Comparison among methods for the assessment of deadwood volume in a former holm oak coppice
Forest@ - Journal of Silviculture and Forest Ecology, Volume 10, Pages 34-42 (2013)
doi: https://doi.org/10.3832/efor0795-010
Published: Apr 15, 2013 - Copyright © 2013 SISEF
Research Articles
Abstract
Comparison among methods for the assessment of deadwood volume in a former holm oak coppice. The paper aims to compare three methods for the assessment of deadwood volume, i.e., LIS (Line Intersect System), FAS (Fixed Area Sampling), and WM (Weighings Method). The control data are represented by the outputs of Xylometric measurement. The study was carried out in a former holm oak (Quercus ilex L.) coppice located in the nature park of Montioni in southern Tuscany. LIS and FAS overestimated significantly the quantity of deadwood (+12% and +50%, respectively). The error become higher as the minimum threshold sampling increases. The WM, besides the operational complexity of its application, led to more promising and precise results.
Keywords
Deadwood, Sampling Methods, Line Intersect System, Fixed Area Sampling, Mediterranean Coppice
Introduzione
Negli ultimi anni numerosi studi hanno accertato il ruolo fondamentale della necromassa legnosa nel complesso funzionamento dell’ecosistema foresta. Il legno morto svolge un ruolo significativo nel sequestro del carbonio [13], [20], [25], [14], [32], [31]), ha un ruolo determinante nei processi di formazione dell’humus ([9], [18], [23]) e di apporto di sostanze nutritive ([11]), incrementando la produttività complessiva della foresta ([38]). Inoltre non è da trascurare l’azione positiva in termini di offerta di habitat per specie animali e vegetali, contribuendo così alla conservazione della diversità biologica ([13], [24], [2], [36], [17], [27], [1], [16], [29], [28]). La necromassa legnosa rappresenta inoltre uno dei parametri determinanti per la caratterizzazione dei boschi vetusti ([26], [6]).
Alla luce di quanto sopra accennato, appare chiaro che la quantificazione della necromassa legnosa diviene essenziale per caratterizzare l’ecosistema forestale.
Ad oggi esistono, e sono applicate, svariate metodologie di stima del legno morto in foresta ([42]), metodologie che sono state nel tempo messe a punto al fine di rendere i rilievi di più semplice e rapida esecuzione, anche se non è sempre ben chiaro il grado di affidabilità.
L’obiettivo di questo lavoro, realizzato in concomitanza con il progetto PRIN 2007 “Metodi innovativi per l’identificazione, caratterizzazione e gestione dei boschi vetusti in ambiente mediterraneo”, è stato il confronto tra alcune delle più utilizzate metodologie di stima del volume del legno morto a terra. Tale confronto è stato condotto in un ceduo invecchiato di leccio (Quercus ilex L.) localizzato, così come l’area utilizzata per il progetto PRIN sopracitato, all’interno del Parco Interprovinciale di Montioni (GR-LI).
L’area di studio
L’area di studio è stata individuata all’interno della Riserva Statale di Popolamento Animale “Marsiliana”, compresa all’interno del Parco Interprovinciale di Montioni. La riserva si estende su 443 ha ricadenti nei comuni di Massa Marittima, Follonica e Suvereto nella parte centro-occidentale della Provincia di Grosseto.
Il substrato geologico è caratterizzato da elevata eterogeneità; prevalgono però formazioni Eoceniche a calcari argillosi e formazioni del Miocene con tracce di travertino. Da tali substrati si sono originati litosuoli, suoli bruni calcarei e suoli mediterranei bruni o rossi ([5]).
Il clima è tipicamente mediterraneo, caratterizzato da inverni miti e umidi ed estati calde e siccitose. Facendo riferimento alla stazione termo-pluviometrica di Suvereto (LI), la piovosità media annua è di 760 mm, con minimo estivo (luglio: 19.2 mm) e un picco autunnale (ottobre: 106 mm), la media del trimestre estivo è pari a 91 mm di pioggia. La temperatura media annuale risulta di 15.3 °C, il mese più caldo è agosto con la temperatura media massima di 29 °C ([5]).
Il territorio è collinare e si articola in superfici agricole alternate a cedui di sclerofille sempreverdi con dominanza di leccio (Quercus ilex L.) e, nelle zone meno aride, di cerro (Quercus cerris L.). Sui rilievi e nelle aree distinte da minor fertilità, la fisionomia dei boschi è quella della macchia mediterranea (macchia alta), caratterizzata dall’elevata ricchezza specifica; si ritrovano, infatti, leccio (Quercus ilex L.), corbezzolo (Arbutus unedo L.), eriche (Erica spp.) e molti altri arbusti xerofili ([37]).
Il soprassuolo oggetto d’indagine è stato oggetto di un rilievo preliminare per la caratterizzazione della struttura nella primavera 2009. Il popolamento è risultato costituito da un ceduo invecchiato di leccio, con presenza di cerro e orniello nelle zone più fresche dell’area. Si ritrovano inoltre, sporadicamente, roverella e olmo. L’età dei polloni, stimata attraverso l’esame dei dati storici delle autorizzazioni di taglio, è di circa 50 anni. Sono presenti matricine di leccio e cerro di ragguardevoli dimensioni e di età valutabile tra i 90 e i 110 anni, in quanto matricine di 2-3 turni del vecchio ceduo ([5]).
La copertura delle chiome è colma e la densità dei fusti piuttosto elevata. Il numero delle piante risulta di 2460 ha-1; densità alla quale corrisponde un’area basimetrica di 34.2 m2 ha-1. Il diametro medio del popolamento è di 13 cm, mentre l’altezza media è pari a 13 m. Il volume stimato è pari a 227.1 m3 ha-1 ([4]).
Materiali e metodi
Nel presente lavoro sono state considerate alcune delle metodologie proposte per il campionamento della necromassa legnosa a terra ([43], [10], [41]) nelle quali la precisione di stima è in parte legata alle caratteristiche di forma, dimensione e stato di decomposizione del materiale legnoso. I metodi più comunemente impiegati si basano sull’utilizzo di aree fisse di campionamento (FAS, Fixed Area Sampling - [12]) oppure di transetti (LIS, Line Intersect System - [44], [43], [22]) e forniscono entrambi come risultato un valore espresso in volume.
Nel presente lavoro sono stati messe a confronto le metodologie LIS, FAS e della determinazione indiretta del volume calcolato come rapporto tra peso e massa volumica (metodo delle “pesate”), utilizzando come valore di riferimento quello ottenuto con il metodo xilometrico mediante immersione in acqua.
I rilievi della necromassa legnosa sono stati effettuati nella primavera del 2010, in un’area di saggio quadrata, con lato di 50 m, suddivisa in 5 sottoaree rettangolari (10 x 50 m), individuate con lettere dalla A alla E e disposte perpendicolarmente alla linea di massima pendenza, e con un gradiente altimetrico di circa 20 m fra le sottoaree estreme. L’area di saggio è stata scelta in maniera soggettiva sulla base dei seguenti criteri: rappresentatività della struttura del popolamento, assenza di disturbo da parte degli ungulati (recinzione dell’area), aspetti logistici connessi all’uso del metodo xilometrico.
L’applicazione della metodologia FAS ha previsto la misurazione di tutti frammenti legnosi a terra, qualora questi presentassero un diametro superiore a 5 cm (FAS 5) per una lunghezza minima di 1 m. Inoltre, in 3 delle 5 sottoaree si è proceduto al rilievo abbassando la soglia diametrale a 3 cm (FAS 3).
Per la determinazione del volume di ciascun frammento, al fine di mantenere l’omogeneità dei dati con quelli raccolti nel progetto PRIN, è stata applicata la formula già utilizzata per i rilievi del progetto ([8]) così come anche indicato da Harmon & Sexton ([12] - eqn. 1):
dove V è il volume, h l’altezza/lunghezza, R è il raggio maggiore, r il raggio minore. I singoli volumi sono stati poi sommati a livello del plot e quindi riportati ad ettaro. Sono state misurate inoltre anche le piante intere morte a terra.
Il volume di ogni albero è stato stimato mediante l’utilizzazione delle tavole di cubatura a doppia entrata per i cedui di specie quercine ([7]), così come previsto nella metodologia del progetto PRIN.
La seconda metodologia impiegata è stata quella LIS, ovvero mediante transetti lineari di 50 m di lunghezza ciascuno. Tali transetti sono stati realizzati nelle 3 sottoaree dove si era proceduto al campionamento con la metodologia FAS, utilizzando soglie diametriche di 3 e 5 cm. In ogni sottoarea sono stati tracciati 3 transetti, paralleli al lato maggiore, alle distanze pre-stabilite di 3, 5 e 8 m dal lato basso al fine di condurre valutazioni sull’intensità di campionamento.
Dopo aver steso la rotella metrica attraverso tutta la sottoarea, si è proceduto alla misurazione del diametro di ogni frammento legnoso nel punto in cui veniva intercettato.
In particolar modo sono stati campionati tutti i frammenti legnosi che misurassero almeno 3 cm di diametro nel punto in cui erano intercettati dalla linea individuata al suolo.
Una volta misurati tutti i frammenti legnosi, si è provveduto al calcolo del volume ad ettaro fornito da ogni singolo transetto mediante l’applicazione della seguente formula ([43] - eqn. 2):
dove di è il diametro (in cm) dell’i-esimo ramo nel punto d’intersezione con il segmento lineare campione, n è il numero totale dei rami intersecati dai segmenti lineari campione, L è la lunghezza totale del segmento.
La terza metodologia impiegata, una volta terminati i rilievi LIS e FAS, è stata quella delle “pesate” ([34], [35]) applicata in tre parcelle di 10 m x 25 m individuate casualmente in ognuna delle 3 sottoaree nelle quali era stata applicata la metodologia LIS. L’applicazione del metodo delle “pesate” ha previsto la raccolta di tutta la necromassa legnosa presente, escludendo quella con diametro di base inferiore a 3 cm, suddividendola in due classi che utilizzavano come soglia discriminante quella di 5 cm. La necromassa così suddivisa è stata pesata utilizzando una bascula modello SIAP con portata di 50 Kg e approssimazione a 100 gr.
Qualunque sia stata la metodologia adottata, è stata annotata la classe di decomposizione di ciascun frammento legnoso o pianta morta a terra facendo riferimento alla classificazione di Hunter ([19]), comunemente adottata in questo tipo di lavori.
Nell’ambito delle due classi diametriche è stato prelevato un campione rappresentativo sia della distribuzione dimensionale che dello stato medio di decomposizione per la determinazione del peso e del volume. La scelta del materiale, a parità di dimensioni e classe di decomposizione, è stata fatta in maniera soggettiva. Il campione è stato chiuso in buste di plastica e le misure sui campioni freschi sono state eseguite nei laboratori del DEISTAF nei due giorni successivi al prelievo.
Ciascun pezzo è stato pesato insieme alla busta di plastica, di peso noto, in modo tale da tener conto anche dell’acqua di condensazione sulla pareti delle buste. Successivamente, dopo aver provveduto alla completa imbibizione di ogni pezzo, si è proceduto alla determinazione del volume attraverso l’immersione di ogni singolo frammento in un contenitore riempito con acqua e la successiva misura del peso dell’acqua spostata.
Dopo circa una settimana di permanenza in un locale areato a temperatura ambiente, i singoli pezzi sono stati essiccati, fino al raggiungimento del peso costante, utilizzando una stufa a ventilazione forzata alla temperatura di 60°. Per tutte le pesate in laboratorio è stata utilizzata una bilancia Mettler P4400 con precisione al centesimo di grammo.
Dello stesso materiale utilizzato in bosco per il metodo delle pesate è stato ricavato il volume esatto attraverso l’utilizzazione di due xilometri della capacità di 300 litri ciascuno, nei quali la necromassa raccolta è stata immersa.
Tutti i valori di volume registrati sono stati poi riportati ad ettaro.
Il volume della necromassa a terra è stato ottenuto moltiplicando il peso fresco ottenuto in bosco per il rapporto tra volume e peso fresco ricavati in laboratorio.
I dati sono stati elaborati con i comuni test di statistica inferenziale. In particolare per saggiare le differenze fra i valori medi ottenuti con le diverse soglie del LIS è stato eseguito il test ANOVA ponendo come fonti di variazione le soglie stesse utilizzando i dati dell’intero transetto (50 m), mentre per confrontare i valori dei diversi metodi di campionamento sono stati considerati solo i dati che fanno riferimento alle stesse superfici.
Risultati
Assumendo come dato di riferimento quello ottenuto con il metodo xilometrico (Tab. 1), l’area di studio dimostra al suo interno una notevole variabilità nella distribuzione sia quantitativa, che di dimensioni, della necromassa.
Tab. 1 - Quantità di necromassa (m3 ha-1) misurata nelle sub-aree com metodo xilometrico.
| Sub-area | Vol (m3 ha-1) | |
|---|---|---|
| Soglia 3 cm | Soglia 5 cm | |
| A | 11.11 | 1.72 |
| C | 33.56 | 21.86 |
| E | 17.77 | 9.04 |
| Media | 20.81 | 10.88 |
Nella sub-area C, risulta presente al suolo una quantità di necromassa quasi doppia rispetto alla sub-area più a monte e tripla nel confronto con quella più a valle. Prendendo come soglia di misura minima quella di 5 cm, il rapporto tra la necromassa presente nella sub area C rispetto alla A sale a oltre 12 e a poco più di 2 rispetto all’area più a monte.
Le differenze sono dovute alla quantità di necromassa minuta, compresa tra 3 e 5 cm, che rappresenta in volume circa l’85% nella sub-area più a valle, scende mediamente al 35% in quella mediana per poi risalire a quasi il 50% a valle nell’area E.
I campioni analizzati in laboratorio per il metodo delle “pesate”, sono risultati caratterizzati da una densità, calcolata sul rapporto peso secco/volume, che tende a diminuire con l’aumentare del livello di decomposizione, e risulta ovviamente più elevata qualora si consideri la soglia dimensionale superiore (Tab. 2).
Tab. 2 - Dati riassuntivi della densità (g cm-3), del rapporto peso secco/peso fresco (Ps/Pf) e del rapporto peso fresco/volume (Pf/Vol) dei campioni analizzati in laboratorio.
| Classe decomposizione |
Classe di diametro |
N. osservazioni |
Densità (g cm-3) |
Ps/Pf | Pf/Vol. |
|---|---|---|---|---|---|
| 1 | < 5 cm | 9 | 0.45 ± 0.06 | 0.73 ± 0.08 | 0.62 ± 0.10 |
| > 5 cm | 8 | 0.59 ± 0.05 | 0.69 ± 0.04 | 0.84 ± 0.05 | |
| 2 | < 5 cm | 14 | 0.45 ± 0.08 | 0.81 ± 0.09 | 0.56 ± 0.13 |
| > 5 cm | 6 | 0.62 ± 0.07 | 0.84 ± 0.07 | 0.74 ± 0.13 | |
| 3 | < 5 cm | 20 | 0.44 ± 0.08 | 0.81 ± 0.05 | 0.54 ± 0.13 |
| > 5 cm | 23 | 0.51 ± 0.09 | 0.82 ± 0.08 | 0.61 ± 0.12 | |
| 4 | < 5 cm | 12 | 0.34 ± 0.09 | 0.80 ± 0.08 | 0.42 ± 0.10 |
| > 5 cm | 7 | 0.35 ± 0.12 | 0.80 ± 0.09 | 0.44 ± 0.17 | |
| 5 | < 5 cm | 0 | - | - | - |
| > 5 cm | 0 | - | - | - |
Il rapporto peso secco/peso fresco, come mostrato in tabella, non presentava invece grosse variazioni sia se relazionato alla classe di diametro che a quella di decomposizione.
Nella medesima Tab. 2 è riportato anche il rapporto peso fresco/volume dei campioni analizzati.
Prescindendo dalla classe di decomposizione, i valori medi per le due soglie diametrali sono risultati rispettivamente 0.51 e 0.52 e i loro reciproci sono stati successivamente utilizzati per ricavare il volume della necromassa pesata in bosco.
I risultati dell’applicazione del metodo delle pesate, espressi in termini di volume (Tab. 3), risultano molto simili a quelli ricavati con l’uso dello xilometro.
Tab. 3 - Volume della necromassa (m3 ha-1) ottenuto con il metodo delle “pesate” alle due diverse soglie di campionamento.
| Sub-Area | Vol (m3 ha-1) | |
|---|---|---|
| Soglia 3 cm | Soglia 5 cm | |
| A | 11.01 | 1.67 |
| C | 33.07 | 21.33 |
| E | 18.04 | 9.05 |
Le differenze percentuali tra le due metodologie, da imputare a errori accidentali, oscillano tra lo 0.5 e l’1.5% per la soglia di 3 cm e tra lo 0.1 e il 3.4% per la soglia di 5 cm.
In Tab. 4 sono riportati i dati relativi alla stima della necromassa effettuata con la metodologia LIS alle due diverse soglie di campionamento.
Tab. 4 - Volume della necromassa (m3 ha-1) ottenuto con la metodologia LIS alle due diverse soglie di campionamento.
| Sub-area | transetto | LIS 3 | LIS 5 |
|---|---|---|---|
| A | 3m | 64.79 | 58.97 |
| 5m | 50.58 | 45.89 | |
| 8m | 50.73 | 44.46 | |
| C | 3m | 20.28 | 12.53 |
| 5m | 25.76 | 16.14 | |
| 8m | 15.5 | 10.36 | |
| E | 3m | 17.77 | 9.28 |
| 5m | 19.59 | 8.09 | |
| 8m | 13.32 | 1.78 | |
| Media (m3 ha-1) | 30.9 | 23.1 | |
| Errore Standard | 6.4 | 6.9 | |
Il primo risultato che emerge con grande evidenza riguarda le differenze di stima, a parità di soglia di campionamento, nell’ambito di ciascuna sub-area al variare della posizione della linea di rilievo dal vertice di riferimento.
Inoltre, anche a prescindere dalla soglia di campionamento e dalla posizione del transetto, l’applicazione della metodologia LIS non porta ad una corretta rappresentazione della differente distribuzione della necromassa nell’area sperimentale: nella sub-area A risulterebbe presente una quantità di necromassa da doppia (LIS 3) a quasi quadrupla (LIS 5) di quella stimata nella sub-area C, mentre le misure del testimone di riferimento mettono in evidenza una situazione reale esattamente opposta.
I dati ricavati con la metodologia LIS confrontati con quelli ottenuti con il metodo xilometrico, evidenziano una sovrastima generalizzata. Il confronto fra le medie delle tre sub-aree, fa oscillare la sopra valutazione tra il 49% (LIS 3) e il 112% (LIS 5). Tale sovrastima è notevolmente più accentuata nell’area A in presenza di una minore quantità di necromassa e, nello stesso tempo, di una maggiore percentuale di quella più minuta.
Va ricordato che il metodo viene diffusamente impiegato nella pratica perché considerato come scarsamente influenzato da possibili errori da parte del rilevatore ([33]) e che per tali motivi è anche stato impiegato come termine di paragone in lavori scientifici volti a valutare metodi diversi per la stima della necromassa ([21]).
I risultati derivanti dall’applicazione della metodologia FAS, sono riportati nella Tab. 5. Così come rilevato per la metodologia LIS, anche le stime effettuate con la FAS hanno portato ad differenze rilevanti nella stima della distribuzione della necromassa all’interno dell’area, sovrastimando quella presente nella sub-area più a monte e sottostimando quella nelle altre due sub aree.
Tab. 5 - Volume della necromassa (m3 ha-1) ottenuto con la metodologia FAS alle due diverse soglie di campionamento.
| Sub-Area | Vol (m3 ha-1) | |
|---|---|---|
| FAS 3 | FAS 5 | |
| A | 33 | 32.2 |
| C | 11.3 | 9.4 |
| E | 10.5 | 7.8 |
| Media | 18.3 | 16.5 |
In media, le stime effettuate con la metodologia FAS, porterebbero ad una quantità di necromassa superiore del 12% (FAS 3) e del 50% (FAS 5) rispetto alla reale xilometrica ma comunque nettamente inferiore rispetto a quella valutata con la metodologia LIS.
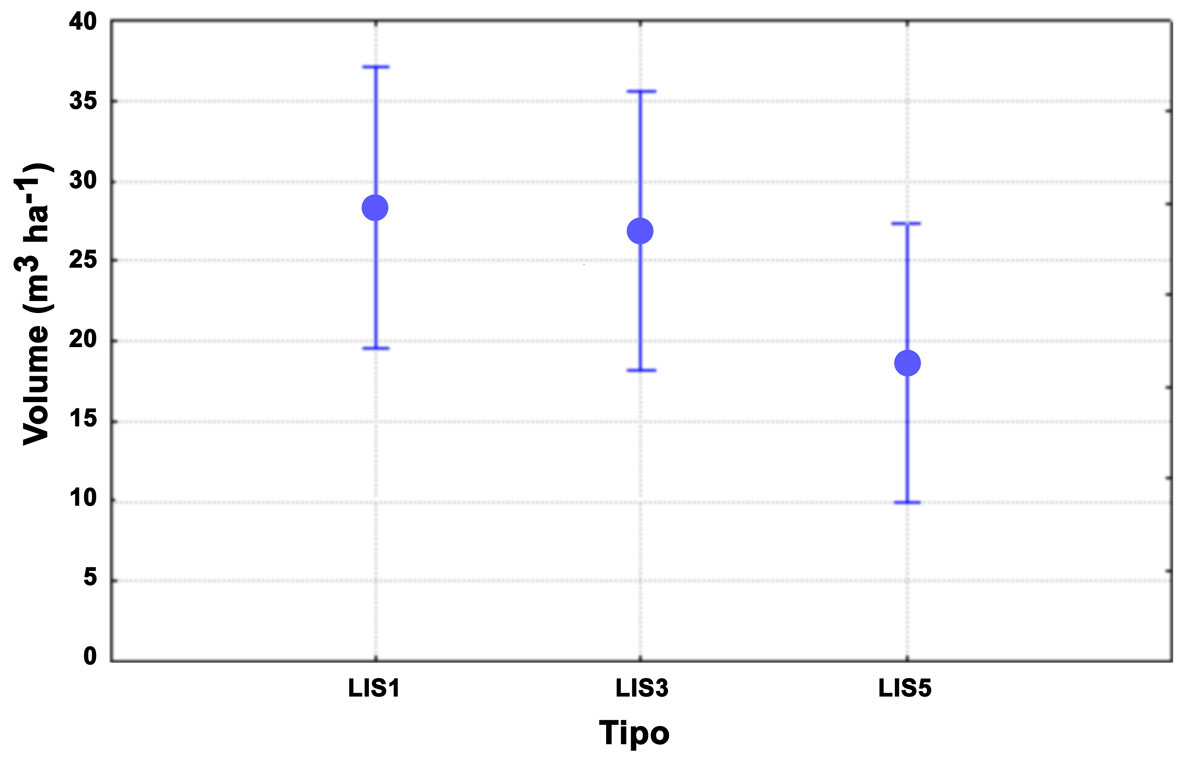
Nell’istogramma di Fig. 1 sono messi a confronto i risultati ottenuti attraverso l’applicazione delle diverse metodologie con differenze che non risultano statisticamente significative (Tab. 6).
Fig. 1 - Confronto tra le soglie di campionamento utilizzate con la metodologia LIS per il sito di Marsiliana. Le barre verticali rappresentano gli intervalli di confidenza (95%).
Tab. 6 - Confronto statistico (analisi della varianza) tra i risultati delle differenti metodologie di campionamento.
| Metodo | Vol (m3 ha-1) | ANOVA F (gl: 5, 24) |
|---|---|---|
| FAS5 | 16.5 | 0.49 ns |
| FAS3 | 18.3 | |
| LIS5 | 23.1 | |
| LIS3 | 30.9 | |
| PESO | 20.7 | |
| XILO | 20.8 |
Teissier du Cros & Lopez ([40]), confrontando FAS e LIS, hanno confermato che il metodo LIS è più economico (in termini di tempo necessario) e che la precisione aumenta all’aumentare della lunghezza del transect lineare; hanno inoltre messo in luce che i transect più lunghi testati (50 m) sono caratterizzati da un coefficiente di variazione di valore elevato e suggeriscono l’impiego di transect di lunghezza ancora maggiore. Gli stessi autori propongono inoltre di affiancare al LIS la stima con il FAS della componente in piedi (ceppaie, e alberi morti in piedi).
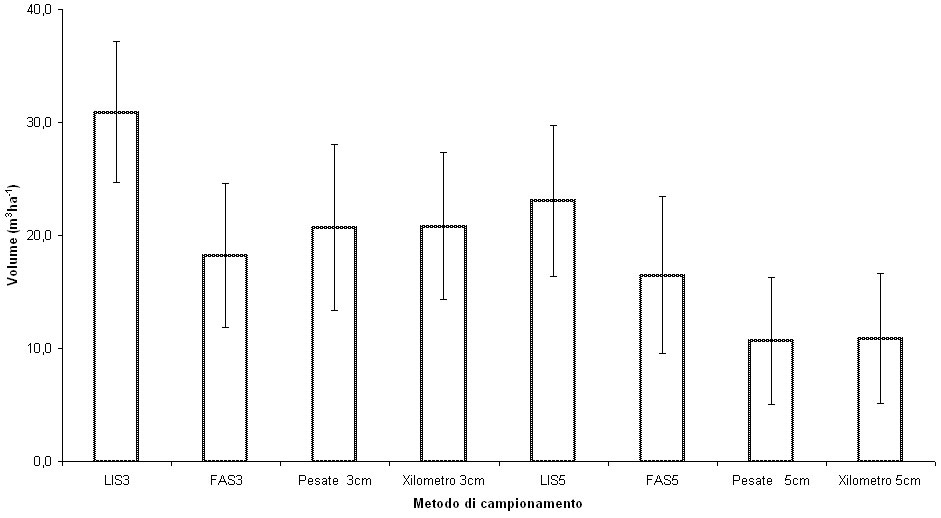
Woldendorp et al. ([45]) hanno osservato che il metodo FAS è da preferirsi nei soprassuoli caratterizzati da una scarsa quantità di necromassa (Fig. 2) e da grande variabilità nella sua distribuzione; nei casi dove invece è più opportuno applicare il metodo LIS si ottengono risultati migliori con transect di lunghezza pari almeno a 100 m.
Fig. 2 - Valori medi di necromassa (± e.s.) ottenuti con le differenti metodologie di campionamento.
Altri studi che mettono a confronto diversi metodi di campionamento ([30], [15], [39], [33], [3]) hanno comunque evidenziato che non è possibile individuarne uno più efficace in assoluto dato che i fattori topografici e le passate forme di gestione del soprassuolo hanno un’influenza significativa sul risultato del rilievo; in ogni caso diventa fondamentale la scelta da parte dell’operatore dei punti di campionamento ([21]).
Conclusioni
Riconosciuta l’importanza della quantificazione della necromassa legnosa presente al suolo, il confronto tra le diverse metodologie di rilievo applicate nel caso di studio, ha messo in luce differenze tali da richiedere ulteriori approfondimenti per quanto riguarda l’affidabilità delle stime della necromassa nei cedui mediterranei invecchiati.
Sia le caratteristiche stazionali che quelle del popolamento esaminato, così come descritte nei paragrafi precedenti, rientrano in quelle medie dei boschi circostanti e non sembrano possedere alcuna particolarità tale da rendere oltremodo complessa la raccolta dei dati. In ogni caso, pendenza e morfologia sono caratteristiche che non dovrebbero poter influenzare in maniera significativa metodologie standardizzate di stima.
Anche la non omogenea distribuzione, sia quantitativa che qualitativa, della necromassa al suolo, così come riscontrata nell’area campione, rappresenta in genere la regola, soprattutto nei boschi invecchiati o comunque non regolarmente assoggettati a trattamenti colturali.
Il metodo xilometrico, utilizzato nell’indagine come “sistema di confronto”, non è sicuramente una metodologia proponibile su larga scala per questo tipo di indagini. Anche a prescindere dal peso e dall’ingombro dell’attrezzatura, richiede la disponibilità di notevoli quantità di acqua e di tempo (raccolta e separazione delle diverse tipologie di necromassa, trasporto al luogo di misura, immersione e imbibizione del legno, pulizia periodica delle vasche, ecc.). In compenso fornendo ragionevoli motivi di affidabilità ed errori sicuramente molto contenuti, rappresenta un ottimo testimone cui far riferimento.
La ricerca ha invece messo in evidenza, pur nei limiti dell’unico caso di studio affrontato (area investigata di 2500 m2), la divergenza, ancorchè risultata non significativa, delle stime derivanti dall’utilizzazione delle metodologie LIS e FAS, soprattutto in presenza di una elevata quantità di necromassa di piccole dimensioni, come in parte già rilevato anche in altri lavori ([42]).
La prima presenta una discreta semplicità e rapidità di esecuzione, soprattutto quando la soglia minima di rilievo è più elevata, non richiede lo spostamento della necromassa misurata e può quindi essere ripetuta nel tempo sulla stessa superficie.
La metodologia FAS è leggermente più laboriosa e può richiedere lo spostamento dei pezzi di legno da misurare.
Il metodo delle “pesate” porta con sé alcuni degli inconvenienti della metodologia xilometrica. Anche se l’attrezzatura necessaria può al limite essere costituita anche solo da una stadera di buona portata, il materiale selezionato deve essere raccolto in ceste per la pesatura o, se di notevole lunghezza, depezzato o legato in fasci.
Occorre poi una certa accuratezza nella scelta del campione da portare in laboratorio per la determinazione dei rapporti peso/volume, anche se poi i risultati hanno dimostrato una discreta omogeneità nelle caratteristiche dei campioni analizzati.
Nonostante la laboriosità, il metodo ha però fornito degli ottimi risultati, con stime che non differiscono dai dati ottenuti con lo xilometro. Per questo motivo il metodo delle “pesate” potrebbe essere utilizzato come controllo, quando ritenuto necessario, nell’ambito dei rilievi che utilizzano le metodologie LIS e FAS.
Ringraziamenti
Questo lavoro è stato reso possibile grazie alla fattiva e costante partecipazione dei tecnici del GESAAF, Fabio Bandini, Alberto Pierguidi e Stefano Teri. Un ringraziamento va anche al personale dell’Ufficio Territoriale per la Biodiversità di Follonica (GR) per il supporto logistico, e in particolare ai funzionari Dott. Stefano Vagniluca e Giovanni Quilghini.
References
CrossRef | Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar