Basic knowledge for forest restoration: the Pomieri forest case study (Madonie regional park, Sicily)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 9, Pages 8-19 (2012)
doi: https://doi.org/10.3832/efor0679-008
Published: Feb 28, 2012 - Copyright © 2012 SISEF
Research Articles
Abstract
The purpose of this paper is to highlight a case study of forest restoration in the Pomieri Wood area (Madonie regional park, Sicily), consisting of a relict population of sessile oak (Quercus petraea subsp. austrothyrrenica Brullo, Guarino & Siracusa). To assess the functionality of the stand some biological indicators were taken into consideration: dendrometric-structural, floristic-vegetation, ecological, fructification, natural regeneration, disturbances. They reveal a state of degradation after a long period of overexploitation of resources. Forest restoration would lead to the regaining of the cultural value of the Pomieri wood by management authorities and local populations. Then a series of measures are to be carried out providing for the removal of degradation factors (grazing, illegal cuttings) and restoring stand density, biodiversity and ecological functionality.
Keywords
Introduzione
Il restauro forestale assume i connotati di una nuova disciplina, rivolta ad attuare misure per il recupero funzionale di ecosistemi forestali degradati anche in ambiente mediterraneo ([40]). Infatti, il restauro degli ecosistemi forestali degradati oltre che un preciso obiettivo di gestione nelle aree protette ([8]) è uno dei maggiori obiettivi della ricerca forestale applicata e delle strategie di politica forestale in vari Paesi ([54], [29], [48], [55], [56]).
In questo contributo si è preso in esame il Bosco Pomieri, localizzato nel Parco Regionale delle Madonie (Sicilia settentrionale), dominato dalla rovere che Brullo et al. ([12]) riferiscono a Quercus petraea (Matt.) Liebl. subsp. austrotyrrhenica Brullo, Guarino & Siracusa, allopatica rispetto alla sottospecie nominale, occupando le stazioni più meridionali dell’areale della specie in Italia e in Sicilia. Il bosco, pertanto riveste un particolare interesse geobotanico e naturalistico ([10], [45], [47], [13], [50]) e rappresenta presumibilmente una delle aree rifugio e di origine della diversità genetica prima della migrazione post-glaciale ([20]), ma ad alto rischio di estinzione per la bassa differenziazione genetica della popolazione ([14], [3]).
Il bosco è stato oggetto di un pesante sfruttamento fra la prima e la seconda guerra mondiale e negli anni 1950, 1952, 1953. Oggi nel quadro di mutate condizioni economico-sociali e per l’affermarsi di un diverso interesse per il bosco si pone il problema della sua salvaguardia. Tuttavia quest’ultimo termine appare generico e potrebbe significare solo misure di esclusione di interventi, anziché misure proattive di riavvio dei processi funzionali. Da qui il restauro forestale, inteso innanzitutto come la riacquisizione culturale del valore del bosco da parte degli enti territoriali di gestione e delle popolazioni locali, e poi come l’applicazione di una serie di misure che prevedano sia la rimozione dei fattori di degrado che interventi colturali di miglioramento riguardo alla densità, biodiversità, struttura, funzionalità ecologica.
Materiali e metodi
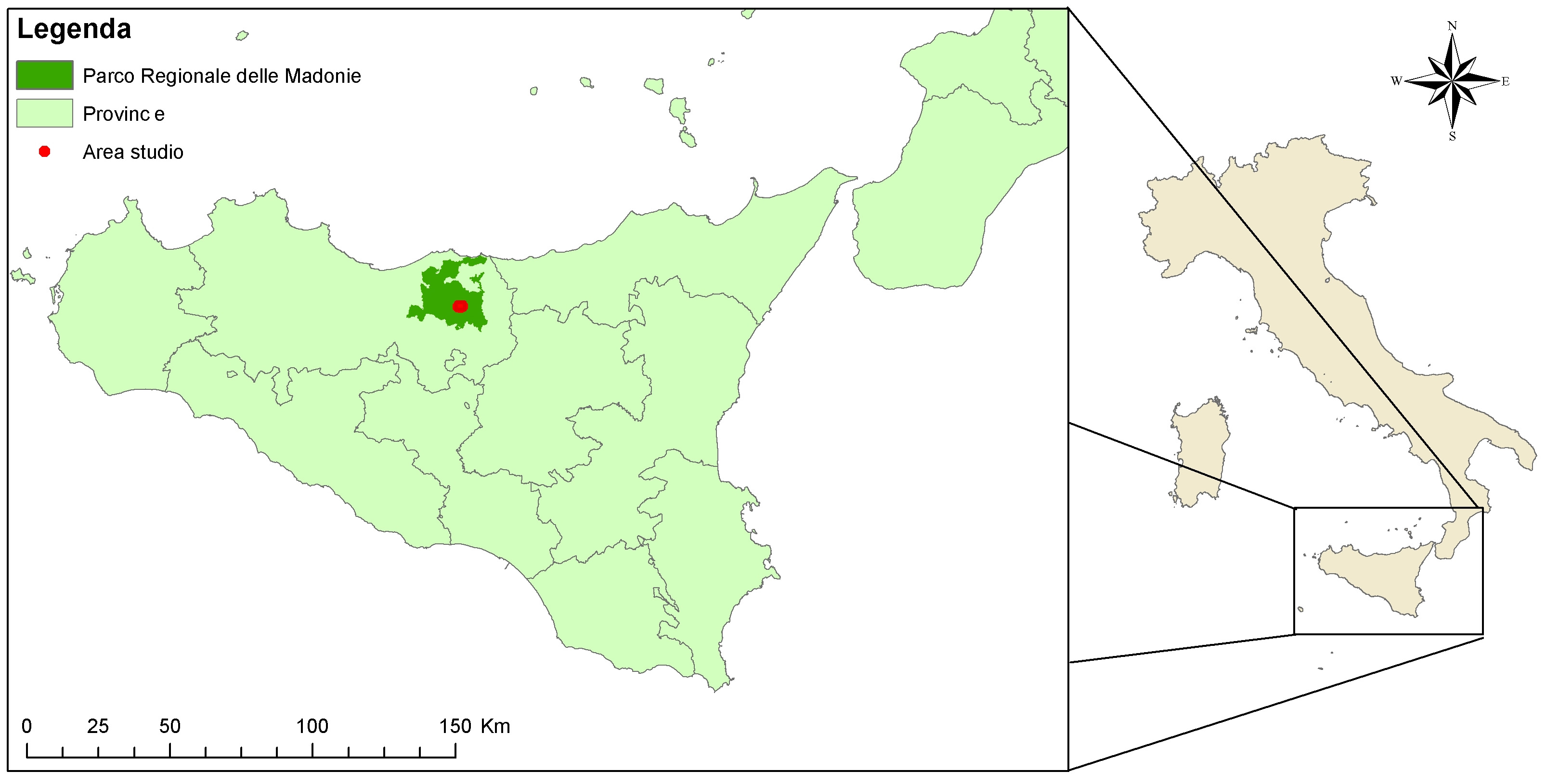
Il Bosco Pomieri, inserito in zona A del Parco Regionale delle Madonie (Comune di Petralia Sottana, Palermo), si estende per circa 1000 ha, tra 1200-1300 e 1500 m s.l.m. (Fig. 1).
Fig. 1 - Localizzazione dell’area di studio.
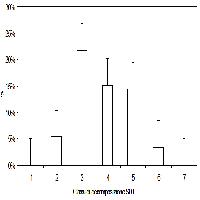
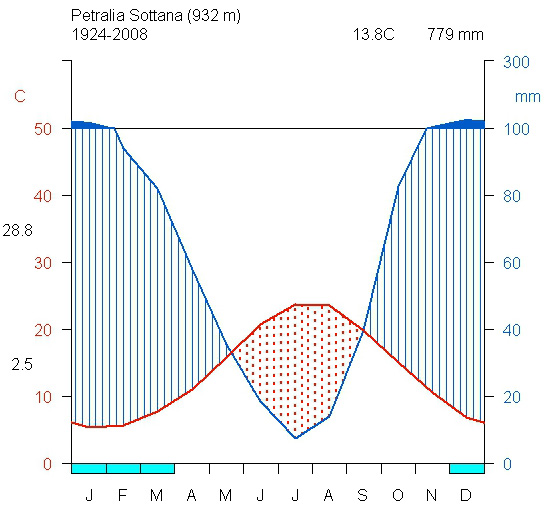
Per la caratterizzazione climatica mancano stazioni di riferimento collocate alla stessa quota. L’unica stazione vicina è quella di Petralia Sottana (Fig. 2) localizzata a 930 m s.l.m. su un diverso versante del sistema montuoso delle Madonie. Qui la temperatura media annua è di 13.8 °C, mentre la precipitazione media annua è di 779 mm. Le precipitazioni nevose interessano il periodo dicembre-febbraio. Per la definizione del bioclima si è quindi fatto riferimento all’Atlante climatologico della Sicilia ([1]). In accordo con Rivas-Martinez ([49]) e Brullo et al. ([11]) l’area oggetto di studio è caratterizzata da un bioclima mediterraneo pluvio-stagionale oceanico con termotipo suprameditterraneo e ombrotipo umido inferiore.
Fig. 2 - Climogramma di Walter & Lieth ([53]), generato con il modulo CLIMATOL in ambiente R, della stazione di Petralia Sottana (PA).
Il substrato pedogenetico è costituito prevalentemente da quarzareniti del Flysch numidico ([33]). I suoli profondi sono riferibili agli Inceptisuoli (Suoli Bruni) e agli Alfisuoli ([22], [23]).
Il Bosco Pomieri è caratterizzato dalla presenza della rovere meridionale (Quercus petraea subsp. austrothyrrenica Brullo, Guarino & Siracusa), spesso presente in esemplari secolari e con un denso strato arbustivo dominato dall’agrifoglio (Ilex aquifolium L.), ad essi si associano nelle aree più fresche ed alle quote superiori, Fagus sylvatica L., Acer pseudoplatanus L., Acer obtusatum W. & K., Ulmus glabra Huds., Sorbus torminalis L., Malus sylvestris (L.) Mill., sporadicamente Quercus cerris L. e altre querce del ciclo di Quercus pubescens, e varie specie arbustive quali Daphne laureola L., Crataegus laciniata Ucria, Ruscus aculeatus L., Rhamnus catharticus L. ([10], [44], [45], [46]).
Il bosco di rovere meridionale sotto l’aspetto fitosociologico rientra nell’associazione Ilici-Quercetum austrotyrrhenicae (Ilici-Quercetumpetraeae Brullo & Marcenò in [10]), descritto da Brullo ([10]) proprio per questo territorio. L’associazione rientra nel Doronico-Fagion ([26]) [52], alleanza dell’ordine Fagetalia sylvaticae Pawlowski in Pawlowski et al. (1928) e della classe Querco-Fagetea Br.-Bl. Vlieger in ([13]).
Le caratteristiche della vegetazione denotano condizioni climatiche con accentuati caratteri di oceanicità, in relazione alle correnti umide provenienti dal mar Tirreno che accrescono l’apporto idrico sotto forma di precipitazioni occulte ([10], [50]) che non è messo in evidenza dai dati della stazione termo-pluviometrica.
Secondo i tipi forestali della Sicilia ([28]) il Bosco Pomieri è inquadrato come segue:
- categoria: Querceti di rovere e roverella; tipo: Querceto di rovere (QU10X);
- categoria: Faggeta mesofila dei substrati silicatici; variante con faggio (FA10B).
Sono state materializzate al suolo sei aree di saggio (AdS) di 50 x 50 m (2500 m2); il disegno di campionamento è di tipo soggettivo ed il numero delle AdS varia da cinque per in popolamenti QU10X (AdS 1, 2, 3, 4, 6) a una AdS in un popolamento FA10B a seconda dell’estensione e della rappresentatività nell’area studio.
Rilievi dendronetrici e strutturali
In ogni AdS, nella stagione 2008-2009, sono stati effettuati i seguenti rilievi: (1) cavallettamento totale delle piante vive con diametro a 1.30 m ≥ 2.5 cm, altezze (sul 20% delle piante rappresentative di tutte le classi diametriche e delle 25 piante di diametro maggiore); (2) età, mediante succhiellamento a 1.30 m degli alberi dominanti; (3) tipo di struttura: mono-, bi-, multi-plana; per la caratterizzazione della struttura verticale è stato applicato l’indice di Latham et al. ([32]); (4) grado di copertura in %; (5) stato vegetativo (senescenza, stress ambientali, patologie, attacchi di insetti); (6) azioni di disturbo.
In ogni AdS si è proceduto alla realizzazione di un transect di struttura, diagonale all’area di saggio, delle dimensioni di 40 x 6 m in cui sono stati raccolti i seguenti dati: (1) classificazione dei soggetti arborei distinguendo la specie, l’origine (gamica o agamica) ed il diametro a 1.30 m; (2) altezza totale e di inserzione del primo ramo verde in m; (3) posizione sociale (dominante, intermedia, dominata); (4) posizione topografica; (5) raggio della chioma secondo le 4 direzioni geografiche in m.
Procedendo alla restituzione grafica con l’utilizzo del software SVS 3.31 (SUAOE VJCVAMJAUJPO SZCUFN, Pacific Northwest Research Station, USDA Forest Service, USA) è stato possibile: (1) evidenziare sia la distribuzione nello spazio orizzontale e verticale; (2) valutare il grado di copertura (valore percentuale della superficie coperta dalle chiome riferita alla superficie del transect).
Sono state rilevate le ceppaie presenti distinguendo se di origine naturale o artificiale (taglio). Ceppaie e soggetti morti in piedi sono stati posizionati acquisendo le coordinate con un GPS.
Rilievi della necromassa
Per ogni AdS è stata stimata la necromassa, indicatore Pan-Europeo per la gestione forestale sostenibile (Criterio 4: Conservazione e appropriato miglioramento della biodiversità negli ecosistemi forestali - [39]) e, per questo, considerata tra gli indicatori di sostenibilità ambientale. La distribuzione spaziale, il volume, la classe e il grado di decomposizione della necromassa possono essere usati come indicatori della biodiversità e della funzionalità degli ecosistemi forestali ([24], [38], [25]). In questo studio la necromassa è distinta in: (1) necromassa in piedi (SDT, Standing Dead Trees), che comprende le piante morte in piedi, intere o troncate; (2) necromassa a terra, (CWD, Coarse Woody Debris) costituita da rami, ceppaie, fusti di alberi e arbusti morti che sono caduti e che si trovano sul terreno ([21], [34]).
La SDT è stata stimata misurando in tutti i soggetti presenti nell’area con diametro ≥ 2.5 cm a 1.30 m, il diametro a metà lunghezza e la lunghezza totale. Inoltre, ogni campione di SDT è stato riferito ad una delle classi di decomposizione di Hunter (Maser et al. 1979 in [38]). Le elaborazioni per la stima della SDT hanno riguardato: (1) il numero di fusti morti in piedi; (2) la percentuale di piante morte in piedi sul totale; (3) la ripartizione in classi di diametro; (4) la ripartizione percentuale della SDT nelle classi di decomposizione di Hunter; (5) il volume con la formula di Huber.
La CWD è stata stimata con un campionamento per intersezione lineare ([51], [15]). Il metodo presuppone che i pezzi siano approssimativamente cilindrici, giacenti sul terreno e che siano distribuiti e orientati casualmente nell’area. Il rilevamento è stato condotto lungo 9 segmenti lineari di campionamento di lunghezza di 30 m per ogni AdS. Con riferimento ad ogni singolo segmento è stata rilevata la lunghezza ed il diametro dei pezzi con diametro > 2.5 cm che intersecano il segmento. Il volume di ogni segmento lineare è stato calcolato con la seguente formula (eqn. 1):
dove m è il numero dei pezzi intersecati, d è il diametro dei pezzi (cm) nel punto di inserzione con il segmento, L è la lunghezza del segmento lineare campione (m).
Per il volume complessivo ad ettaro, tenuto conto che i segmenti sono di lunghezze diverse è stata utilizzata la seguente formula (eqn. 2):
dove n è il numero dei segmenti campione e Vha è la massa ad ettaro dei residui legnosi stimata sul j-esimo segmento campione.
Analisi della fruttificazione
La valutazione quantitativa della fruttificazione è stata fatta mediante una stima visiva della presenza/assenza di frutti sul totale della chioma in questo modo è stato possibile distinguere quattro classi: (1) assente (nessun frutto); (2) scarsa (la fruttificazione su <1/3 della chioma); (3) media (frutti da 1/3 a 2/3 della chioma); (4) Abbondante (frutti >2/3 della chioma). La valutazione è stata effettuata nel periodo di fruttificazione su tutte le piante comprese nelle AdS.
Analisi della rinnovazione naturale e della PAR (Photosintetic Active Radiation)
L’analisi dell’efficienza funzionale nei processi di rinnovazione naturale è un elemento fondamentale per valutare le potenzialità di perpetuazione di un popolamento. Si considera rinnovazione naturale l’insieme delle piante nate da seme con diametro < 2.5 cm a 1.30 m ([40]). L’efficienza di questo processo è stata valutata analizzando: (1) le modalità di rinnovazione (densità, distribuzione); (2) i fattori limitanti e di disturbo che impediscono o compromettono l’insediamento e l’affermazione della rinnovazione naturale.
La rinnovazione naturale è stata rilevata con un campionamento sistematico per allineamenti paralleli di aree campione ogni 10 m ([15]). Con riferimento ad ogni singolo allineamento è stata definita ogni 5 m una microarea di 1 m di raggio su cui effettuare i rilievi. Di ogni individuo sono stati rilevati: specie, altezza, età indicativa, localizzazione (coperto, scoperto, margine). Per ogni area è stato determinato l’indice di rinnovazione (IR = Hm’ D) di Magini ([36]).
La quantità e la qualità della luce che arriva ai livelli inferiori del popolamento influenza le condizioni microclimatiche e quindi la vegetazione del sottobosco, i tassi di decomposizione della sostanza organica, i processi di rinnovazione naturale. In ogni area sono state eseguite misure di PAR con un ceptometro (AccuPAR, Degagon Devices Inc., Pullman, WA, USA) a 1.5 m di altezza dal suolo con lo strumento in posizione orizzontale. Le misure sono state effettuate nei mesi di luglio, agosto e settembre, in giornate di cielo sereno, alle 12 (ora solare). Le misure di radiazione sono state eseguite: (1) all’esterno in condizioni di totale assenza di copertura forestale: (2) all’interno dell’area di saggio eseguendo 4 misurazioni secondo i punti cardinali.
Dai valori ottenuti da questi rilievi è stata calcolata la trasmittanza (T %), ossia la percentuale di radiazione che penetra al di sotto di una superficie più o meno continua rispetto a quella che incide al di sopra di essa ([42]).
Analisi fitosociologica
Il rilievo della vegetazione è stato eseguito secondo il metodo fitosociologico ([9]). La stima dell’indice di abbondanza-dominanza è stata effettuata secondo una scala convenzionale (5 = copertura > 75%; 4 = 50-75%; 3 = 25-50%; 2 = <25%; 1 = <5%; + = copertura molto scarsa). Gli strati di vegetazione considerati sono: (6) strato arboreo superiore, con altezza >25 m; (5) strato arboreo intermedio, 12-25 m; (4) strato arboreo inferiore, 5-12 m; (3) strato arbustivo superiore, 2-5 m; (2) strato arbustivo inferiore 0.5-2 m; (1) strato erbaceo. La nomenclatura delle specie segue Giardina et al. ([27]) e Brullo et al. ([12]) per il genere Quercus.
Analisi dei disturbi
L’analisi dei disturbi per stimare lo stato di degrado ha preso in considerazione gli impatti di natura antropica e quelli generati dal pascolo degli ungulati (domestici e selvatici). Gli impatti sono stati stimati qualitativamente secondo la scala temporale, spaziale e la magnitudo mediante la raccolta di indici di presenza/assenza di indicatori. Riguardo alla scala temporale è stata utilizzata la seguente classificazione: (1) temporaneo: il disturbo si verifica sporadicamente; (2) semi-permanente: il disturbo si verifica costantemente ma in modo irregolare; (3) permanente: il disturbo si verifica in modo permanente. Riguardo alla scala spaziale è stata utilizzata la seguente classificazione: (1) localizzato: il disturbo si verifica su piccole superfici; (2) non localizzato: il disturbo si verifica nell’area studio e nelle aree circostanti; (3) esteso: il disturbo si verifica nell’area studio ed in tutte le aree circostanti in modo indifferenziato.
Riguardo alla magnitudo è stata utilizzata la seguente classificazione: (1) bassa: il disturbo causa la modificazione delle componenti biotiche, abiotiche ed ecologiche in modo reversibile nel brevissimo periodo (1-2 mesi); (2) media: il disturbo causa la modificazione delle componenti biotiche, abiotiche ed ecologiche in modo reversibile nel lungo periodo; (3) alta: il disturbo causa la modificazione delle componenti biotiche, abiotiche ed ecologiche in modo irreversibile.
Per i disturbi antropici sono stati presi in considerazione i seguenti indici di presenza/assenza: (1) tagli abusivi; (2) raccolta della necromassa. Per gli ungulati sono stati raccolti indici di presenza/assenza: (1) escrementi; (2) danni da grufolamento e da calpestio; (3) brucature; (4) scortecciature; (5) fenomeni di costipazione del suolo.
Risultati e Discussione
Elementi dendrometrico-strutturali
Il Bosco Pomieri è risultato disomogeneo dal punto di vista strutturale e floristico, e rispecchia anche un diverso grado di degrado: le AdS poste a quote più basse (vicine alle strade ed alle abitazioni) sono molto più degradate e rispetto a quelle localizzate a quote più elevate (lontane da strade ed abitazioni). Anche i valori dei parametri dendrometrici (area basimetrica, volume) più bassi confermano questa condizione (Tab. 1).
Tab. 1 - Elementi dendrometrico-strutturali. (G): area basimetrica; (V): volume; (GC): grado di copertura; (a): età media in anni (tra parentesi valori minimo e massimo).
| AdS | n. piante(ha-1) | G ha-1(m2) | V ha-1(m3) | GC (%) | Età (a)(anni) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ceduo | Fustaia | Tot | |||||||
| Faggio | Rovere | Faggio | Rovere | ||||||
| 1 | - | 488 | - | 996 | 1484 | 25.7 | 197 | 100 | 160 (92-229) |
| 2 | - | 267 | - | 460 | 727 | 13.3 | 83 | 100 | 159 (102-217) |
| 3 | - | 228 | - | 144 | 372 | 10.5 | 104 | 96 | 93 (86-98) |
| 4 | - | 148 | - | 764 | 912 | 30.4 | 272.2 | 81 | 165 (135-195) |
| 5 | 336 | - | 240 | 40 | 616 | 33.4 | 827.3 | 93 | 140 (118-163) |
| 6 | - | - | - | 428 | 428 | 21.6 | 278.6 | 94 | 152 (123-185) |
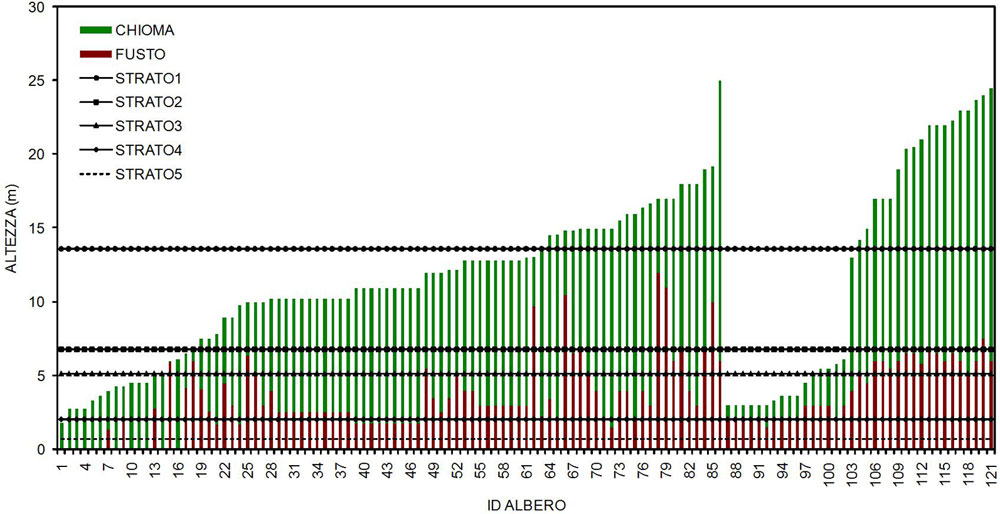
Per la tipologia QU10X le strutture riscontrate sono quella biplana e quella multiplana. L’indice di Latham et al. ([32]) ha messo in evidenza una struttura verticale in cinque strati (Fig. 3) ma, per scopi pratici, è opportuno distinguere tre strati principali: (1) strato dominante (Fig. 3- Strato 1: 13.6 m) formato dalle piante più vecchie che si distingue nettamente da uno o più strati dominati a seconda delle situazioni; (2) strato dominato (Fig. 3 - Strato 2: 6.8 m + Strato 3: 5.1 m), quando presente, comprende piante di rovere di origine gamica ed agamica che si distinguono per altezza ed età rispetto alle piante dominanti e sporadici soggetti di agrifoglio; (3) strato sotto dominato (Fig. 3 - Strato 4: 2 m + Strato 5: 0.7 m), maggiormente diffuso nell’area studio, comprende i soggetti di agrifoglio misto a giovani sporadici soggetti di rovere aduggiati dall’eccessiva copertura.
Fig. 3 - Grafici di stratificazione della struttura verticale secondo Latham et al. ([32]) per la tipologia QU10X.
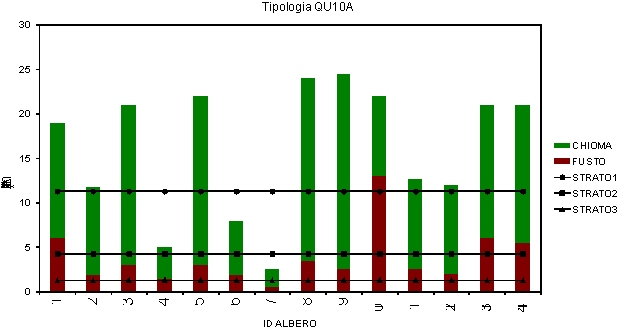
Per la tipologia FA10B l’indice di Latham et al. ([32]) ha messo in evidenza una struttura verticale in tre strati (Fig. 4), ma anche in questo caso per scopi pratici è opportuno distinguere due strati principali: (1) strato dominante (Fig. 4 - Strato 1: 11.3 m), che comprende le piante adulte di faggio e sporadici soggetti di rovere di origine sia gamica che agamica: (2) strato dominato (Fig. 4 - Strato 2: 4.3 m + Strato 3: 1.3 m), che comprende i soggetti di agrifoglio e la rinnovazione sporadica.
Fig. 4 - Grafici di stratificazione della struttura verticale secondo Latham et al. ([32]) per la tipologia QU10A.
La necromassa
La mortalità, riferita al numero di individui in piedi, è in media dell’8.1% (2.6-15 - Tab. 2). La mortalitàè dovuta essenzialmente a fattori di competizione tra gli individui, danni meteorici (vento e neve) e nel caso dell’AdS 4, alla presenza di un fungo patogeno Fomes fomentarius (L.) J.J. Kickx. La maggior parte delle piante morte in piedi appartengono alle classi più piccole comprendendo principalmente le classi da 3 a 13 cm (Tab. 3). Vi sono anche soggetti di maggiore diametro fino a 63 cm che presentano cavità alla base e lungo il fusto. Questi ultimi possono essere considerati degli “alberi habitat” in quanto vengono utilizzati come luoghi di rifugio e nutrimento da piccoli roditori, dagli uccelli e dagli insetti. Per quanto riguarda le classi di decomposizione prevalgono quelle delle fasi iniziali (2-4) ad eccezione dell’area 1 e 2 dove si hanno quelle tipiche delle fasi più avanzate della decomposizione (5-7 - Tab. 4). Il quantitativo di legno morto è molto variabile perché dipende dall’età, dal tipo di trattamento selvicolturale, dal grado di decomposizione, dal tipo e dall’intensità dei disturbi.
Tab. 2 - Mortalità degli individui in piedi.
| AdS | Vivi(n ha-1) | Morti (n ha-1) | Totale(n ha-1) | Morti in piedi (%) |
|---|---|---|---|---|
| 1 | 1484 | 164 | 1648 | 11.1 |
| 2 | 727 | 107 | 834 | 15 |
| 3 | 372 | 28 | 400 | 7.5 |
| 4 | 912 | 72 | 984 | 7.9 |
| 5 | 616 | 16 | 932 | 2.6 |
| 6 | 428 | 20 | 448 | 4.7 |
Tab. 3 - Ripartizioni in classi di diametro degli individui morti in piedi (%), diametro (cm).
| Diametro/AdS | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| 3 | 12.2 | 62.5 | - | 5.6 | - | 20 |
| 4 | 41.5 | 18.7 | 28.6 | - | - | - |
| 5 | 17.1 | 6.2 | 14.3 | 5.6 | - | 20 |
| 6 | 9.8 | 12.5 | - | 5.6 | 50 | 40 |
| 7 | 9.8 | - | - | 11.1 | - | - |
| 8 | 2.4 | - | - | - | - | 20 |
| 9 | 2.4 | - | 28.6 | 16.7 | - | - |
| 10 | 4.9 | - | 14.3 | 11.1 | - | - |
| 11 | - | - | - | 5.6 | 25 | - |
| 12 | - | - | - | 11.1 | - | - |
| 13 | - | - | 14.3 | - | 25 | - |
| 14 | - | - | - | 5.6 | - | - |
| 15 | - | - | - | 5.6 | - | - |
| 16 | - | - | - | 5.6 | - | - |
| 57 | - | - | - | 5.6 | - | - |
| 63 | - | - | - | 5.6 | - | - |
Tab. 4 - Ripartizione della SDT in classi di decomposizione di Hunter (%).
| Classe/AdS | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| 1 | - | 2.4 | - | 2.4 | 19.5 | 46.3 | 29.3 |
| 2 | - | - | 25 | 25 | 50 | - | - |
| 3 | - | 42.9 | 42.9 | 14.3 | - | - | - |
| 4 | - | 11 | 39 | 50 | - | - | - |
| 5 | - | - | 100 | - | - | - | - |
| 6 | - | 20 | 40 | 40 | - | - | - |
Il volume della necromassa in piedi è in media di 7.2 (0.6-20) m3 ha-1 (Tab. 5), mentre quello della necromassa a terra è in media di 1.7 (0.2-8.3) m3 ha-1 (Tab. 5). Tale valore è inferiore rispetto a quello rilevato nei popolamenti di querce (Quercus robur e Q. petraea) nel sud della Svezia (3.9 m3 ha-1 - [41]), nelle fustaie pluristratificate di cerro del Molise (12.8 m3 ha-1 - [37]), nei boschi di farnetto della Calabria (5.3 m3 ha-1 - [4]), nei boschi di cerro sia gestiti (4.5 m3 ha-1) che non gestiti (31.6 m3 ha-1) del Molise ([35]), nei cedui di cerro invecchiati dell’Italia centrale (21.8-40.3 m3 ha-1 - [7]).
Tab. 5 - Volume della necromassa.
| AdS | CWD(m3 ha-1) | SDT(m3 ha-1) | Totale(m3 ha-1) |
|---|---|---|---|
| 1 | 0.9 | 2.8 | 3.7 |
| 2 | 0.3 | 0.6 | 0.9 |
| 3 | 0.2 | 1.8 | 2 |
| 4 | 8.3 | 4.4 | 12.7 |
| 5 | 0.3 | 14.3 | 14.6 |
| 6 | 0.2 | 20 | 20.2 |
Il volume della necromassa totale (CWD e SDT) è in media di 9 m3 ha-1 (0.9- 20.2 - Tab. 5) quindi per la metà delle AdS al di sotto della soglia di 10 m3 ha-1 indicata come il valore minimo ([40]) per le foreste normalmente coltivate del Bacino del Mediterraneo.
Il quantitativo di necromassa aumenta spostandosi dalle AdS più disturbate (collocate a quote più basse, vicino alle strade ed alle abitazioni quindi dove si effettua la raccolta della legna) verso quelle meno disturbate (collocate a quote più alte).
La fruttificazione
Dalla stima visiva è emerso che la produzione di seme di rovere, nel 2008-2009, è stata classificata come scarsa nelle AdS 1, 2, 3 e 6 e abbondante nelle aree 4 e 5. Ciò è da attribuire principalmente alle maggiori dimensioni delle chiome dei soggetti nelle AdS 4 e 5 dove il seme si è presentato integro, ben sviluppato e potenzialmente fertile. La fruttificazione nell’area studio non è risultata mai assente nel periodo dello studio.
La rinnovazione naturale e la PAR
La rinnovazione naturale di rovere è presente in quattro AdS (1, 2, 3, 4), con densità≤ 5.5 semenzali m-2 e con un età media < 3 anni (Tab. 6). I semenzali difficilmente riescono a superare l’età di 5-6 anni; questa infatti è l’età limite oltre la quale i semenzali non riescono a sopportare l’aduggiamento ([5]).
Tab. 6 - Rinnovazione naturale. (C): coperto; (S): scoperto; (M). margine; (Q): rovere; (L): leccio.
| AdS | 1 | 2 | 3 | 4 | 5 | 6 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ubicazione | C | S | M | C | S | M | C | S | M | C | S | M | C | S | M | C | S | M | |
| Specie (ID) | Q | Q | Q | Q | Q | L | Q | Q | Q | Q | Q | Q | Q | - | - | - | - | - | - |
| Densità (n sem. m-2) | 4.6 | 5.7 | 2 | 4.1 | 7 | 1.5 | 7.6 | - | 2.5 | 1 | 5.5 | 1 | 4.3 | - | - | - | - | - | - |
| Diam. coll. medio (mm) | 2.4 | 1.5 | 2.2 | 2.1 | 1.1 | 0.4 | 2.5 | - | 2.3 | 2.5 | 2.6 | 1.2 | 1.5 | - | - | - | - | - | - |
| Altezza media (cm) | 8.2 | 7.7 | 5.5 | 8.3 | 10.5 | 5.5 | 8.3 | - | 6 | 4 | 4.8 | 4 | 3.1 | - | - | - | - | - | - |
| Età media (anni) | 2.1 | 2.3 | 2 | 1.8 | 3 | 2 | 1.8 | - | 2 | 1 | 1.5 | 2 | 1.1 | - | - | - | - | - | - |
| IR (Magini) | 37.7 | 43.9 | 11 | 34.3 | 73.5 | 5.2 | 63.1 | - | 15 | 4 | 26.4 | 4 | 13.3 | - | - | - | - | - | - |
La mancanza di rinnovazione è messa in evidenza dall’indice di rinnovazione di Magini ([36]). L’IR varia da 4 a 73.5 mentre i valori riportati per il Bosco di Sargiano (AR) da Cutini & Mercurio ([16]) sono > 1400.
L’insediamento della rinnovazione naturale è compatibile con i livelli di area basimetrica e volume, ad eccezione dell’AdS 5, che supera quei valori soglia riportati anche in precedenti lavori ([18], [16]).
Tuttavia la rinnovazione si concentra nelle zone più aperte, non solo all’interno del bosco ma anche lungo le scarpate stradali dove i semenzali si presentano in ottimo stato vegetativo.
Le cause della mancanza di rinnovazione sono da ricondursi a un’insieme di fattori: (1) scarsa fruttificazione; (2) al pascolo; (3) abbandono colturale.
I valori di trasmittanza variano da 0.6 a 8.6 % (Tab. 7) e indicano una condizione di scarsa illuminazione che caratterizza il bosco in esame e che inibisce lo sviluppo dei semenzali.
Tab. 7 - Valori di PAR e trasmittanza (media ± deviazione standard).
| ADS | PAR | T(%) | |
|---|---|---|---|
| Interno(µmol m-2 s-1) | Esterno(µmol m-2 s-1) | ||
| 1 | 47 (± 20.8) | 1813 (± 313.3) | 2.5 |
| 2 | 164 (± 123) | 1813 (± 313.3) | 8.6 |
| 3 | 37 (± 29.7) | 1813 (± 313.3) | 1.9 |
| 4 | 67 (± 51.7) | 1813 (± 313.3) | 3.6 |
| 5 | 11 (±2.6) | 1813 (± 313.3) | 0.6 |
| 6 | 19 (± 30) | 1813 (± 313.3) | 1.0 |
La rinnovazione è addirittura assente nelle AdS 5 e 6 dove i livelli di trasmittanza sono inferiori all’1 %. Infatti il punto di compensazione per la rovere è del 2% ([30]). [16], [17]) hanno rilevato che valori di trasmittanza <5 % non ostacolano l’insediamento della rinnovazione, ma lo sviluppo dei semenzali richiede valori del 50-80 % variabile in funzione della maggiore o minore umidità del terreno.
La vegetazione
I rilievi fitosociologici svolti nelle aree di saggio e riportati nella Tab. 8 evidenziano una formazione forestale nel complesso degradata, piuttosto povera floristicamente. Lo stato arboreo è dominato essenzialmente dalla rovere meridionale con valori di copertura quasi mai molto elevati, sporadica è la presenza di altre specie arboree. L’agrifoglio (Ilex aquifolium) è molto diffuso soprattutto nelle AdS 3 e 6, dove assume il significato di specie differenziale dell’associazione. Ben rappresentate sono altre specie sempreverdi come Hedera helix Ruscus aculeatus e Daphne laureola, che bene evidenziano la peculiarità di questo bosco misto con elementi sempreverdi laurifilli, tipica espressione relittuale preglaciale della fascia colchica ([43]). Nello strato erbaceo le specie nemorali mesofile tipiche delle formazioni forestali dei Querco-Fagetea sono scarsamente rappresentate, mentre assumono un certo rilievo alcune specie sinatropiche come Pteridium aquilinum, Urtica dioica e Chaerophyllum temulum e altre legate ad ambienti preforestali e pascolivi come Genista cupani, Sedum amplexicaule subsp. tenuifolium, Alyssum nebrodensis, ecc.
Tab. 8 - Rilievi fitosociologici.
| Codice area di saggio | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Quota (m s.l.m.) | 1310 | 1322 | 1316 | 1381 | 1434 | 1334 |
| Esposizione | N | N | NW | SE | E-SE | N |
| Superficie (m2) | 500 | 500 | 500 | 500 | 500 | 500 |
| Copertura totale (%) | 90 | 85 | 100 | 95 | 95 | 100 |
| Inclinazione (%) | 38 | 38 | 32 | 38 | 48 | 44 |
| Copertura strato arboreo (%) | 85 | 80 | 75 | 95 | 90 | 90 |
| Copertura strato arbustivo (%) | 15 | 10 | 5 | 10 | 5 | 10 |
| Copertura strato erbaceo (%) | 10 | 5 | 5 | 20 | 5 | 1 |
| Strato Arboreo | ||||||
| Quercus petraea (Mattuschka) Liebl. subsp. austrotyrrhenica Brullo et al. | 4 | 4 | 4 | 5 | 1 | 5 |
| Quercus ilex L. | 1 | - | - | - | - | - |
| Fagus sylvatica L. | - | - | - | - | 4 | + |
| Strato Arbustivo | ||||||
| Quercus petraea (Mattuschka) Liebl. subsp. austrotyrrhenica Brullo et al. | - | - | + | + | . | - |
| Fagus sylvatica L. | - | - | - | - | 1 | - |
| Hedera helix L. | 2 | 2 | 1 | 2 | + | 1 |
| Ilex aquifolium L. | 2 | 2 | 3 | 1 | + | 3 |
| Ruscus aculeatus L. | 1 | + | + | 1 | + | + |
| Daphne laureola L. | 2 | 1 | . | 2 | 2 | . |
| Crataegus monogyna Jacq. subsp. monogyna | + | - | 1 | - | + | . |
| Rubus hirtus Waldst. & Kit. | + | + | + | - | - | + |
| Strato Erbaceo | ||||||
| Doronicum orientale Hoffm. | 2 | 1 | - | 1 | - | - |
| Trifolium pratense L. subsp. semipurpureum (Strobl) Pignatti | + | + | - | - | - | - |
| Euphorbia meuselii Mazzola & Raimondo | + | - | - | - | + | - |
| Crepis leontodontoides L. | 2 | 1 | + | 1 | - | + |
| Genista cupani Guss. | + | - | - | + | - | - |
| Cyclamen hederifolium Aiton | - | - | - | 1 | 1 | + |
| Fragaria vesca L. | + | + | - | - | - | - |
| Quercus petraea (Mattuschka) Liebl. subsp.austrotyrrhenica Brullo et al. | 2 | 1 | + | + | - | - |
| Acer campestre L. | . | . | + | . | - | - |
| Ranunculus velutinus Ten. | 2 | 1 | + | + | - | - |
| Sedum amplexicaule DC. subsp. tenuifolium (Sm.) Greuter | - | - | - | + | - | - |
| Chaerophyllum temulum L. | - | - | 1 | + | - | + |
| Urtica dioica L. | 1 | 1 | 1 | 1 | - | + |
| Teucrium scorodonia L. subsp. crenatifolium (Guss.) Arcang. | - | - | - | + | - | - |
| Alyssum nebrodense Tineo | - | - | - | + | - | - |
| Pteridium aquilinum (L.) Kuhn | 1 | 1 | + | + | - | 1 |
| Quercus ilex L. | + | + | . | + | - | . |
| Aquilegiavulgaris L. | + | - | - | + | - | + |
| Asparagus acutifolius L. | 2 | 1 | 1 | 2 | - | - |
I fattori di disturbo
Le attività antropiche hanno un ruolo primario sulla funzionalità degli ecosistemi, le informazioni raccolte indicano un impatto differenziato: (1) nelle AdS poste a quote più basse vicino alle strade ed alle abitazioni il disturbo considerato è stato classificato come temporaneo, non localizzato e di magnitudo alta; (2) nelle AdS poste a quote più elevate lontano dal strade ed abitazioni il disturbo è stato classificato come semi-permanente, localizzato e di magnitudo bassa.
Gli impatti causati dalla presenza di ungulati hanno un ruolo sulla composizione e sulla funzionalità del bosco, in generale, essi sono la causa principale dell’impoverimento della composizione floristica, dell’erosione del suolo e del danneggiamento della rinnovazione naturale e della presenza di specie sinantropiche quali Urica dioica e Pteridium aquilinum, rilevate in tutte le aree di saggio.
Gli indici di presenza raccolti nelle AdS hanno indicato: un elevato numero di escrementi; la presenza di aree grufolate con evidenti segni di calpestio; la presenza di rinnovazione danneggiata; danni da morso. Queste informazioni hanno consentito di classificare il disturbo secondo le tre caratteristiche principali: scala temporale (frequenza), scala spaziale e magnitudo. Il pascolo degli ungulati domestici e selvatici si presenta come un disturbo semi-permanente secondo la scala temporale, diffuso secondo la scala spaziale e di intensità alta secondo la magnitudo (Tab. 9).
Tab. 9 - Sintesi dei disturbi dell’area studio in relazione alle caratteristiche temporali, spaziali e di magnitudo. (*): AdS poste a quote inferiori prossime alla strada ed alle abitazioni; (**): AdS poste a quote superiori lontane da strade e dalle abitazioni.
| Disturbo | Tipologia di disturbo in relazione a: | ||
|---|---|---|---|
| Tempo | Spazio | Magnitudo | |
| Disturbi antropici | Temporaneo (*) Semi-Permanente (**) | Non Localizzato (*) Localizzato (**) | Alta (*)Bassa (**) |
| Pascolo di ungulati | Semi- Permanente | Esteso | Alta |
Ciò indica che gli impatti generati sono significativi ovvero che i disturbi possono avere delle incidenze che richiedono la predisposizione di opportune misure di mitigazione.
Considerazioni conclusive
Le analisi condotte concorrono a creare la base conoscitiva per avviare un’azione di restauro forestale. Essi hanno messo in evidenza una scarsa funzionalità dell’ecosistema, più marcata nelle aree poste a quote più basse rispetto a quelle poste a quote più elevate.
Il restauro è pienamente giustificato sul piano etico trattandosi di un bosco di elevato interesse scientifico, ubicato all’interno di un’area protetta dove il rischio di estinzione della popolazione di rovere è elevato ([14]).
Sul piano applicativo il restauro presupporrà sempre l’intervento dell’uomo per ripristinare la funzionalità del sistema, dal momento che si tratta di sistemi molto degradati. In concreto, si articolano una serie di misure preliminari per rimuovere quei fattori di disturbo che al momento sono i responsabili dello stato di degrado, ossia: (1) divieto del pascolo di animali domestici; (2) divieto di raccolta di legna da ardere e di accesso ai non addetti alla gestione.
Dopo qualche anno sarà necessario intervenire con azioni selvicolturali mirate nelle diverse tipologie strutturali-funzionali che in breve si posso riassumere:
- conversione attiva in fustaia dei soprassuoli rappresentati dalle AdS 1, 2, 3, mediante periodici diradamenti, rispettando la configurazione strutturale del soprassuolo, senza ancorarsi a schemi numerici rigidi, fino ad arrivare al taglio di conversione vero e proprio una volta che il sistema suolo-soprassuolo ha raggiunto la maturità;
- controllo della diffusione dell’agrifoglio, per favorire la rinnovazione naturale della rovere, che come è noto si riproduce in modo straordinario anche per polloni radicali;
- apertura di piccole buche nei soprassuoli adulti e densi per favorire la rinnovazione di rovere: raggio medio delle buche uguale all’altezza degli alberi vicini secondo Bary-Lenger & Nebout ([5]). Anche Johnson et al. ([31]) sono dell’avviso che il gap di rinnovazione deve essere sufficientemente ampio in modo tale che la copertura delle chiome degli alberi all’intorno si chiuda dopo alcuni anni, non prima che la rinnovazione si sia affermata;
- valorizzare il ruolo dei grandi alberi, tenendo presente che queste formazioni hanno un tempo di permanenza molto lungo, almeno 500 anni ([19]). Essi oltre ad essere di grande pregio estetico e di richiamo turistico, sono molto importanti dal punto di vista biologico: gli alberi senescenti e cavi sono utilizzati da uccelli, roditori, piccoli mammiferi come luogo di rifugio e alimentazione ([25]);
- salvaguardia delle specie arboree consociate (aceri, ecc.);
- monitoraggio delle dinamiche evolutive, soprattutto nelle zone di tensione verso la faggeta e verso il basso anche in relazione ai previsti cambiamenti climatici.
Il principio ispiratore degli interventi futuri sarà quello di assecondare e guidare le dinamiche evolutive naturali, sempre sotto il costante controllo da parte dell’uomo.
Questa azione per completarsi richiederà un arco temporale abbastanza lungo, presumibilmente 20-30 anni (e non solo 5 anni come previsto dal Piano di Gestione Monti Madonie 2008 per la zona A del Parco Regionale). Questo potrebbe sembrare incompatibile e impraticabile sul piano operativo, ma queste sono le leggi della natura da rispettare se si vogliono raggiungere gli obiettivi del restauro. La politica ambientale a favore della biodiversità richiede, al di là delle solenni affermazioni di principio (come quelle dell’anno della biodiversità 2010), azioni concrete. Questa è una forma di conservazione in situ ([6], [2]) secondo quanto previsto anche dalla Convenzione per la Conservazione della Biodiversità. E questo potrebbe essere un esempio. Più che un problema tecnico si pone semmai un problema finanziario e organizzativo. Al riguardo, in Italia non sono mancati esempi negativi, vedi la rete delle principali formazioni forestali sottratte all’azione antropica creata da Aldo Pavari negli anni ’50 e ora quasi smantellata per passaggi di competenze tra enti, per mancanza di fondi e non ultima per ignoranza da parte di coloro che avrebbero dovuto garantirne una gestione efficace.
References
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar