Effects of thinning intensities in experimental plots of Black European pine in “Foreste Casentinesi, Monte Falterona and Campigna National Park” (Tosco-Romagnolo Apennine, Italy), eight years after the felling
Forest@ - Journal of Silviculture and Forest Ecology, Volume 7, Pages 73-83 (2010)
doi: https://doi.org/10.3832/efor0616-007
Published: Apr 01, 2010 - Copyright © 2010 SISEF
Research Articles
Guest Editors: 7° SISEF Congress (Pesche, IS - 2007)
« Development, adaptation, naturality and conservation »
Collection/Special Issue: Marco Marchetti, Roberto Tognetti
Abstract
Different thinning intensities were compared in a 50-years-old plantation of Pinus nigra sp. located in the “Foreste Casentinesi Monte Falterona e Campigna” National Park (northern Apennine - Italy). Six experimental square plots (900 m2) were set up for comparison of two thinning intensities: heavy thinning (F: approximately 30% of basal area removed), light thinning (d: approximately 15% of basal area removed) and control (Co). The comparison concerned: (1) the periodic increment of the trees in terms of volume (Ipv), average dbh (Ipd), height (Iph) and the variation of height/diameter ratio (S) in time; (2) forest canopy cover, indirectly expressed in term of relative irradiance (I.R.). Eight years after the felling, the results highlight that F thinning: (a) increases the homogeneity of timber, as the stand volume is concentrated in dominant trees; (b) favours tree diameter increment: Ipd values increase significantly according to the thinning intensity; (c) reduces tree height increment (lower values of Iph were recorded in the F plots); (d) promotes stand stability: lower value of S are highlighted in F plots, and no significant increase of this parameter in time has been recorded. As regards canopy cover, a significant negative correlation was recorded between stand basal area after the thinning and I.R. at ground level. After harvesting, a significant difference in terms of average I.R. between thinning intensities (F, d and Co) was also recorded: nowadays, significant differences of average I.R. are observed between Co and d, whereas F shows intermediate values: the canopy gap originated from the felling has been quickly filled. Our results highlight the growth potential of Black European pine in the studied area; however, only heavy thinning shows interesting results in terms of both tree growth and stand stability.
Keywords
Pinus nigra, Thinning, Forest stand stability, Canopy cover, Forest management
Premessa
Secondo l’Inventario Forestale della Toscana ([38]), i boschi a prevalenza di pino nero occupano 20496 ettari, di cui 7500 sono cedui coniferati e fustaie miste: tali dati sono in linea con quelli del recente Inventario Nazionale ([39]) che attribuisce ai rimboschimenti della conifera una superficie pari a 18427 ettari, di cui 10117 sono rappresentati da boschi puri.
La maggior parte di questi soprassuoli hanno ricevuto scarse e spesso occasionali cure colturali. La mancanza di diradamenti in molti casi ha portato ad uno stato di elevata fragilità che frequentemente determina crolli strutturali più o meno intensi ed estesi, rischiando di compromettere le azioni positive degli impianti in termini di difesa idrogeologica ([2], [49]).
Nella definizione classica, i diradamenti sono tagli intercalari che hanno fra le finalità principali il miglioramento della qualità della produzione legnosa, l’ottenimento di una certa quantità di prodotti intermedi, l’aumento della stabilità fisica e biologica del soprassuolo ([58]). Tali interventi, tuttavia, possono avere effetti positivi sulla rinaturalizzazione dei soprassuoli di origine artificiale ([26], [42], [10], [48], [13], [18]). La letteratura italiana ([30], [51]) e quella estera ([31], [23] e [24], [34], [28]) mettono in evidenza come modificazioni del grado di copertura del soprassuolo principale si manifestino con variazioni della radiazione luminosa al suolo, e come tutto ciò sia strettamente collegato con la possibilità di affermazione di piante nel piano di rinnovazione.
Il presente lavoro, iniziato nel 2000 da una collaborazione fra DISTAF dell’Università di Firenze e la Comunità Montana del Casentino (AR), ha come obbiettivo la valutazione degli effetti indotti dalla riduzione della copertura su: (1) incremento legnoso delle piante; (2) dinamiche successionali. Dal momento che la quantità di energia radiante che arriva sotto copertura è uno dei fattori che maggiormente influenzano le dinamiche evolutive ([45], [47], [67], [6], [7], [53]), le variazioni del grado di copertura indotte dal taglio intercalare sono state valutate in termini di modificazioni di disponibilità luminosa al livello del suolo.
L’ambiente
L’area oggetto di studio è localizzata nel comune di Stia (AR), nella proprietà agricolo-forestale della Regione Toscana gestita dalla Comunità Montana del Casentino (complesso “Foreste Casentinesi”), all’interno del Parco Nazionale delle Foreste Casentinesi, Monte Falterona e Campigna.
Le temperature medie annue oscillano tra 9.2 °C della Verna (950 m s.l.m.), con 18 °C di escursione media annua, e 14 °C di Subbiano (300 m s.l.m.), con 16 °C di escursione media annua, mentre la piovosità va da 1700 mm annui a Camaldoli (1100 m s.l.m.), con massimo autunno-invernale e minimo estivo senza piovosità, a 1016 mm a Subbiano (con regime analogo a quello di Camaldoli, ma con due mesi aridi estivi).
Facendo riferimento alla classificazione del Pavari, l’area di studio si colloca in una fascia fra il Castanetum, sottozona fredda, e il Fagetum, sottozona calda.
Seguendo, invece, la classificazione climatica di Rivas-Martínez et al. ([61]) Stia può essere inquadrata nel macrobioclima temperato, bioclima oceanico temperato (Teoc), termotipo mediterraneo (Mte), ombrotipo umido (Hum).
Secondo la classificazione di Ozenda ([55]), la vegetazione del territorio preso in esame è inquadrabile nella fascia collinare (serie della roverella, del cerro, dei carpini e della farnia) e in quella montana (serie della faggeta e della faggeta-alpina).
Facendo riferimento alla carta della vegetazione potenziale della Regione Toscana di Mondino ([50]), nel territorio studiato si distinguono le zone delle cerrete (fino a 1000 m s.l.m.), dei castagneti (solo su suoli arenacei, fino a 900 m s.l.m.), delle faggete e delle abetine (oltre 900-1000 m s.l.m.).
La geologia del territorio in cui sono comprese le “Foreste Casentinesi” è quasi interamente rappresentata da arenaria Macigno dell’Olocene (Macigno del Chianti e Macigno del Mugello), con affioramenti di Marne di Vicchio nella zona di Moggiona.
L’impianto in cui è stata condotta l’indagine è una pineta di pino nero ubicata a 850 m s.l.m., con esposizione Est - Sud Est, che presenta una pendenza media del 20% ed è costeggiata da una strada forestale camionabile. Attualmente il soprassuolo ha un’età di 50 anni, anche se questa deve essere considerata come l’età prevalente del popolamento; infatti dopo l’impianto i risarcimenti sono proseguiti per 4-5 anni utilizzando anche materiale di diversa età (secondo le disponibilità del vivaio): il risultato è che in una stessa parcella l’età delle piante può variare in un range di 7-8 anni.
Materiali e metodi
Nel 2001 sono state delimitate in maniera permanente sei parcelle, di forma quadrata con 30 m di lato, adiacenti ma separate le une dalle altre da “corridoi” di 10 m (corrispondenti a circa 4 file di piante). Al centro di ogni parcella è stato realizzato un transect lineare di 30 x 5 m; nel complesso l’intera area sperimentale occupa una superficie di 9600 m2.
In ciascuna parcella, per ogni pianta sono state eseguite le seguenti operazioni: 1) numerazione, 2) misura del diametro a 1.30 m (sono state prese due misure ortogonali fra loro per ogni individuo), 3) attribuzione della posizione sociale secondo tre categorie, prendendo spunto dalla classificazione di Kraft (in [58]): dominanti (D, con chioma inserita nella parte superiore dello strato, vigorosa ed omogeneamente espansa in ogni direzione), codominanti (Cd, con chioma inserita nella parte superiore dello strato ma almeno su un lato marcatamente compressa a seguito della concorrenza con gli individui adiacenti), dominate (dm, con chioma ridotta, del tutto o in parte compressa e spesso sovrastata da quella di individui limitrofi).
In ciascun transect, per ogni pianta è stato rilevato: (1) numero; (2) altezza totale (usando l’ipsometro Vertex II con transponder a 1.30 m).
Prima dell’attribuzione della tesi sperimentale sono state verificate le condizioni di omogeneità strutturali fra le parcelle ([12]). Successivamente a ciascuna coppia di parcelle è stata attribuita casualmente una tesi che prevedeva una diversa intensità di intervento: (1) asportazione del 15% dell’area basimetrica (d, parcelle I e V); (2) asportazione del 30% dell’area basimetrica (F, parcelle II e IV); (3) nessun intervento (Co, parcelle III e VI).
Il tipo di diradamento è quello che comunemente viene applicato dai tecnici della Comunità Montana del Casentino, ovvero un diradamento misto che abbina un diradamento dal basso (anche se occasionalmente qualche pianta dominata viene rilasciata) a interventi di selezione positiva volti a favorire individui particolarmente promettenti.
L’utilizzazione delle piante è avvenuta nel febbraio del 2002: delle piante abbattute è stato determinato il volume cormometrico (comprensivo di corteccia) con il metodo per sezioni di Heyer ([44]). Nel settembre del 2009 è stato rilevato il diametro a 1.30 m di ogni pianta di ciascuna parcella, per quelle ricadenti all’interno del transect è stata rilevata anche l’altezza.
Nel luglio 2002, 2006 e 2009 sono state eseguite le misure della radiazione solare. In particolare è stata misurata l’irradianza nel campo della PAR, utilizzando una coppia di ceptometri (Sun Scan Canopy Analysis System, AT Delta-T Devices Ltd). Le misure sotto copertura sono state eseguite lungo l’allineamento centrale di ciascun transect; simultaneamente sono stati rilevati i valori di irradianza in piena luce in un’ampia radura adiacente all’impianto. Le misure sono state eseguite durante le ore centrali del giorno, in un arco di tempo compreso fra le 11.30 e le 12.30 (ora solare), in giornate serene (situazione corrispondente ad almeno 1850 μmol m-2sec-1 in piena luce).
Elaborazione dei dati
Per valutare l’incremento medio periodico di volume (Ipv, m3 ha-1anno-1, riferito alle stagioni vegetative dal 2002 al 2008) è stata adottata una tavola di cubatura empirica messa a punto dai tecnici della Comunità Montana del Casentino per le pinete di pino nero del territorio. La tavola restituisce il volume cormometrico (V, espresso in m3, comprensivo di cimale e corteccia) in funzione del diametro a 1.30 m (dbh, espresso in cm) secondo il seguente algoritmo (eqn. 1):
Ipv è stato calcolato per differenza (eqn. 2) fra il volume del soprassuolo subito dopo l’intervento (V1) e quello derivante dal rilievo delle piante nel 2009 (V2 - eqn. 2):
Con i test dell’ANOVA e di Duncan (in caso di valori di F significativi) sono stati analizzati i valori medi di:
- incremento periodico di diametro (Ipd, in cm), ponendo come fonti di variazione: (i) tesi; (ii) posizione sociale (escludendo le dm); e (iii) interazione (i x ii);
- incremento periodico di altezza (Iph, in m), ponendo come fonti di variazione: (i) tesi; (ii) posizione sociale (escludendo le dm); e (iii) interazione (i x ii).
- rapporto di ipsodiametrico (S = h/dbh), ponendo come fonti di variazione: i) tesi; (ii) posizione sociale (escludendo le dm); e (iii) interazione (i x ii).
- Irradianza relativa (IR), ovvero il rapporto fra le misure di irradianza prese sotto copertura e quelle all’aperto, analizzata per ogni anno rilevato e ponendo come fonti di variazione le tesi.
Risultati
Facendo riferimento alla situazione prima dell’intervento e riferendo i dati ad ha (Tab. 1), nelle parcelle sottoposte alla tesi F erano presenti 1200 individui, per un area basimetrica di 58.9 m2, in quelle della tesi d erano presenti 1360 piante, per un area basimetrica di 61.6, mentre nelle parcelle della tesi Co erano presenti 1200 piante, per un’area basimetrica di 55.8 m2.
Tab. 1 - Principali dati dendrometrici delle parcelle prima dell’intervento. n°: numero di piante; G: area basimetrica; dg: diametro della pianta con area basimetrica media; hdg: altezza della pianta con diametro pari a dg).
| Parcella | n°/ha | G (m2 ha-1) |
dg (cm) |
hdg (m) |
|---|---|---|---|---|
| I | 1256 | 60.8 | 24.8 | 17.5 |
| II | 1222 | 59.8 | 25.0 | 17.6 |
| III | 1133 | 59.3 | 25.8 | 17.7 |
| IV | 1178 | 58.1 | 25.1 | 17.6 |
| V | 1467 | 62.3 | 23.3 | 17.2 |
| VI | 1267 | 52.2 | 22.9 | 17.1 |
Il taglio intercalare ha asportato, nella tesi F, il 26.4% dell’area basimetrica (in questo caso la massa asportata è stata inferiore a quella preventivata per non oltrepassare i limiti imposti dal Regolamento Forestale della Toscana - n. 48/R, DPGR 8 agosto 2003, art. 30) pari a 279.0 m3 ha-1 e al 40.3 % degli individui e nella tesi d il 14.5% dell’area basimetrica - pari a 143.1 m3 ha-1 e al 22% degli individui.
Incrementi periodici
Volume (Ipv)
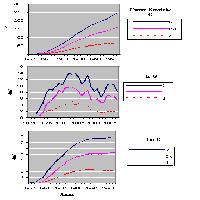
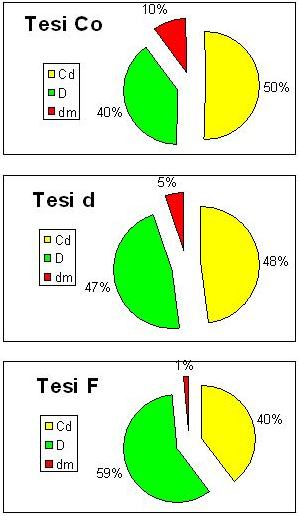
Nella tesi F l’Ipv risulta pari a 11.6 m3 ha-1 anno-1, la massa in piedi è 460 m3 ha-1. Nella tesi d l’Ipv è pari a 12.2 m3 ha-1anno-1, la massa in piedi a 536 m3 ha-1; nella tesi Co l’Ipv è 9.3 m3 ha-1 anno-1, la massa in piedi 514 m3 ha-1. In Fig. 1 è riportata la ripartizione della massa in piedi delle piante distinte in base alla posizione sociale. Nella tesi F la maggior parte della massa è concentrata nelle piante D (59%) mentre in Co nelle Cd (50%). Le piante dm costituiscono il 10 % del volume in Co e solo l’1% in F, mentre nella tesi d si osservano valori intermedi.
Fig. 1 - Ripartizione della massa in piedi (volume cormometrico) fra le piante, distinte in base alla posizione sociale nelle tesi messe a confronto (per la simbologia si rimanda a materiali e metodi).
Diametro (Ipd)
L’ANOVA ha evidenziato differenze molto significative (p<0.01) dei valori medi di Ipd, confrontando sia le tesi (F[2; 449] = 13.21) sia le posizioni sociali (F[1; 449] = 89.43), mentre l’interazione fra le due suddette fonti di variazione non risulta statisticamente significativa. Il test di Duncan separa significativamente il valore medio rilevato in F (2.8 cm) da quello di d (2.4 cm), e quest’ultimo da quello di Co (2 cm). Lo stesso test mette in luce che Ipd medio per le piante D (3 cm) è significativamente più alto delle piante Cd (1.9 cm).
Altezza (Iph)
I valori medi di Iph, in base al test ANOVA, danno luogo a differenze molto significative (p<0.01) fra le tesi confrontate (F[2;118] = 6.95), ma non fra le posizioni sociali e l’interazione delle due fonti di variazione. In base al test di Duncan, Iph medio in Co (3.6 m) è significativamente più alto che in F (2.6 m), con valori intermedi in d (3.2 m), non significativamente diversi da quelli delle altre tesi.
Rapporto ipsodiametrico (S)
Subito dopo il taglio intercalare il valore medio di S è pari a 69 nelle tesi d e Co e a 70 in F: le differenze fra le tesi non sono statisticamente significative (F[2;118] = 0.33). Differenze molto significative (F[2;118] = 71.32, P<0.01) emergono invece fra le posizioni sociali (Cd = 76, D = 62).
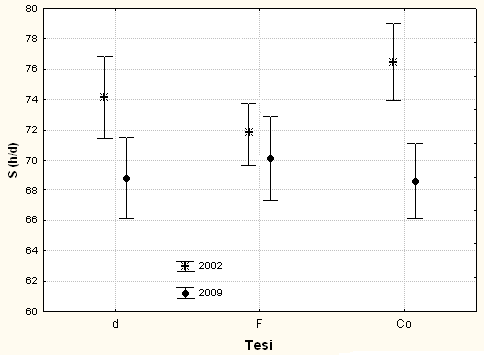
L’ANOVA, per i valori medi di S nel 2009, evidenzia differenze significative (P<0.05) fra le tesi (F[2;118] = 3.18) e molto significative (P<0.01) fra le posizioni sociali (F[1;118] 90.24), mentre l’interazione fra le due fonti di variazione non risulta statisticamente significativa. Il test di Duncan mette in luce differenze significative tra le tesi Co (77) e F (70), mentre valori intermedi e non significativamente differenziati rispetto agli altri spettano a d (74). Lo stesso test evidenzia che il valore medio di S nella tesi Cd (82) è significativamente più alto di quello della tesi D (67). Il valore di S, dal 2002 e al 2009, è aumentato significativamente (F[2;236] = 3.039; p<0.05) solo nel caso delle tesi Co e d (Fig. 2).
Fig. 2 - Valori medi di S (con limiti fiduciari per p<0.05), distinti per tesi, subito dopo l’utilizzazione (2002) e nel 2009.
Variazione dell’irradianza relativa (I.R.)
I valori medi di irradianza sotto copertura sono cambiati sensibilmente nel corso della sperimentazione (Fig. 3).
Fig. 3 - Valori medi di I.R. (con limiti fiduciari per p<0.05): confronto fra tesi nelle varie annate di rilievo.
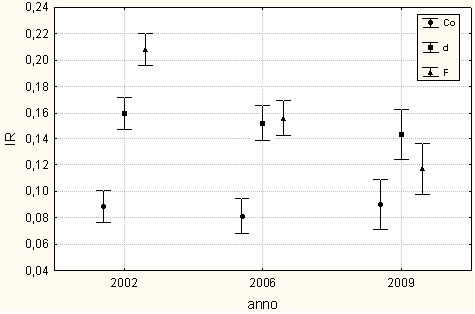
Nell’estate seguente all’intervento (2002), i valori medi di I.R. si differenziavano statisticamente (P<0.01) fra le tesi (F[2;1918] = 81.57): in particolare il test di Duncan evidenzia differenze significative fra tutte e tre le tesi messe a confronto (Co 9%, d 16%, F 21%).
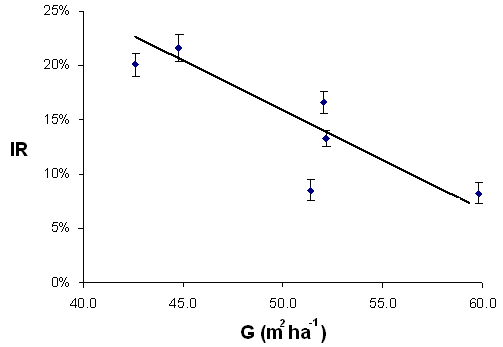
I valori medi di I.R. rilevati sotto copertura in ciascuna parcella sono correlati negativamente (r=-0.92) e molto significativamente (P<0.01) con l’area basimetrica (G) ad ettaro (Fig. 4).
Fig. 4 - Correlazione fra area basimetrica (G) dopo il taglio intercalare e irradianza relativa media (IR ± e.s.).
In base all’ANOVA i valori medi di I.R. rilevati nel 2006 risultano significativamente (P<0.01) diversi fra le tesi (F[2;1553] = 53.1): il test di Duncan mette in evidenza differenze significative fra Co (8.6 %), da una parte, e d (15.5) ed F (15.8), omogenei fra loro, dall’altra.
Anche nel 2009 i valori medi di I.R. risultano significativamente (P<0.01) diversi fra le tesi (F[2;765] = 10.11), ma in questo caso la significatività emerge fra Co (9 %) e d (14.8 %), mentre F (12.5 %) non si differenzia in maniera significativa dalle altre due tesi.
Discussione
Dopo 8 stagioni vegetative dall’intervento, i risultati della ricerca sono in linea sia con quelli ottenuti nelle pinete di pino laricio della Calabria ([21], [9], [4]), nei rimboschimenti di pino nero umbri ([20]) e in quelli toscani ([3], [18]): nelle pinete di buona fertilità, il pino manifesta una grande reattività al taglio intercalare, con accrescimenti proporzionati all’intensità dell’intervento. L’incremento periodico di volume stimato in questo studio per le parcelle diradate è riferibile a quello delle specie a rapido accrescimento (>10 m3 ha-1 anno-1 - [22]) ed è paragonabile con i risultati di indagini dendro-auxometriche condotte nelle pinete della zona ([52], [66]).
La posizione sociale degli individui prima del taglio intercalare esercita grande influenza sui successivi accrescimenti: i diradamenti non esaltano l’incremento legnoso delle piante di medie dimensioni (codominanti), bensì quello delle piante di maggiori dimensioni (dominanti). Riteniamo che il fenomeno sia dovuto al fatto che, all’aumentare dell’intensità del prelievo, aumenta la quantità di luce che arriva alla parte bassa della chioma: ne consegue che le piante che traggono maggior beneficio sono quelle la cui chioma è già ben sviluppata prima del taglio intercalare (piante dominanti).
A seguito dell’intervento più intenso la maggior parte della massa si concentra nelle piante dominanti, con ricadute positive sulla qualità degli assortimenti ottenibili a fine turno, dovuta alla maggiore uniformità del materiale legnoso ([9], [16]).
Come ampiamente dimostrato per l’abete rosso, l’abete bianco e la douglasia ([1], [43], [5], [63]) il rapporto h/dbh è inversamente correlato alla stabilità dei soprassuoli: tale parametro tende inoltre ad aumentare con l’età del popolamento. Nel nostro caso, l’influenza positiva del diradamento sulla stabilità delle piante del soprassuolo è evidente solo nella tesi F. Infatti, solo nella tesi in cui il diradamento è stato più intenso il valore medio di S è risultato significativamente inferiore rispetto a quello delle parcelle non diradate: tale valore, inoltre, non è variato significativamente negli ultimi 8 anni. Quest’aspetto evidenzia come solo gli interventi di una certa intensità siano efficaci nel promuovere la stabilità dei popolamenti, altrimenti destinata a diminuire nel tempo.
La progressiva riduzione dei valori di I.R. osservati nella tesi F indica il graduale recupero della copertura a seguito del taglio intercalare. Del resto, anche se non sono state fatte misurazioni specifiche per valutare l’espansione della chioma, è nota la relazione diretta fra sezione trasversale dell’alburno e LAI ([29], [15]). Nella tesi d, la presenza di un più elevato numero di piante Cd (con incrementi diametrici inferiori e quindi minor velocità di espansione della chioma) ostacola l’espansione della chioma delle piante D, determinando così la più lenta chiusura della copertura. Anche Sullivan et al. ([65]), in uno studio effettuato in una pineta di Pinus contorta del Nord America, osservano un maggior incremento medio dello sviluppo delle chiome nelle parcelle diradate più intensamente. Nei boschi di latifoglie in Québec (Canada), Beaudet & Messier ([7]) concludono che la differenza di radiazione luminosa sotto copertura tra le parcelle diradate e quelle non diradate decresce in funzione del tempo e che la velocità del cambiamento è molto maggiore nei primi anni dopo il taglio; ciò conferma i risultati di questo studio, secondo il quale dopo 8 anni le differenze fra F e Co sono ormai quasi azzerate. Il diradamento più intenso è evidentemente molto efficace ad esaltare le potenzialità incrementali della specie, senza pregiudicare, anche nel breve periodo, la continuità copertura.
Hale ([34]), per Picea sitkensis in Gran Bretagna, evidenzia come la correlazione tra irradianza relativa e intensità del diradamento possa essere usata come guida per i gestori che desiderano “modificare” l’ambiente luminoso al fine di incoraggiare la rinnovazione naturale. Per assicurare lo sviluppo del piano di rinnovazione in tali popolamenti è suggerito un livello di irradianza relativa minima del 20%, corrispondente a diradamenti di intensità pari al 45% dell’area basimetrica. La relazione che lega l’irradianza all’area basimetrica è sottolineata anche in altri studi ([40], [57],[25], [59], [35]), e nel nostro caso può essere quantificata in un aumento di circa il 10% della irradianza relativa in corrispondenza di una riduzione di 10 m2 di area basimetrica. Tutto ciò potrebbe assumere un significato negli interventi di rinaturalizzazione tramite sottopiantagione, come proposto da Grifoni ([32]) e Cantiani et al. ([19]), nel momento in cui si vogliano creare le condizioni idonee per lo sviluppo delle piante introdotte. A tal fine, il taglio intercalare dovrà precedere l’intervento di sottopiantagione e l’intensità sarà in funzione del livello di irradianza che si vuol ottenere all’interno dell’impianto, a sua volta legata alle esigenze della specie che si vuol introdurre ([13]). Un lavoro inerente a sottopiantagioni di Picea glauca e Pinus banksiana ([46]) in boschi misti a dominanza di pioppo (Populus tremuloides) e betulla (Betula papyrifera) evidenzia che l’accrescimento dei semenzali aumenta con l’intensità del prelievo prima della sottopiantagione, e mette in rilievo alti rischi di insuccesso qualora si effettuino sottopiantagioni in soprassuoli non diradati. Krasowski & Wang ([41]), confrontando l’effetto sulla rinnovazione di Abies lasiocarpa di diverse intensità di diradamento in popolamenti di Betula papirifera, pongono l’accento su come la copertura residua influenzi positivamente lo sviluppo dei semenzali, e concludono che i diradamenti sono il modo migliore per gestire soprassuoli misti di conifere e latifoglie.
Studi condotti in Giappone ([68]) evidenziano che i diradamenti sono efficaci per promuovere l’insediamento dei semenzali nel piano di rinnovazione nelle pinete litoranee di Pinus thunbergii, con densità crescenti con l’intensità dell’intervento. Sullivan et al. ([65]) concludono che, in giovani soprassuoli di Pinus contorta del Nord America sottoposti a diverse intensità di diradamento, dopo dieci anni dall’intervento sia l’incremento medio delle piante sia la diversità specifica e strutturale del soprassuolo sono significativamente più alti nei popolamenti più radi (dove cioè il grado di diradamento è stato forte o medio), rispetto a quelli più densi (soprassuoli diradati debolmente o non diradati). Cameron ([16]) e Nyland ([53]) evidenziano come i diradamenti siano essenziali per passare da strutture regolari a strutture irregolari, le più idonee nel favorire una maggiore diversità di nicchia nei popolamenti forestali ([27], [54], [56]).
In uno studio condotto sulle dinamiche strutturali delle pinete di pino nero del Casentino ([10]) è stato osservato che, in popolamenti analoghi (per caratteristiche stazionali ed età del popolamento) a quello oggetto della nostra indagine, è presente (o comunque si inizia a delineare) un piano inferiore di latifoglie (acero montano, frassini, cerro, castagno, ecc.) con o senza partecipazione di abete bianco. Nella pineta oggetto di questo studio l’assenza di un piano di successione non è attribuibile né alla disponibilità di seme (nelle immediate vicinanze sono presenti abetine di abete bianco, cerrete, faggete, oltre a gruppi di piante di frassino maggiore, castagno, acero di monte, ecc.) né a livelli radiativi insufficienti (quelli rilevati sarebbero comunque compatibili con le esigenze di molte specie tolleranti l’ombra). Riteniamo piuttosto che il vero fattore limitante per la rinnovazione forestale sia costituito dal carico di ungulati presente nelle Foreste Casentinesi (Fig. 5). Nel territorio in cui si è svolta l’indagine sono presenti popolazioni di cervo, capriolo, daino, cinghiale: la sola popolazione di cervo presenta una densità di 15.5 capi per 100 ha ([33]). Vale la pena di segnalare che l’impatto negativo dell’eccessivo carico di fauna ungulata sulle dinamiche successionali è ampiamente documentato sia nel territorio del Parco ([14]), sia in altre zone italiane ed estere ([62], [17], [8], [36] [37], [60]).
Fig. 5 - Semenzali di abete bianco di 1 anno (sulla sinistra) e di 4 anni (quest’ultimo ripetutamente brucato).

Conclusioni
Il taglio intercalare promuove l’incremento legnoso delle piante, fenomeno evidente soprattutto con l’intervento più intenso e per le piante con chioma ben conformata (dominanti). Il diradamento meno intenso influisce poco sulla qualità e sull’omogeneità degli assortimenti ritraibili a fine turno, inoltre è meno efficace nel promuovere l’incremento diametrico e la stabilità delle piante del soprassuolo.
Gli impianti adulti di pino nero, nelle situazioni di miglior fertilità, evolvono piuttosto velocemente verso il bosco misto con latifoglie ([49], [10]). Col presente lavoro si è cercato di mettere in evidenza che, con le comuni pratiche colturali, si creano anche le condizioni favorevoli per “l’avvio” di dinamiche successionali. Intervenendo con un’appropriata intensità di prelievo, il gestore potrà indirizzare la dinamica dei popolamenti mediante sottopiantagioni di specie di pregio se queste non sono presenti, o selezionando quelle in ingresso. Il soprassuolo di pino rimasto in piedi, oltre a garantire ottimi accrescimenti, svolgerà una funzione protettiva nei confronti dei piani inferiori. La frequenza e l’intensità degli interventi sarà dettata dalle esigenze luminose delle specie in ingresso e dalla necessità di ridurre i danni al novellame ([64]) al fine di garantirne uno sviluppo ottimale. Quando uno o più piani inferiori sono definiti, gli interventi colturali assumono una funzione diversa da quella dei tradizionali diradamenti, divenendo “tagli di liberazione” mirati alla progressiva affermazione dei piani inferiori ([11]).
La gestione dei soprassuoli forestali mirata ad assecondarne le dinamiche evolutive trova nell’eccessiva pressione della fauna uno dei fattori maggiormente limitanti, al punto di sconsigliarne l’applicazione fino a quando l’equilibrio fra carico di animali selvatici e offerta alimentare del territorio non sia ristabilito.
Ringraziamenti
Hanno collaborato ai rilievi in campo: Alessandro Varallo, Francesco Calzolari, Filippo Cartei, Enrico Stefanini, Francesca Giannetti. Gli autori sono grati alle guardie e agli operai della Comunità Montana del Casentino per il costante impegno con il quale hanno seguito negli anni la sperimentazione.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar