Additive partition of diversity as standard for the selection of protected areas: the natural reserves network of the Siena province (Italy)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 7, Pages 28-43 (2010)
doi: https://doi.org/10.3832/efor0615-007
Published: Apr 01, 2010 - Copyright © 2010 SISEF
Research Articles
Abstract
Estimating the effects of the spatial components on species diversity represents an important step to establish the conservation “capacity” in a network of protected areas. A clear evaluation on how α, β, and γ diversity are partitioned among and within spatial (and temporal) scales can help us to drive manager decisions and provide methods for monitoring species diversity. Moving from these concepts, a probabilistic sample was applied here 1) for quantifying plant species diversity at different spatial scales within the network of protected areas existing in the Siena Province (Central Italy) and 2) to evaluate the effects on the species diversity of the whole network due to the addition of two new protected areas (Ripa d’Orcia and Bogatto). Focusing on both common species and those defined as “focal” (included in regional, national or continental “red” lists), our results indicated that species richness of the protected areas differed each other independently from the sampling efforts. Diversity partitioning indicated that most of the diversity within the network is due to larger-scale β-diversity, i.e., the differences in species composition among reserves. Moreover, total β-diversity was decomposed into two components: βarea (due to the differences in area among reserves) and βreplacement (due to the compositional differences across protected areas). βareawas particularly important for all species, while βreplacement was the most important factor for the set of focal species. Noteworthy, the analyzed variation in diversity components due to the inclusion of the two new natural reserves into the network (Ripa d’Orcia and Bogatto) resulted in a proportional increase in βarea for the whole network, and, on the other hand, in a reduction for βreplacement diversity component. Based on these quantitative results, criteria for the selection and the inclusion of protected areas in existing networks of natural reserves (such as the Natura 2000 Network) can be achieved.
Keywords
Additive Partitioning, Conservation biology, Focal species, Plant Diversity, Reserve selection, Siena Province
Introduzione
L’istituzione di reti di aree protette rappresenta, ad oggi, un mezzo indispensabile per la conservazione della biodiversità ([6], [7]). La selezione di nuove aree protette da destinare alla conservazione delle risorse naturali, richiede l’uso di strategie scientificamente valide, soprattutto per fornire una concreta protezione della biodiversità ed un efficace supporto agli interventi di gestione ([76]). A tale scopo, è quindi essenziale misurare il grado di diversità biologica associato alle diverse scale spaziali e temporali negli ecosistemi e, soprattutto, valutarne la variazione attraverso il monitoraggio.
Le misure di diversità specifica ricoprono un ruolo fondamentale sia nell’analisi ecologica, che nella biologia della conservazione e, di conseguenza, rappresentano misure indispensabili per un monitoraggio efficace ([74], [41], [52], [44], [50]). Classicamente, gli ecologi riconoscono tre differenti componenti della diversità specifica: α (diversità entro le comunità), β (diversità tra le comunità) e γ (diversità totale di una regione; [75]). La partizione additiva della diversità, formalizzata da Lande ([39]), rappresenta un metodo per combinare le tre componenti appena descritte ([18]). Attraverso questo metodo la diversità totale (γ) è semplicemente scomposta nel contributo dato dalla diversità specifica media alla scala di comunità (αmedio) sommato al contributo dovuto alla complementarietà specifica tra le comunità considerate (β); con l’approccio additivo tutte le componenti sono espresse nella stessa unità di misura, ovvero la ricchezza di specie ([45]). Inoltre, poiché la β-diversitàè espressa come numero di specie, può essere comparata anche tra scale spaziali e temporali diverse ([72], [21], [31], [63]). Tali proprietà rendono la partizione additiva della diversità un potente strumento operativo nei piani di monitoraggio, fornendo dati comprensibili e ripetibili sulla diversità specifica e sulla sua distribuzione spaziale ([31], [12], [13]).
Sebbene gli aspetti teorici dell’analisi partitiva della diversità siano stati ampiamente discussi ([39], [31]), tale metodo è stato applicato ai problemi ecologici o conservazionistici in maniera sporadica. Ad esempio, Allan ([3]) ha utilizzato la partizione della diversità per analizzare la distribuzione di insetti bentonici a diverse scale spaziali (da quella dei piccoli substrati rocciosi fino a quella che occorre lungo un gradiente altitudinale di 1000 m in un ruscello alpino in Colorado). In modo simile, Wagner et al. ([72]) descrivono le componenti della diversità vegetale alle diverse scale spaziali considerando alcuni ecosistemi agricoli. De Vries et al.[22], [23] - si veda anche [21]) hanno usato la partizione additiva per descrivere la distribuzione spaziale e temporale di lepidotteri nelle foreste pluviali. Infine, Gering et al. ([31]) hanno utilizzato questo metodo per indagare la distribuzione della biodiversità a varie scale spaziali nelle foreste temperate, al fine di sviluppare strategie di conservazione. In riferimento a reti di aree protette, Chiarucci et al. ([12], [13]) hanno applicato il metodo della partizione additiva alla valutazione delle componenti α, β e γ della diversità floristica nei Siti di Importanza Comunitaria (SIC) della Rete Natura 2000 (Direttiva Habitat 92/43/EEC) della Provincia di Siena. Considerando che le reti di aree protette vengono nel tempo implementate con l’aggiunta di nuove aree così come è successo per la rete delle riserve naturali della provincia di Siena, è possibile testare l’efficacia del metodo additivo come strumento di analisi della variazione delle componenti della diversità. In altri termini, attraverso il metodo della partizione additiva è possibile quantificare la variazione delle tre componenti α, β e γ della diversità floristica in seguito all’inserimento di nuove aree protette.
In questo studio il metodo della partizione additiva della diversitàè stato utilizzato come strumento semplice ed intuitivo per due scopi: (1) descrivere la partizione della diversità vegetale esistente nella rete di riserve naturali della provincia di Siena a differenti scale spaziali (da quella locale a quella di intera rete); e (2) valutare la variazione delle componenti di diversità della rete di riserve naturali della provincia di Siena in seguito all’aggiunta, avvenuta nel 2008, di due nuove riserve (“Ripa d’Orcia” e “Bogatto”). Tale analisi è stata sviluppata separatamente per la totalità delle specie vegetali e per le specie “focali”, ovvero le specie di particolare importanza inserite all’interno di liste rosse regionali, nazionali ed internazionali e per la cui conservazione è richiesta un’azione concreta.
Area di studio
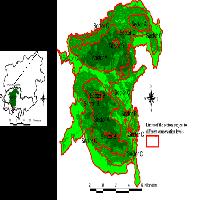
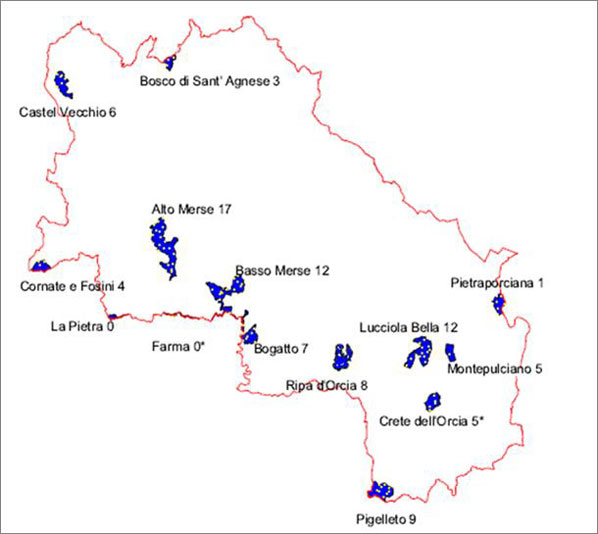
La rete di riserve naturali della provincia di Siena copre una superficie di 9661 ha (2.58% della superficie provinciale) ed è caratterizzata da aree di dimensioni medio - piccole (minimo 260 ha, massimo 1950 ha, Fig. 1). Undici delle quattordici riserve sono state istituite nell’anno 1996, mentre le riserve naturali “Ripa d’Orcia” e “Bogatto” sono state incluse nella rete nel 2008 (insieme alla riserva naturale “Crete dell’Orcia”, non considerata in questo studio). Le caratteristiche ambientali delle riserve sono molto eterogenee. Dal punto di vista geolitologico sono presenti principalmente terreni argillosi ed arenacei con affioramenti di rocce carbonatiche cavernose e di rocce ofiolitiche. La variabilità altitudinale va da un minimo di 71 m (Basso Merse) ad un massimo di 913 m (Pigelleto), mentre la piovosità media annua va da 650-700 mm (Lucciola Bella) a 1100 mm (Alto Merse).
Fig. 1 - Area di studio: sono raffigurati i confini della provincia di Siena, le riserve provinciali (in blu) e il numero di unità campionarie per riserva. (*): Riserve non incluse in questo studio.
Le fisionomie vegetazionali maggiormente rappresentate sono schematicamente descritte in Tab. 1.
Tab. 1 - Caratteristiche delle 14 riserve provinciali (e della riserva statale di Tocchi) considerate nel piano di campionamento sviluppato. È indicata l’area in ettari, il numero di plot campionati e il tipo di vegetazione prevalente per ogni riserva. (*): Area priva di punti campionari; (**): riserva non campionata.
| Riserva | Area (ha) | N° plot | Tipo di vegetazione prevalente |
|---|---|---|---|
| Pigelleto | 872 | 9 | Abetine con Fagus sylvatica e foreste di Q. cerris |
| Alto Merse | 1952 | 17 | Boschi sempreverdi (Q. ilex) - Pinus pinaster |
| Tocchi | 575 | 2 | Piantagioni di Pinus pinaster |
| Basso Merse | 1418 | 12 | Macchia Mediterranea |
| Pietraporciana | 336 | 1 | Boschi di Faggio |
| Lucciola Bella |
1183 | 10 | Vegetazione di biancana -Coltivi |
| Cornate e Fosini | 393 | 3 | Boschi di Q. ilex, Q.cerris e Q.pubescens |
| Bogatto | 588 | 7 | Lecceta, vigneti |
| Lago di Montepulciano | 455 | 5 | Coltivi, vegetazione acquatica, praterie palustri |
| Bosco di Sant’Agnese | 262 | 3 | Cupressus sempervirens |
| Ripa d’Orcia | 839 | 9 | Macchia Mediterranea - Boschi sempreverdi Q. ilex |
| Castelvecchio | 628 | 6 | Boschi di Faggio - Boschi dominati da Q. cerris |
| La Pietra* | 102 | 0 | Boschi misti dominati da Q. cerris |
| Farma** | 86 | 1 | Boschi dominati da Q. cerris - Boschi di Castanea sativa |
| Crete dell’Orcia** | 963 | 5 | Coltivi - Vegetazione di Biancana |
| Totale | 9691 | 90 | - |
Materiali e metodi
Disegno campionario e raccolta dati
L’applicazione di campionamenti di tipo probabilistico, come quello utilizzato in questo studio, risponde alla necessità di rispettare criteri di oggettività nella raccolta dati, permettendo, inoltre, di valutare l’accuratezza delle stime di interesse, anche per dati relativi alla diversità biologica ([76], [29]).
Il disegno campionario utilizzato in questo studio si basa sullo schema impiegato per l’Inventario Nazionale delle Foreste e dei Serbatoi del Carbonio (INFC - [27]). Le unità campionarie sono state selezionate in modo casuale all’interno delle maglie di una griglia (disegno casuale ristretto, [26]), in numero di una per ciascuna maglia, composta da quadrati contigui di 1 km2, non sovrapposti, che si estende su tutto il territorio nazionale. Le unità campionarie INFC ricadenti all’interno delle riserve naturali presenti in Provincia di Siena sono state selezionate per i rilievi vegetazionali utilizzati nel presente studio, per un totale di 90 unità campionarie riferite all’intera rete di riserve naturali (intensità di campionamento nominale di 1 unità per km2 di superficie di ogni riserva). La diversità vegetale è stata rilevata in tutti i punti durante il corso di più anni (dal 2005 al 2008). Ad oggi, per sei punti (riferibili a 2 riserve naturali - Val di Farma e Crete dell’Orcia) non sono ancora disponibili i dati sulla diversità floristica campionata; ne risulta che il data set totale considerato in questo lavoro risulta costituito da 84 unità campionarie (Tab. 1).
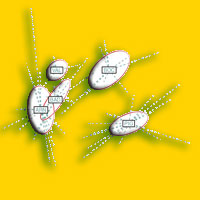
I dati a terra sono stati raccolti utilizzando un’unità di campionamento composta da un quadrato di 10 m di lato (plot), centrato nel punto di campionamento sopra descritto. Il plot è stato suddiviso in 4 quadranti (5x5 m) e ogni quadrante ulteriormente diviso in 4 subplot con lato pari a 2.5 m (Fig. 2). In ciascun subplot sono state annotate tutte le entità (specie o sottospecie, nel seguito dell’articolo riportate per semplicità come entità specifiche) di piante vascolari presenti (direttamente in campo ove possibile, o mediante raccolta di campioni e successiva determinazione in laboratorio). Per la determinazione dei campioni sono stati utilizzati principalmente, Pignatti ([53]), in minor misura Tutin et al. ([65], [66]) e Eggenberg & Moehl ([25]) per gli esemplari sterili). Successivamente alla determinazione, sotto il profilo nomenclaturale, i nomi dei taxa sono stati adeguati a quelli della checklist della Flora Italiana ([16]); sotto il profilo tassonomico, però, non sempre è stato possibile spingere la determinazione fino al livello di sottospecie, cui la checklist fa ampiamente ricorso: pertanto, i nomi delle specie da noi utilizzati corrispondono generalmente al taxon indicato dalla checklist come “s.l.” e non necessariamente alla sottospecie nominale. Per tutte le specie campionate, è stata successivamente verificata la loro appartenenza a liste di interesse internazionale (Allegati II, IV, V della Direttiva “Habitat” e “IUCN Red List of Threatned Species”, [36]), nazionale (Lista Rossa delle Piante d’Italia, nella versione aggiornata allegata alle liste rosse regionali in [15]) o regionale (lista RE.NA.TO - [1] - e lista delle specie di Interesse Regionale - L.R. 56/00 - [57]). Le specie appartenenti a tali liste sono state denominate “specie focali”. Sebbene le liste rosse nazionali e regionali disponibili per l’Italia presentino alcune criticità metodologiche e problemi di aggiornamento ([62]), si ritiene che ai fini di questo studio esse rappresentino comunque una fonte adeguata per testare sotto il profilo metodologico e concettuale il rapporto tra specie “di pregio” e biodiversità complessiva. In questo studio è stata analizzata la distribuzione alle diverse scale spaziali delle specie focali, la cui influenza sui modelli distributivi di ricchezza specifica rimane ad oggi ancora poco esplorata ([40], [5]).
Fig. 2 - Schema del disegno campionario utilizzato per la raccolta dei dati floristici nel sistema di riserve della Provincia di Siena. All’interno di ciascuna cella di 1x1 km della griglia INFC è stato selezionato un punto casuale. Tutti i punti ricadenti all’interno delle riserve provinciali sono stati campionati utilizzando un’unità campionaria (plot) di 10x10 m (suddiviso in 16 settori di 2.5x2.5m).

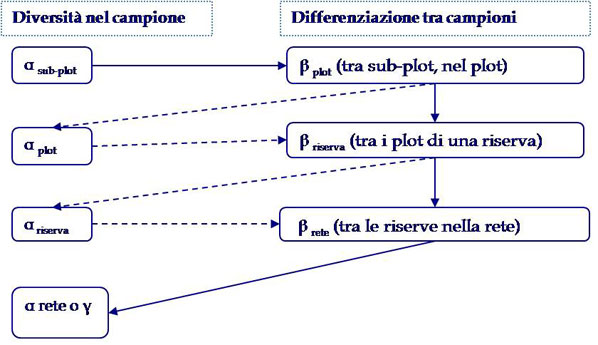
Nel complesso, la diversità vegetale è stata analizzata a quattro diverse scale spaziali, organizzate gerarchicamente: subplot, plot, riserva naturale e, infine, intera rete di aree protette (Fig. 3).
Fig. 3 - Relazioni additive tra le diverse componenti della diversità per tutte le scale spaziali di indagine analizzate.
Tutte le analisi sono state sviluppate sia per le specie totali (tutte le specie rilevate) che per le sole specie focali (le specie incluse in una delle sopra citate liste di interesse).
Analisi della diversità tra riserve naturali
Per confrontare la ricchezza di specie tra siti campionati con intensità differente sono state utilizzate le curve di rarefazione basate su campioni, calcolate sia in funzione dell’accumulo di plot che di intere riserve naturali. In generale, la curva di rarefazione esprime il numero atteso di specie in un piccolo gruppo di n individui o n campioni, preso a caso dal numero N totale degli individui o campioni rilevati ([37]); può essere calcolata sulla media di un numero sufficiente di curve di accumulazione ([33]) o attraverso l’uso di una formula analitica ([67], [38], [13] - eqn. 1):
dove E(Sn) è il numero atteso di specie per il sottocampione di dimensione n, S è il numero di specie totale osservato, N è il numero totale di campioni mentre Ni è il numero di campioni in cui è presente l’i-esima specie. In questo caso per numero di campioni si intende il numero di plot o di riserve naturali analizzate.
Per indagare il pattern di diversità delle specie focali in relazione alla ricchezza specifica totale campionata, è stato calcolato il rapporto tra i valori ottenuti dalle due curve di rarefazione (specie focali/specie comuni - [12]).
Partizione della diversità vegetale nella rete di riserve naturali della provincia di Siena
La diversità presente nell’intera rete di riserve naturali è stata ripartita nelle componenti a scala di subplot (αsubplot), che sommata alla complementarietà specifica tra sub-plot all’interno del medesimo plot (βplot) fornisce la diversità totale di ciascuno di essi (αplot); quest’ultima componente, sommata con la complementarità media tra i plot all’interno della stessa riserva (βriserva), risulta nella diversità specifica di ciascuna riserva (αriserva); infine, l’αriserva, sommata alla complementarietà tra riserve della rete (βrete - Fig. 3), risulta nella diversità specifica totale del sistema di riserve naturali provinciali, αrete o γ-diversità totale. In questo modo, la diversità totale può essere scomposta nella somma proporzionale della diversità individuata ad ogni livello del disegno di campionamento gerarchico (Fig. 3, per i dettagli matematici si veda [12]).
Un’ulteriore indagine sulla scomposizione della diversità nella rete di aree protette è stata effettuata prendendo in considerazione la variazione delle componenti analizzate alle varie scale spaziali prima e dopo l’introduzione di due nuove riserve (Ripa d’Orcia e Bogatto).
In questo modo è stato possibile verificare quali componenti siano proporzionalmente aumentate o diminuite in seguito all’incremento delle riserve naturali nella rete preesistente.
Infine, è stato possibile effettuare un’ulteriore scomposizione della β diversità (calcolata come in precedenza) nelle due componenti βarea e βreplacement; la prima componente dipende dalla differenza in estensione tra le aree paragonate (ossia dalle differenze in superficie tra le singole riserve naturali), mentre la seconda componente dipende dalla differenza nella loro composizione in specie (intesa come un surrogato dell’eterogeneità ambientale, si veda [17], [12]). Quindi, la βrete è stata scomposta nelle componenti βarea e βreplacement utilizzando le eqn. 2, eqn. 3, e eqn. 4:
Il livello di significatività delle componenti della diversità specifica (α e β) alle varie scale spaziali è stato testato con il software Partition 2.0 ([70]): mediante questo software, vengono effettuati dei classici Monte Carlo test che prevedono la randomizzazione della matrice di occorrenza specifica di un certo livello spaziale rispetto al livello spaziale superiore (ad esempio, si testa la significatività della componente α a scala di subplot randomizzando le specie presenti nei subplot rispetto al plot in cui queste sono state rilevate, per i dettagli si faccia riferimento a [18]).
Per il calcolo delle curve di rarefazione è stato utilizzato il software R ([55]).
Risultati
Diversità floristica campionata
Negli 84 plot rilevati, la ricchezza specifica totale campionata è pari a 529 specie. In tale numero non sono stati considerati i campioni identificati solo a livello di genere e inoltre sono state escluse due specie annuali coltivate, Solanum lycopersicon ed Ocimum basilicum. Al contrario, altre specie coltivate (Triticum) sono state inserite nel data set in quanto caratterizzanti l’uso del suolo nell’area di studio e dominanti le comunità vegetali esaminate. La ricchezza specifica media riscontrata a scala di plot è risultata di 28.6 specie, con una variabilità compresa tra 1 e 81 (Tab. 2). Le specie più frequenti (relativamente ai plot rilevati e al numero di frequenze all’interno del plot) sono risultate essere Fraxinus ornus, Quercus ilex, Q. cerris e Arbutus unedo tra le specie arboree ed arbustive, Brachypodium rupestre, Carex flacca e Rubia peregrina per quelle erbacee e Hedera helix per le specie lianose.
Tab. 2 - Media, minimo e massimo della ricchezza specifica a scala di plot e di riserva, valori dell’indice di Shannon e di Pielou per riserva. μ: media; DS: deviazione standard.
| Riserva | Totalità delle specie | Specie focali | ||||
|---|---|---|---|---|---|---|
| N° Specie/ plot (μ ± DS) |
Min-Max | N °Specie/ Riserva |
N° Specie/ plot (μ ± DS) |
Min-Max | N° Specie/ Riserva |
|
| Pigelleto | 37.3 ± 12.9 | 16 - 59 | 145 | 2.2 ± 0.8 | 1 - 4 | 7 |
| Alto Merse | 27.9 ± 18.1 | 6 -74 | 202 | 1.7 ± 0.9 | 1 - 4 | 13 |
| Tocchi | 39 ± 33.9 | 15 - 63 | 73 | 1.0 ± 0 | 1 - 1 | 2 |
| Basso Merse | 23.7 ± 13.8 | 7 - 55 | 157 | 0.7 ± 1 | 0 - 3 | 7 |
| Pietraporciana | 27.0 ± 0 | 27 - 27 | 27 | 2.0 ± 0 | 2 - 2 | 2 |
| Lucciola Bella | 25.9 ± 12.5 | 10 - 38 | 133 | 0.2 ± 0.4 | 0 - 5 | 11 |
| Cornate e Fosini | 37.8 ± 6.7 | 23 - 37 | 97 | 0 ± 0 | 0 | 0 |
| Bogatto | 21.4 ± 12.7 | 10 - 48 | 80 | 0.3 ± 0.8 | 0 - 2 | 2 |
| Lago di Montepulciano | 4.0 ± 3.6 | 1 - 9 | 19 | 0.2 ± 0.4 | 0 - 1 | 1 |
| Bosco di Sant’Agnese | 27 ± 13.2 | 17 - 42 | 71 | 0.7 ± 1.2 | 0 - 2 | 2 |
| Ripa d’ Orcia | 29.9 ± 28.5 | 1 - 81 | 131 | 1.5 ± 1.7 | 0 - 5 | 8 |
| Castelvecchio | 42.8 ± 17.2 | 12 - 62 | 132 | 2 ± 1.3 | 1 - 4 | 6 |
| Intera Rete | 28.6 ± 14.4 | 1 - 81 | 529 | 1 ± 0.8 | 0 - 5 | 40 |
In totale, le specie focali campionate sono risultate 40 con una media di 1.0 per plot (Tab. 2). Tra queste, la più abbondante è risultata Ruscus aculaeatus.
L’intervallo di variabilità del numero di specie rilevato a scala di riserva varia da 202 specie (Alto Merse), a 19 (riserva Lago di Montepulciano). Diverso è il modello di distribuzione delle specie focali, con un massimo di 13 (Alto Merse) ed un minimo di 0 specie (Cornate e Fosini).
Analisi comparativa della diversità tra riserve
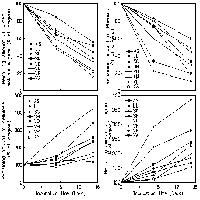
L’andamento delle curve di rarefazione calcolate a scala di plot è mostrato in Fig. 4a: tutte le curve sono caratterizzate da una crescita pressoché continua, con le eccezioni delle riserve Lago di Montepulciano e Bogatto, che presentano una curva con pendenza molto ridotta e comportamento quasi asintotico. È da osservare che le curve di Tocchi, Pigelleto, Castelvecchio e Cornate e Fosini hanno un’elevata diversità specifica media a scala di plot e crescono con una pendenza molto elevata, superiore alla curva di rarefazione costruita aggregando casualmente tutti i plot. Questo a sottolineare non solo (i) l’alto numero di specie a scala locale, ma anche (ii) un’elevata complementarietà specifica tra plot all’interno di queste riserve.
Fig. 4 - Curve di rarefazione: a) Curve di rarefazione plot-based costruite sull’intero insieme di specie per ogni riserva; b) Curve di rarefazione plot-based per le sole specie focali (analisi effettuata per quelle riserve con almeno una specie focale). Gli assi di tutti i grafici sono mostrati fino ad un numero di 20 plot. Il numero di specie totali è 529. Le focali sono 40. Il numero di plot totali è 84.
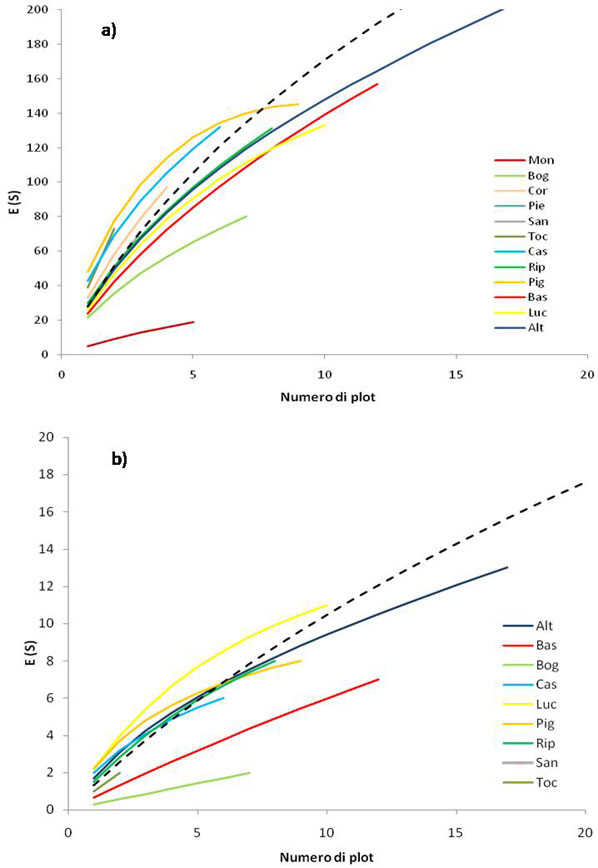
Per le specie focali è evidente un’alta variabilità nel pattern delle curve di rarefazione (Fig. 4b). In particolare, la curva di Lucciola Bella mostra un’elevata pendenza e quindi una elevata complementarità specifica tra i plot. Un simile andamento caratterizza anche Pigelleto e Castelvecchio. La curva con pendenza minore è, al contrario, quella per la riserva Bogatto.
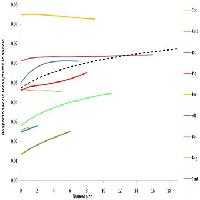
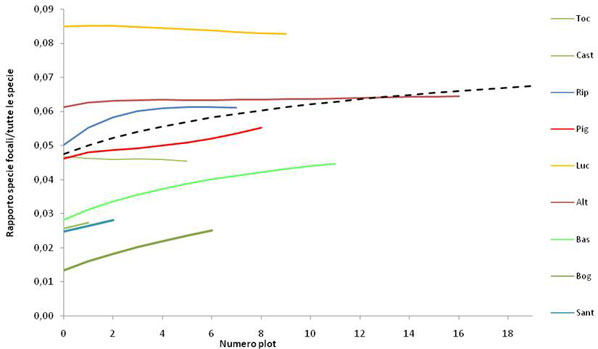
Il rapporto tra la curva di rarefazione delle specie focali rispetto a quella costruita per tutte le specie campionate (Fig. 5), mostra un andamento crescente all’aumentare del numero di plot cumulati. Osservato a scala di singola riserva, tale andamento varia ampiamente.
Fig. 5 - Rapporto tra curve di rarefazione (numero di specie focali atteso/ricchezza specifica totale attesa) per numero di plot cumulati. È mostrata anche la curva per il rapporto calcolato aggregando tutti i plot. Da notare che sono state riportate solo le riserve che presentavano più di un plot con una specie rara presente (sono escluse le riserve di Pietraporciana, Montepulciano e Cornate e Fosini).
Partizione della diversità vegetale nella rete di riserve naturali della provincia di Siena
La maggior parte della diversità specifica riscontrata nel campione, 423.5 specie (sulle 529 campionate), è imputabile alla βrete (Fig. 6a e Tab. 3) e tale dato non è il risultato di un assemblaggio casuale di specie (p = 0.09). Osservando nel dettaglio la scomposizione della βrete nelle sue due componenti principali, si osserva (Tab. 5) che la componente βarea ha un peso molto maggiore (circa 327 specie sulle 423.5 della βrete) rispetto alla componente βreplacement (96.5 specie sulle 423.5 della βrete). Alle scale spaziali inferiori, la diversità media di ogni riserva (αriserva) corrisponde a 105.5 specie (21.2 % del totale), mentre quella dovuta alla β-diversità tra i plot all’interno delle riserve (βriserva) corrisponde a 77.6 specie (15.5% - Tab. 3).
Fig. 6 - Partizione additiva della diversità per l’intera rete di riserve naturali: a) rispetto al numero totale di specie rilevate e b) per le sole specie focali.
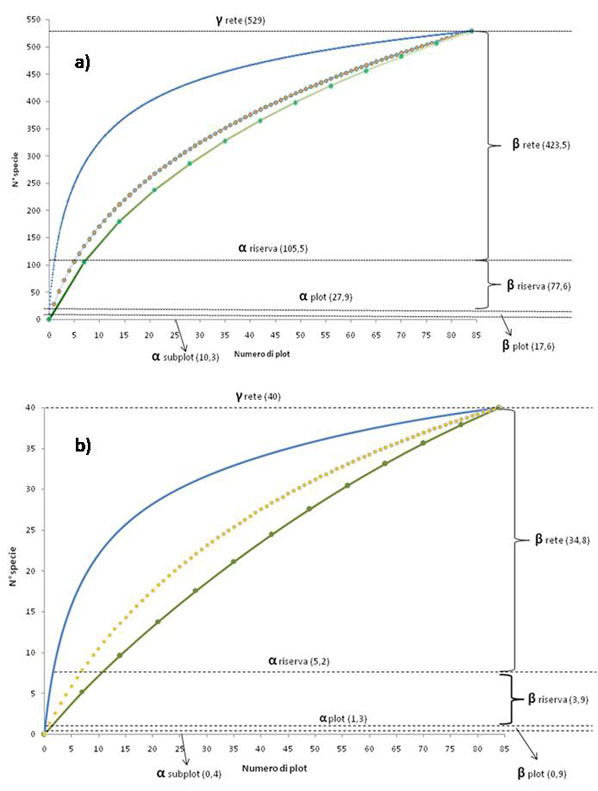
Tab. 3 - Partizione additiva della diversità di tutte le specie con relativa significatività e contributo percentuale alla diversità totale delle varie scale analizzate.
| Diversità (N° di specie) | Diversità (%) | |||||
|---|---|---|---|---|---|---|
| Scala | Prima | Dopo | Prima | Dopo | ||
| S | Significatività (p-value) |
S | Significatività (p-value) |
|||
| βrete | 390.5 | 0.27 | 423.5 | 0.09 | 78.7 | 80.1 |
| αriserva | 105.5 | 0.27 | 105.5 | 0.1 | 21.3 | 19.9 |
| βriserva | 77.2 | 0 | 77.6 | 0 | 15.6 | 14.7 |
| αplot | 28.3 | 0 | 27.9 | 0 | 5.7 | 5.3 |
| βplot | 18 | 0 | 17.6 | 0 | 3.6 | 3.3 |
| αsubplot | 10.3 | 0 | 10.3 | 0 | 2.1 | 1.9 |
Tab. 5 - Partizione della componente βrete in βarea e βreplacement. prima e dopo l’introduzione delle due nuove riserve. per tutte le specie rilevate e per il solo gruppo di specie definite come focali (ciascuna componente è espressa come numero assoluto di specie ma anche come percentuale. in modo da rendere le componenti della diversità direttamente paragonabili nel tempo).
| Parametro | Componente | Ricchezza Specifica Totale |
Ricchezza Specifica Specie Focali | ||
|---|---|---|---|---|---|
| Prima | Dopo | Prima | Dopo | ||
| N° | βarea | 294 | 327 | 23 | 27 |
| βreplacement | 96.5 | 96.5 | 7.8 | 7.8 | |
| % | βarea | 59.3 | 61.8 | 63.9 | 67.5 |
| βreplacement | 19.5 | 18.2 | 21.7 | 19.5 | |
La scomposizione additiva delle specie focali ricalca parzialmente quella appena descritta per tutte le specie (Fig. 6b, Tab. 4), ma diventa ancora più importante il peso della differenziazione tra riserve (βrete), che raggiunge l’87 % del totale. Inoltre, l’ulteriore partizione della βrete ha dimostrato come, anche per questo gruppo di specie, gran parte di questa sia dovuta alla sua componente βarea (circa 27 specie delle 35 relative alla βrete).
Tab. 4 - Partizione additiva della diversità delle specie focali con relativa significatività e contributo percentuale alla diversità totale delle varie scale analizzate.
| Diversità (N° di specie) | Diversità (%) | |||||
|---|---|---|---|---|---|---|
| Scala | Prima | Dopo | Prima | Dopo | ||
| n° | Significatività (p-value) |
n° | Significatività (p-value) |
|||
| βrete | 30.8 | 0.15 | 34.8 | 0.06 | 85.5 | 87 |
| αriserva | 5.2 | 0.92 | 5.2 | 0.82 | 14.5 | 13 |
| βriserva | 3.8 | 0 | 3.9 | 0 | 10.6 | 9.7 |
| αplot | 1.4 | 0 | 1.3 | 0 | 3.9 | 3.3 |
| βplot | 0.9 | 0 | 0.9 | 0 | 2.5 | 2.3 |
| αsubplot | 0.5 | 0 | 0.4 | 0 | 1.4 | 1 |
Effetto dell’aggiunta di due nuove riserve sulla diversità vegetale dell’intera rete provinciale
Confrontando i valori delle componenti spaziali della diversità rispetto all’introduzione delle nuove riserve naturali (Ripa d’Orcia e Bogatto), si osserva un generale incremento della βrete (che passa dal 78.7% all’80.1%) a discapito di tutte le altre componenti (Tab. 3). Nel dettaglio, l’incremento della βrete sembra imputabile all’effettivo incremento dell’area totale della rete (la componente βarea passa infatti dal 59.3% al 61.8%) a cui però corrisponde una lieve diminuzione proporzionale della componente βreplacement (Tab. 5). Considerando le sole specie focali, il valore della βrete risulta incrementato con l’introduzione delle nuove riserve naturali (βrete varia infatti dall’85.5% all’87%) e anche in questo caso l’incremento osservato può essere attribuito al solo aumento in superficie dell’intera rete di riserve naturali (βarea passa proporzionalmente dal 63.9% al 67.5%, Tab. 5).
Discussione
Diversità floristica campionata
A scala di plot, sono state riscontrate 28.6 specie. Il plot con maggior numero di specie (81) è stato campionato nella riserva di Ripa d’Orcia in un’area di margine tra bosco dominato da Quercus ilex e un’area a vegetazione aperta e disturbata a dominanza di Elymus repens. All’opposto, sono tre i plot con ricchezza di specie uguale ad una sola specie, costituiti da un coltivo di Triticum turgidum subsp. durum (Lucciola Bella) e da due plot ricadenti in acqua con Phragmithes australis e Utricularia vulgaris (Lago di Montepulciano). Anche la ricchezza specifica a scala di riserva è molto variabile e dipendente soprattutto dall’estensione dell’area campionata: Alto Merse, infatti, conta il maggior numero di specie (202) e il maggior numero di specie focali (13 - Tab. 3). Tale andamento rispecchia le classiche osservazioni sulla dipendenza esistente tra ricchezza di specie e area (curve specie-area, [4], [32], [61]). Due sono le principali cause di tale andamento ([73]): (i) l’effetto dell’“Area per sé”, secondo il quale, dato un habitat uniforme, una porzione piccola di spazio può supportare solo un numero limitato di individui e, conseguentemente, anche di specie; (ii) effetto “Habitat diversity”, per il quale, all’aumentare dell’area, aumenta anche la variabilità dell’habitat e il numero di nicchie ecologiche potenzialmente disponibili per specie con esigenze ecologiche diverse.
La maggiore frequenza nella rete di riserve di specie arboree quali Fraxinus ornus, Quercus ilex, Q. cerris ed Arbutus unedo è collegata a una vegetazione dominata da macchia mediterranea e boschi termofili misti. Molto frequenti sono Hedera helix tra le specie lianose, e Brachypodium rupestre tra le specie erbacee. L’alta frequenza di H. helix è legata alla larghissima ampiezza ecologica di questa specie che, pur con un baricentro suboceanico e mesofilo, è molto diffusa in Italia in tutti gli ambienti forestali dal livello del mare fino a più di 1200 m ([53], [56]); B. rupestre è specie anch’essa ad ecologia piuttosto ampia, leggermente basifila e mesoxerofila, particolarmente competitiva su substrati calcarei e argillosi; spesso avvantaggiata da fenomeni di degradazione della copertura vegetale e nell’Italia submediterranea è frequente sia in comunità prative che all’interno di boschi decidui termofili ([53], [56], [2]). Tra le specie focali, la più frequente è Ruscus aculeatus, che è stata rinvenuta un elevato numero di volte (110). Questa specie, pur essendo elencata tra le specie di Interesse Comunitario a causa dell’eccessiva raccolta (Direttiva 92/43/CEE, All. V - Specie animali e vegetali di interesse comunitario il cui prelievo in natura e il cui sfruttamento potrebbero essere oggetto di misure di gestione) è piuttosto comune in Toscana e in Italia, a causa della sua larga valenza ecologica (con un carattere mediterraneo-atlantico); è frequente nelle leccete e nei boschi caducifogli termofili ([53], [56]), e spesso si avvantaggia della degradazione del sottobosco per sovrappascolo o altro tipo di disturbo ([30]).
Analisi comparativa della diversità tra riserve
Ricercare i modelli di distribuzione della diversità attraverso la costruzione di curve di rarefazione basate su campioni è una strategia, ad oggi, largamente utilizzata. Queste curve sono state utilizzate per interpretare la diversità vegetale a scala regionale in Svizzera ([38]) e per quantificare la diversità floristica a diverse scale spaziali nelle foreste della Toscana ([8]). Il loro utilizzo in questo lavoro, oltre che per testare l’efficienza del campionamento ([28]), è stato quello di rendere confrontabili aree con estensioni molto differenti. Le curve di rarefazione sviluppate presentano un andamento diversificato tra le singole riserve e per i due gruppi di specie considerate. La curva con crescita maggiore, considerando l’insieme delle specie rilevate, è quella ottenuta per la riserva Pigelleto, ciò ad esprimere sia un’elevata ricchezza di specie, che un’alta complementarità nella composizione specifica tra i plot di questa riserva. La riserva naturale del Pigelleto, che ospita formazioni forestali a dominanza di Fagus sylvatica, Abies alba e anche Quercus cerris, comprende un’elevata eterogeneità ambientale e presenta valori elevati di diversità specifica. Le curve relative alle riserve naturali di Castelvecchio, Cornate e Tocchi hanno un andamento simile a Pigelleto, mentre le due riserve più estese, Alto Merse e Basso Merse, presentano curve con una pendenza al di sotto della media. Interessante è notare come le riserve Ripa d’Orcia e Bogatto, siano entrambe poco ricche in specie e presentino una bassa complementarietà tra i plot al loro interno, presentando nel complesso, ambienti molto omogenei da un punto di vista vegetazionale.
Relativamente alle specie focali, è interessante l’andamento della curva di rarefazione per Lucciola Bella, con pendenza molto elevata e costantemente al di sopra della curva di rarefazione media. Tale pattern non è comune, poiché ci si attende che la curva calcolata aggregando tutti i plot sia quella a pendenza maggiore. Questo perché l’insieme di tutti i plot dovrebbe essere più eterogeneo rispetto ai dati provenienti da un gruppo di campioni uniti da rapporti spaziali più stretti ([67], [68], [8], [14]). Per queste ragioni Lucciola Bella risulta una riserva di estrema importanza che raccoglie un elevatissimo numero di specie focali, ospitate nei diversi habitat che la caratterizzano. Qui, infatti, la ricchezza di specie focali è in parte dovuta alla presenza di formazioni geomorfologiche tipiche che contraddistinguono la riserva anche da un punto di vista paesaggistico: le biancane. Specie come Parapholis strigosa e Artemisia caerulescens subsp. cretacea sono proprie dei terreni argillosi sub-salsi in erosione, mentre la presenza di Hordeum marinum, Allium paniculatum e Consolida regalis è attribuibile alle particolari condizioni climatiche e alla presenza di aree riservate a coltivazioni con tecniche tradizionali ([9], [42], [43]). Inoltre, queste considerazioni confermano l’importanza degli habitat semi-naturali come serbatoi di biodiversità e, dunque, la necessità della loro presenza all’interno di reti di aree protette (come richiesto anche dalla stessa Direttiva Habitat 92/43/CEE).
È interessante notare che la quasi totalità delle curve di rarefazione calcolate (sia per tutte le specie che per le specie focali) non presentano un andamento asintotico. Tale dato può essere spiegato con il fatto che, nonostante sia stato campionato un elevato numero di plot, questo non è stato sufficiente a campionare tutta la diversità specifica presente nelle riserve ([33]), data l’alta eterogeneità ambientale dell’ area di studio indagata ([12], [13]).
Studi recenti ([51]) riportano che i pattern di distribuzione della ricchezza di specie possono riflettere in qualche modo quelli di differenti gruppi di specie (come ad esempio quelle rare): ciò dovrebbe risultare in un andamento monotonamente crescente della ricchezza di specie focali all’aumentare della ricchezza di specie campionate ([40]). Tuttavia, nel presente studio, le specie focali mostrano curve con modelli di crescita molto diversi rispetto al numero totale di specie censite. Questo suggerisce che: (i) la distribuzione delle specie focali (specie rare, endemiche o minacciate) può in realtà essere diversa dalla distribuzione delle specie comuni, come sottolineato anche in altri studi sui modelli di ricchezza di uccelli svolti in Gran Bretagna ed Africa ([40]); (ii) le specie comuni non rappresentano buoni predittori dei pattern di ricchezza dell’insieme delle specie ([12]). Il rapporto tra l’andamento delle specie focali e quello di tutte le specie campionate aumenta con l’aumentare del numero di plot, testimoniando come, all’interno della rete di riserve studiate, da un punto di vista generale, le specie focali riscontrate aumentino rispetto alle specie comuni, all’aumentare dello sforzo campionario (quantificato come numero di plot campionati). Tale andamento risulta legato al fatto che queste specie sono, in media, meno frequenti della totalità delle specie ([12]). Per questo motivo, le specie focali risultano sotto campionate durante un campionamento probabilistico disegnato per avere una rappresentatività dell’intero territorio e, quindi, non appositamente progettato per rilevare con maggiore probabilità gli habitat più rari.
Partizione additiva della diversità per la rete di riserve naturali
L’importanza del ruolo della complementarietà composizionale tra siti, nel determinare la diversità a larga scala, è stato sottolineato già da Chiarucci et al. ([12]), che indagando il ruolo della diversità vegetale nel network di SIC della Rete Natura 2000 della provincia di Siena, evidenziano come la maggior parte della diversità floristica sia dovuta alla complementarietà specifica (β-diversità) tra i singoli SIC che compongono la rete (in altri termini, all’unicità della composizione floristica che caratterizza i diversi SIC).
Similmente, la partizione additiva della diversità di coleotteri arboricoli nelle foreste decidue dell’Ohio e Indiana ha mostrato come la β-diversità entro i siti di campionamento e quella tra i siti entro la medesima eco-regione siano le componenti che contribuiscono in misura maggiore alla diversità totale ([31]). Veech & Crist ([71]) evidenziano come normalmente la β-diversità a livello di paesaggio contribuisce per il 50% o più alla γ-diversità totale e sia correlata positivamente e in maniera significativa all’eterogeneità ecosistemica e climatica. In accordo con questi lavori, anche la diversità nell’intero sistema di riserve naturali della Provincia di Siena è dovuta (per l’80% della diversità totale) alle differenze nella composizione specifica che intercorrono tra le riserve (βrete). L’alta percentuale che la βrete fornisce alla diversità floristica totale non è, dunque, il risultato di un assemblaggio casuale di specie ma è attribuibile ad una serie di fattori ambientali e antropici specifici che hanno creato una grande diversità di vegetazione e di nicchie ecologiche. Tale effetto è dovuto alle condizioni diversificate in termini di geologia, topografia, uso del suolo e ad una gestione forestale perpetrata da lungo tempo ([20]). L’utilizzo millenario di queste aree per scopi agricoli, pastorali e selvicolturali, ha prodotto un mosaico complesso, composto da poche aree naturali residue, miste a molti habitat semi-naturali e ad una ricca varietà di tipi di vegetazione secondaria ([20]).
La partizione additiva della diversità delle specie focali rivela che i loro modelli di distribuzione ricalcano solo parzialmente i modelli di ricchezza di tutte le specie campionate. Infatti, per questo gruppo di specie si assiste ad una maggiore diversità dovuta alla differenziazione delle riserve (87% della diversità totale - Tab. 4), che consegue in un contributo percentuale minore delle restanti componenti alla diversità totale. Per questo insieme di specie, l’αsubplot è la componente che ha in proporzione la maggiore riduzione (rispetto alla totalità delle specie). L’estensione molto piccola e la relativa omogeneità delle condizioni ambientali nei subplot campionati all’interno dello stesso plot influiscono sulla presenza delle specie focali, che, alla scala spaziale esaminata, non coesistono. Ciò viene spiegato considerando l’autoecologia delle specie focali, quasi tutte con particolari esigenze ecologiche (edafiche, climatiche, ecc.), e che difficilmente sono rappresentabili in una stessa area contemporaneamente, e per tale motivo risultano spazialmente isolate ([35]).Verosimilmente questo è il motivo per cui le specie focali presentano una diversità maggiore del 7% rispetto all’insieme di specie campionate, alla scala di analisi più estesa (βrete).
Effetto dell’ampliamento della rete di riserve provinciali
La creazione di un sistema di aree protette è una necessità stabilita dalle Convenzioni sulla Diversità Biologica, di Ramsar, di Berna ed è la loro istituzione è obbligatoria per tutti i membri dell’Unione Europea (attuata dalle Direttive Habitat ed Uccelli - [59]). Il primo obiettivo nella creazione di un sistema di aree protette è conservare nel lungo termine e per una data regione, la diversità delle sue caratteristiche biologiche ([60]). La selezione delle riserve basata sul principio di complementarietà ([69]) presuppone che le risorse per la protezione della biodiversità saranno sempre limitate. Infatti, attraverso questo principio, si cerca di ottenere la massima efficienza (sensu [54]) nella rappresentazione della biodiversità con il minimo costo.
La procedura con cui si può soddisfare tale principio è attraverso il calcolo dell’area minima in cui tutte le caratteristiche della diversità biologica sono rappresentate almeno una volta ([47], [64], [19], [34]). Tale procedura sottintende che, se una specie è contenuta nelle riserve, può essere in qualche modo protetta (anche se è stato dimostrato che questo approccio non è sempre sufficiente ad assicurare una persistenza a lungo termine della diversità nel sistema, [60]).
La rete di riserve della provincia di Siena è caratterizzata da avere aree protette di dimensioni medio - piccole e di racchiudere in se un’alta diversità di ambienti. I criteri di selezione rispecchiano, quindi, i risultati dei recenti studi di Zhou & Wang ([77]), che cercando la migliore configurazione di habitat per la persistenza delle meta-popolazioni in un sistema di aree protette naturali, individuano la soluzione ottimale in una rete di molte aree di dimensioni relativamente piccole, piuttosto che poche aree grandi o molte aree molto piccole.
Nel 2008, con delibera del consiglio provinciale di Siena, sono state istituite tre nuove riserve entranti a far parte della rete di riserve naturali (Ripa d’Orcia, Bogatto e Crete dell’Orcia), tutte ricadenti nella Val d’Orcia. Le due nuove riserve naturali considerate in questo studio (si ricorda che i dati vegetazionali non sono al momento disponibili per la riserva “Crete dell’Orcia”) hanno mostrato, da una prima analisi, curve di rarefazione e valori di βplot e βriserva inferiore rispetto alle riserve naturali già presenti nella rete. La riserva naturale Bogatto è caratterizzata per oltre il 50% da macchia mediterranea e cedui di leccio, per il 35% da boschi misti di caducifoglie e per il restante 15% da coltivi e vigneti ([24]). Anche se più diversificata, la vegetazione della riserva naturale Ripa d’Orcia risulta simile, soprattutto per la fisionomia vegetazionale che domina l’area (boschi a sclerofille con dominanza di Quercus ilex). In base ai risultati ottenuti dal partizionamento additivo è possibile osservare che l’inserimento delle nuove riserve naturali ha contribuito ad aumentare la diversità totale della rete (sono state infatti campionate 33 specie in più rispetto alla rete preesistente, di cui 4 classificate come focali). Comunque, il loro contributo è stato soprattutto quello di accrescere la componente βrete, cioè la diversità tra riserve, che ha avuto un incremento percentuale dell’1.4% per l’insieme totale delle specie e dell’1.5% per le specie focali. L’ulteriore scomposizione della componente βrete nelle due componenti che la determinano, ovvero βarea e βreplacement, ha permesso di attribuire completamente l’aumento di diversità alla componente βarea, mentre la componente βreplacement, in proporzione, è diminuita. È necessario notare come in una rete di aree protette tra loro isolate, la diversità non dovrebbe aumentare come sola funzione dell’incremento di area dell’intera rete ma dovrebbe essere presente una certa variabilità nella diversità di ogni area protetta non associata all’aumento dell’area ([18]). Un aumento minimo o addirittura nullo della componente βreplacement, come quello osservato, è invece tipico di aree a stretto contatto o annidate, caratterizzate da una elevata omogeneità ambientale che si riflette in una bassa variabilità in composizione specifica.
Dalle analisi effettuate e dai risultati ottenuti, quindi, si osserva come l’introduzione delle due nuove riserve naturali Ripa d’Orcia e Bogatto non comporti un aumento della complementarietà tra riserve, poiché le due nuove aree presentano una componente floristica (e, quindi, una varietà di habitat) già abbondantemente rappresentata nel sistema di aree protette della provincia di Siena.
D’altra parte è necessario considerare che anche l’aggiunta di una semplice area ad un sistema di aree protette serve a garantire una certa ridondanza delle caratteristiche biologiche presenti e, quindi, la loro conservazione a lungo termine ([58]). Infatti, se da una parte il fine ultimo e principale delle attuali strategie di conservazione sta nel minimizzare il rischio di estinzione delle specie ([46], [49]), risulta chiaro che la mancanza di sufficienti informazioni in grado di tradurre il grado di rappresentatività di una specie in un dato territorio nel relativo rischio di estinzione, rende necessaria la massimizzazione della connettività, dell’estensione e della “ridondanza” delle aree protette, con l’obiettivo indiretto di garantirne la persistenza ([48]).
Conclusioni
La partizione additiva della diversità, connessa ad altre tecniche di analisi quali le curve di rarefazione, quando applicata ad un campionamento di tipo probabilistico, consente di valutare in modo preciso e ripetibile le componenti della diversità esistenti alle varie scale spaziali di riferimento. Nella rete di riserve naturali della provincia di Siena è stato possibile indicare come la grande maggioranza della diversità vegetale è attualmente allocata nella differente composizione specifica che esiste tra le singole riserve naturali (dovuta principalmente all’elevata complementarietà composizionale tra le riserve naturali). Descrivendo l’andamento nello spazio delle componenti della diversità, si è potuto valutare l’efficienza del sistema “rete di riserve naturali provinciali”, stimandola sotto diversi punti di vista, legati in particolare a: (i) la complementarietà di ambienti inclusi nella rete ([69]); (ii) la dimensione delle aree delle riserve rispondente a valori ottimali ([77]); (iii) l’incremento del numero di riserve naturali avvenuto nel tempo. In particolare, l’aggiunta delle due nuove riserve naturali, sebbene non abbia contribuito ad aumentare la differenziazione ecosistemica dell’intera rete, contribuisce alla ridondanza ambientale e specifica all’interno della rete, ritenuta una caratteristica fondamentale per la robustezza della rete di riserve naturali e per la perpetuazione delle diversità contenuta in essa ([58]).
Una valutazione a priori di nuove aree da proteggere attraverso il metodo della partizione additiva, affiancata da studi relativi alla massimizzazione di altri taxa ([10], [11]) potrebbe quindi rappresentare un efficace metodo per la selezione di nuove aree adibite alla conservazione della natura. Inoltre, è importante sottolineare la possibilità di poter quantificare tutte le componenti della diversità in maniera ripetibile, soprattutto per poter sviluppare efficienti piani di monitoraggio della diversità.
Ringraziamenti
Il progetto a cui si fa riferimento è stato realizzato grazie ad un supporto economico della Provincia di Siena e al finanziamento da parte dell’Università di Siena PAR 2005 Sviluppo di metodologie per la quantificazione e il monitoraggio della biodiversità in reti di aree protette. Si ringrazia il Corpo Forestale dello Stato per aver reso possibile l’integrazione di questo studio con le attività dell’Inventario Nazionale delle Foreste e dei serbatoi forestali di Carbonio (INFC).
Si ringraziano inoltre tutte le persone che hanno partecipato alla raccolta dei dati floristici, in particolare: Marta Chincarini, Valerio Amici, Clara Santi, Francesco Geri, Francesco Vitillo, Giulia Bennati, Sara Ghisleni, Lia Pignotti, Elisa Baragatti.
References
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar