottoposti ad un severo autodiradamento. Ne deriva che in una foresta non “tutte le condizioni” possano essere realmente osservate.
Torniamo, ora, alla foresta proposta dalla selvicoltura sistemica che dovrebbe tendere verso una condizione “astrutturata”. Tale condizione, mi sembra, possa essere interpretata in due soli modi. La prima interpretazione è che per “foresta astrutturata” ([1]) si intenda una foresta in cui si possono osservare tutti i tipi di struttura possibili. Tuttavia, abbiamo visto che non tutte le strutture sono possibili e quindi la condizione di “tutte le strutture” non è compatibile con quello che si osserva in natura.
L’altra interpretazione (che, in verità, mi sembra esclusa anche dalla selvicoltura sistemica stessa) è che il sistema sia “privo di struttura”, ossia ogni parte sia indipendente dalle altre che costituiscono il tutto o più semplicemente ogni albero sia indipendente dalla presenza degli alberi vicini. Questa seconda ipotesi porta alla paradossale conseguenza che una foresta astrutturata, visto il legame intimo tra struttura e funzione, sia anche “afunzionale” ossia senza funzione come entità complessiva, quindi morta.
Ne deriva la conclusione, in entrambe le interpretazioni, che la selvicoltura sistemica proponga, di fatto, una condizione non “in armonia con la natura” ([1]) ma in antitesi ad essa. Appare evidente, così, anche la sostanziale differenza rispetto alla selvicoltura naturalistica in cui, invece, determinati modelli strutturali naturali hanno sempre rappresentato la base di riferimento per la definizione degli interventi selvicolturali, come ci ha insegnato a Padova Lucio Susmel ([9], [10]).
Le conseguenze dell’applicazione della selvicoltura sistemica appaiono preoccupanti per i selvicoltori oltre che per gli ecologi. Infatti, viene proposto un procedimento tecnico basato sul sistema “tentativi ed eliminazione degli errori” ([1]) che ha fornito, in passato, anche indicazioni importanti ma che, ora, sta diventando via via sempre meno utile ([3]). Il vicolo cieco imboccato dalla selvicoltura sistemica risulta evidente: negando la possibilità da parte del gestore di immaginare e seguire specifici modelli strutturali non resta che la possibilità di lasciarlo completamente libero senza alcuna indicazione tecnica se non quella di valutare il risultato a posteriori (con significativa probabilità di compiere errori). La selvicoltura sistemica sembra alzare bandiera bianca di fronte alla possibilità dell’uomo di comprendere il funzionamento della foresta, che diviene, quindi, realtà“inattesa ed imprevedibile” ([1]). Tale approccio alla conoscenza della natura fa emergere ancora l’immane questione metafisica se il mondo sia o no regolato da leggi rigorose. Popper ([8]) propone di risolvere la questione affermando che “le leggi che scopriamo sono sempre ipotesi: e ciò significa che possono sempre essere soppiantate...(omissis). Tuttavia, negare la causalità sarebbe lo stesso che tentar di persuadere il teorico a rinunciare alla sua ricerca... (omissis). Anche la credenza nella causalitàè metafisica. Non è nient’altro che un’impostazione metafisica di una regola metodologia ben giustificata: la decisione dello scienziato di non abbandonare mai la ricerca delle leggi. Dunque la credenza metafisica nella causalità sembra più fertile, nelle sue varie manifestazioni della metafisica indeterministica. (omissis)”.
Che la “fertilità” della ricerca della causalità sia elevata è dimostrato dalla proposta di nuovi contributi riguardo i fattori guida della dinamica della foresta ([4], [11], [2], [5], [6]). In sostanza, quindi, la selvicoltura sistemica mi sembra animata dall’idea di (ri)portare le Scienze Forestali sotto il dominio dell’indeterminazione. Come ricercatori credo, invece, “dovremmo astenerci dall’emettere proibizioni che possano porre limiti alle possibilità di ricerca” ([8]). Per questo mi auguro che la ricerca delle leggi che regolano il funzionamento della foresta non venga trascurata ma, più che mai, fermamente promossa.
References
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar