Climate change and mountain Grouse: recent evidences from alpine habitats
Forest@ - Journal of Silviculture and Forest Ecology, Volume 10, Pages 26-32 (2013)
doi: https://doi.org/10.3832/efor0938-010
Published: Feb 04, 2013 - Copyright © 2013 SISEF
Technical Reports
Abstract
Current climate change, referring as well to the observed rain and temperature patterns as to the increased frequency and intensity of extreme weather conditions, has a deep influence on biotic communities and, in particular, on mountain Grouse. These species show great adaptation to coldness, are highly sedentary and have quite “strict” ecological requirements, when it deals with habitat selection. Moreover, their alpine ranges are dangerously marginal to the main distribution areas, which increases the risk of dramatic changes in occurrence, demography and ecology. However, not all the species will predictably be exposed in the same way to the menace of climate change over the next 50-100 years. This article gives a brief review of the main data acquired in the alpine environment in this matter. It also underlines the utmost need to proceed with research and monitoring activities, in order to effectively adapt and manage conservation strategies on mid-long terms.
Keywords
Il cambiamento climatico e le specie animali
Il sistema climatico globale sta rapidamente cambiando, sia in termini di patterns osservati di temperature e precipitazioni, sia per quanto riguarda l’aumento della frequenza e dell’intensità di fenomeni meteorologici estremi. I modelli climatici globali oggi disponibili prevedono inoltre un ulteriore riscaldamento significativo a livello planetario, variabile tra 1.1 e 6.4 °C entro il 2100 ([6]).
Negli anni più recenti si sono potuti documentare sotto il profilo scientifico gli impatti del cambiamento climatico (CC) su differenti aspetti della vita degli organismi animali ([22]): in particolare, per quanto concerne gli uccelli, si è potuto dimostrare che nella dimensione spaziale il CC ha indotto spostamenti dei range distributivi specifici verso i poli ed a quote superiori (ad es., [14]). Il CC in atto sta anche causando l’estinzione di alcune specie appartenenti a taxa differenti ([10]) e continuerà a provocarne altre nel prossimo futuro ([17], [18]).
È d’altro canto ormai ritenuto patrimonio acquisito dalla comunità scientifica internazionale il fatto che gli organismi animali adattati al freddo sono - e saranno - quelli maggiormente condizionati dal CC in atto. A puro titolo di esempio, quello che potremo definire il “portabandiera” degli ecosistemi artici, ossia l’orso polare (Ursus maritimus - Fig. 1), soffre oggi sia a causa della perdita diretta di habitat che per la ridotta accessibilità alle sue prede ([13], [21], [5]).
Fig. 1 - Un subadulto di orso polare sembra interrogarsi su come riuscire a superare una superficie marina prima ghiacciata ed ora in corso di scioglimento. L’innalzamento delle temperature comporta un’esposizione sempre maggiore degli orsi polari a superfici ghiacciate solo marginali e ad aree di mare aperto laddove, storicamente, vi erano habitat di ghiaccio marino stabile (Foto: Daniel J. Cox - ⇒ http://www.naturalexposures.com).

In realtà, in termini più generali, sono state proposte nel tempo una grande varietà di tipologie di risposta degli organismi animali al CC, alcune delle quali piuttosto complesse ([20]). Negli uccelli, il CC può così influenzare direttamente i tassi demografici ([16], [19]), ma più comunemente - come accennato - è associato a spostamenti degli areali distributivi ([12], [22]) o a modificazioni nella fenologia riproduttiva, che possono condurre al fenomeno noto come “mismatch” (=discordanza, discrepanza). In questo caso i patterns di CC a scala regionale differiscono tra la stagione di deposizione delle uova e quella della loro schiusa, determinando una cosiddetta “asimmetria (climatica) stagionale”, con la conseguenza che la sincronizzazione tra le esigenze trofiche della prole e la massima disponibilità di nutrimento può venire irrimediabilmente compromessa ([7]). Gli impatti per le specie sono inoltre tanto più gravi quanto più la selezione evolutiva, che conduce nel lungo periodo a modificare le date di inizio della riproduzione nella maniera più opportuna, ha possibilità di azione con un tempo di latenza necessariamente prolungato ([11]). In altre parole certe specie potranno sì adattarsi a questo cambiamento, alla sola condizione però che la selezione genetica di individui adattati alle nuove condizioni possa intervenire in modo sufficientemente rapido.
Valutazioni dell’impatto del cambiamento climatico sui Galliformi di montagna nelle Alpi
I Galliformi di montagna sono specie marcatamente sedentarie e la loro capacità di dispersione non eccede qualche decina di km: le loro esigenze vitali inoltre, dal punto di vista della selezione degli habitat sia riproduttivi che di svernamento, sono relativamente “strette” ([11]).
Sulle Alpi, inoltre, la situazione geografica marginale di queste specie le rende, nell’ipotesi di proseguimento del CC in atto, particolarmente esposte a modificazioni importanti in termini distributivi, demografici ed ecologici, in maniera decisamente più marcata di quanto ci si potrebbe aspettare nel cuore della loro area di distribuzione.
È da dire che oggi i previsti spostamenti degli areali distributivi specifici sono largamente dominati dai risultati di modelli di contesto bioclimatico. Sebbene questi modelli possano rilevare le dimensioni generali degli effetti del CC sulla biodiversità, sulla distribuzione degli areali e sui rischi relativi di estinzione delle specie, essi soffrono di diverse limitazioni, la principale delle quali - oltre alle numerose semplificazioni necessariamente impostate (ad es., la generale mancanza di valutazione del livello delle comunità piuttosto che delle relazioni inter- ed intra-specifiche) - è costituita dal non collegamento dell’impatto del CC sulle specie bersaglio rispetto all’effetto delle altre modificazioni ambientali ([2]). Queste includono processi che influenzano la vocazionalità degli habitat e l’evoluzione della vegetazione e le mettono in relazione con i cambiamenti nell’uso del territorio da parte dell’uomo risultante da fattori socio-economici, a loro volta spesso dipendenti dal clima ([1]). L’impatto combinato di questi cambiamenti può generare diminuzione, aumento e/o frammentazione degli habitat di specie differenti ([8]).
La composizione e la struttura della vegetazione determinano la vocazionalità dell’habitat di molti animali terrestri ed influenzano fortemente l’uso del territorio da parte dell’uomo. Sotto questo profilo solo modelli dinamici degli spostamenti della distribuzione potenziale delle specie, in grado di incorporare sia i processi in atto negli habitat che le caratteristiche demografiche ed il potenziale di dispersione, come anche le risposte antropiche nell’uso del territorio, possono realizzare più realistiche previsioni per gli scenari futuri.
Sulle Alpi, è prevedibile come i principali impatti negativi del CC interesseranno gli specialisti di habitat degli ecosistemi subalpini ed alpini. Qui, cinque specie di Galliformi di montagna (quattro Tetraonidi: Gallo cedrone, Tetrao urogallus; Francolino di monte, Bonasa bonasia; Fagiano di monte, Tetrao tetrix; Pernice bianca, Lagopus muta; ed un Fasianide: Coturnice, Alectoris graeca) sono distribuite secondo un gradiente ecologico che varia dai paesaggi culturali semi-aperti di mezza montagna alla tundra alto-alpina. I Tetraonidi sono organismi-modello assolutamente appropriati per studiare gli impatti diretti ed indiretti del CC sulla biodiversità alpina. Tutte le quattro specie di Tetraonidi alpini mostrano caratteristiche morfologiche di organismi adattati al freddo e si trovano al limite sud del loro areale distributivo europeo ovvero mondiale, ma è prevedibile che non tutte saranno ugualmente esposte alla minaccia del CC nei prossimi 50-100 anni, proprio in considerazione del fatto che l’impatto relativo del CC e del cambiamento dell’uso del territorio (e le loro interazioni) condizionerà in maniera abbastanza differenziata la loro vitalità. Certo è che in ogni caso si tratta di indicatori molto sensibili delle conseguenze del CC degli ecosistemi montani ed alpini.
Oggi, Gallo cedrone e Francolino di monte, i due Tetraonidi alpini forestali, sono classificati con rischi di estinzione più elevati rispetto al Fagiano di monte ed alla Pernice bianca se teniamo in considerazione le “liste rosse” dei paesi europei che hanno territorio nelle Alpi: Gallo cedrone e Francolino di monte sono infatti classificati come specie minacciate in sei su sette “liste rosse” nazionali, il Fagiano di monte in quattro e la Pernice bianca in tre ([2]). In sintesi, le minacce complessive per le specie di Tetraonidi sembrano decrescere negli habitat alpini passando dalle quote inferiori a quelle maggiori. Evidentemente, negli ultimi 150 anni questa caratteristica è stata determinata da differenze nel tipo e nell’intensità dell’utilizzo antropico del territorio: cambiamenti indotti dall’uomo nella composizione e nella struttura della vegetazione hanno così influenzato la vocazionalità degli habitat delle foreste montane più di quanto non sia accaduto per i pascoli e la tundra alpini, habitat rispettivi di Fagiano di monte e Pernice bianca.
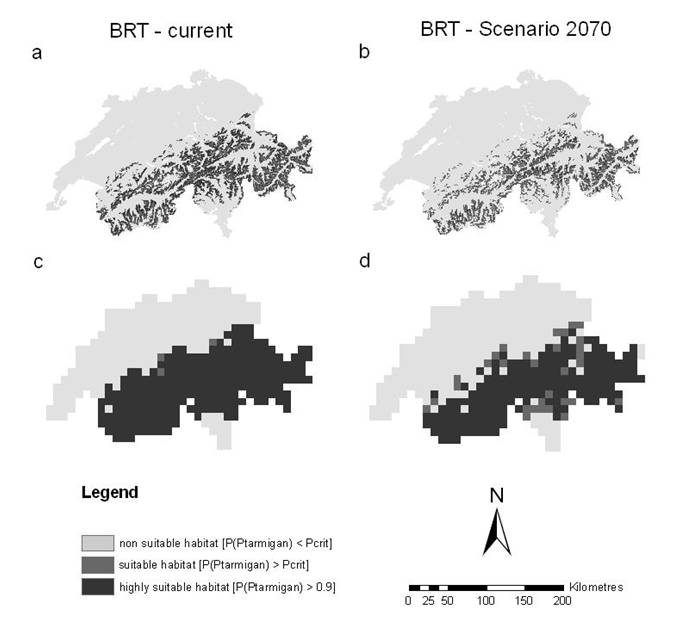
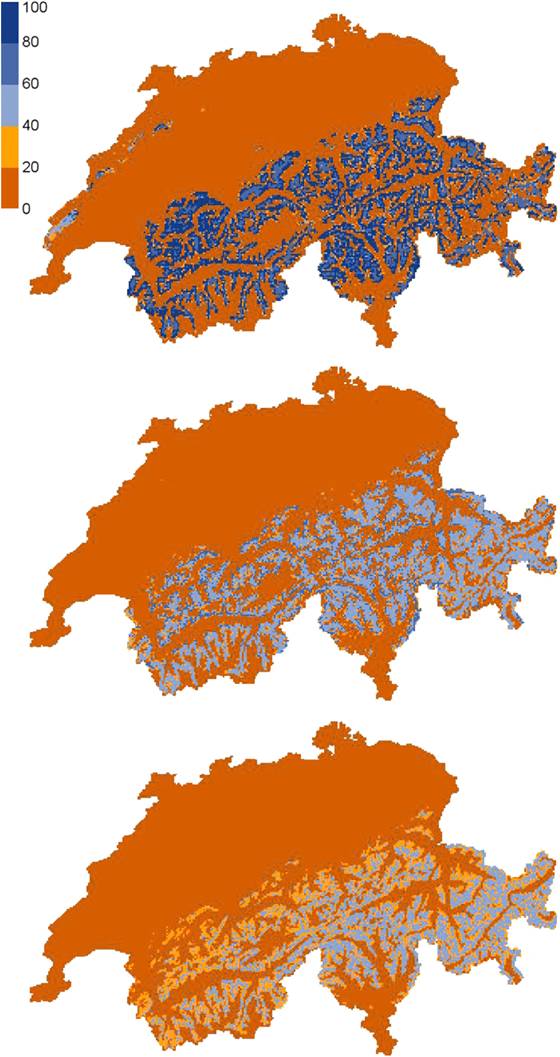
Al momento, vi sono alcune evidenze che probabilmente il CC ribalterà le differenze nell’entità delle minacce complessive tra le specie di Tetraonidi tipiche di foreste montane e dei pascoli/tundra alpini. Il previsto incremento di eventi meteorologici estremi, con conseguenti tempeste di vento e siccità estive, direttamente o indirettamente influenzerà positivamente la vocazionalità degli habitat di Gallo cedrone e Francolino di monte. D’altro canto, le dinamiche più recenti registrate nell’espansione forestale soprattutto nella fascia più esterna delle Alpi (causate dall’abbandono dell’utilizzo del suolo da parte dell’uomo per i tradizionali fini agro-silvo-pastorali) avranno un impatto negativo sulle condizioni dell’habitat del Fagiano di monte. Probabilmente quest’ultima specie in aree montane periferiche, poste a quote inferiori e scarsamente strutturate, perderà una notevole quota del proprio habitat nei prossimi decenni, indipendentemente dagli effetti diretti del CC. La Pernice bianca, al contrario, è meno esposta agli effetti di cambiamenti di uso del suolo, mentre - localmente - impatti negativi potranno registrarsi per attività ricreative più frequenti, sebbene questi raramente possano avere conseguenze su larga scala. D’altro canto, la specialista degli habitat di tundra alpina è più esposta rispetto agli altri Tetraonidi a limitazioni fisiologiche provocate dal CC. I modelli di contesto climatico sono concordi nel predire una drastica riduzione dell’areale distributivo di questa specie nel prossimo secolo (ad esempio, Fig. 2- [15]): ma occorre valutare come le possibilità di persistenza della Pernice bianca in condizioni climatiche meno favorevoli sono al momento del tutto sconosciute. È assolutamente difficile, in altri termini, prevedere gli effetti di un innalzamento delle temperature sulla eco-fisiologia della specie, tema di ricerca che meriterebbe davvero di essere approfondito ([11]).
Fig. 2 - Habitat potenziale della Pernice bianca in Svizzera nelle condizioni attuali (a, c) e alle condizioni degli scenari intermedi al 2070 (b, d) secondo il modello BRT (boosted regression trees) alle due scale spaziali.(a, b): dimensione della cella = 1 km2; (c, d): dimensione della cella = 100 km2 (fonte: [15]).
In letteratura, come sopraccennato, è stato documentato anche un effetto della asimmetria stagionale nel CC regionale in Finlandia che influenza negativamente le condizioni meteorologiche per la riproduzione del Fagiano di monte (mismatch hypothesis - [7]). Temperature più calde in primavera hanno infatti determinato, a partire dal 1987, una riproduzione ed una schiusa anticipata in diverse popolazioni studiate. Dal momento però che le condizioni meteorologiche preferibili per lo sviluppo delle covate appena schiuse (Fig. 3) non si sono simultaneamente anticipate, questo CC ha determinato una cosiddetta asimmetria stagionale tra primavera ed estate, che ha condizionato negativamente il successo riproduttivo, fino a portare il tasso intrinseco di crescita delle popolazioni studiate a diminuire, tanto che le stesse si sono ridotte significativamente negli ultimi 4-5 decenni. Un simile pattern d’altro canto non è stato ancora dimostrato per quanto riguarda i Tetraonidi alpini, sebbene qualche indicazione sia disponibile per il Gallo cedrone sui Pirenei ([11] - Fig. 4). È in effetti piuttosto complesso valutare come questa asimmetria stagionale tra condizioni meteorologiche medie primaverili ed estive in scenari di CC possa influenzare le condizioni delle femmine riproduttrici piuttosto che l’abbondanza di insetti durante l’allevamento dei pulli o la sopravvivenza dei pulli per se. In relazione a questo aspetto merita anche ricordare come nella situazione alpina Zbinden & Salvioni ([23]) hanno dimostrato come, inversamente alla situazione del contesto scandinavo, l’alimentazione primaverile delle femmine non costituisce un fattore limitante della riproduzione annuale, perché la fase di riscaldamento primaverile condizionante la crescita delle piante dura circa due mesi: all’inverso, le estati presentano una grande variabilità, in termini di freschezza e di umidità, ed inducono delle variazioni interannuali importanti nella mortalità dei pulcini. Di conseguenza, la demografia della specie sarà particolarmente condizionata in ambiente sub-artico dall’evoluzione climatica primaverile e, in zona alpina, da quella estiva. In termini di azioni di conservazione quindi, gli sforzi dovranno essere riservati in ambiente alpino al miglioramento della qualità degli habitat di allevamento delle covate, per limitare i rischi di compromissione del tasso di sopravvivenza dei pulli.
Fig. 3 - Relazione tra temperatura media massima dei 10 giorni post-schiusa e indice di output riproduttivo, r=0.63, P<0.001, N=40 (fonte: [7]).
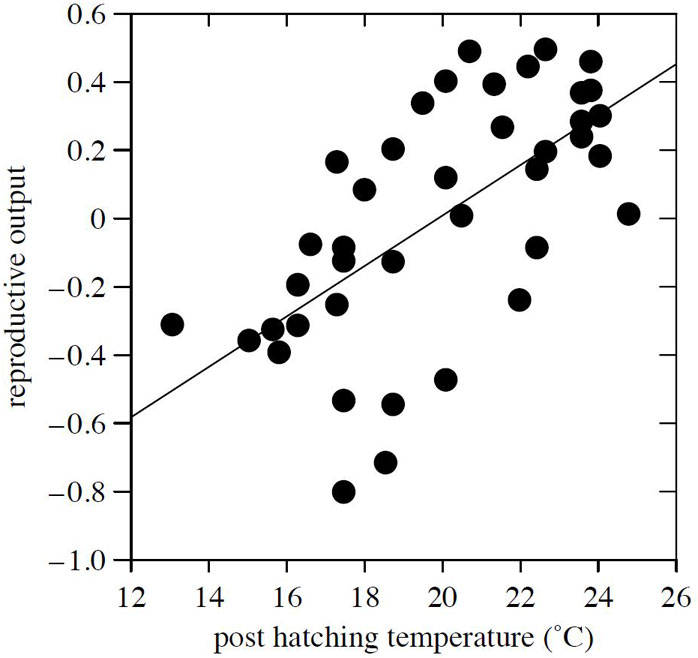
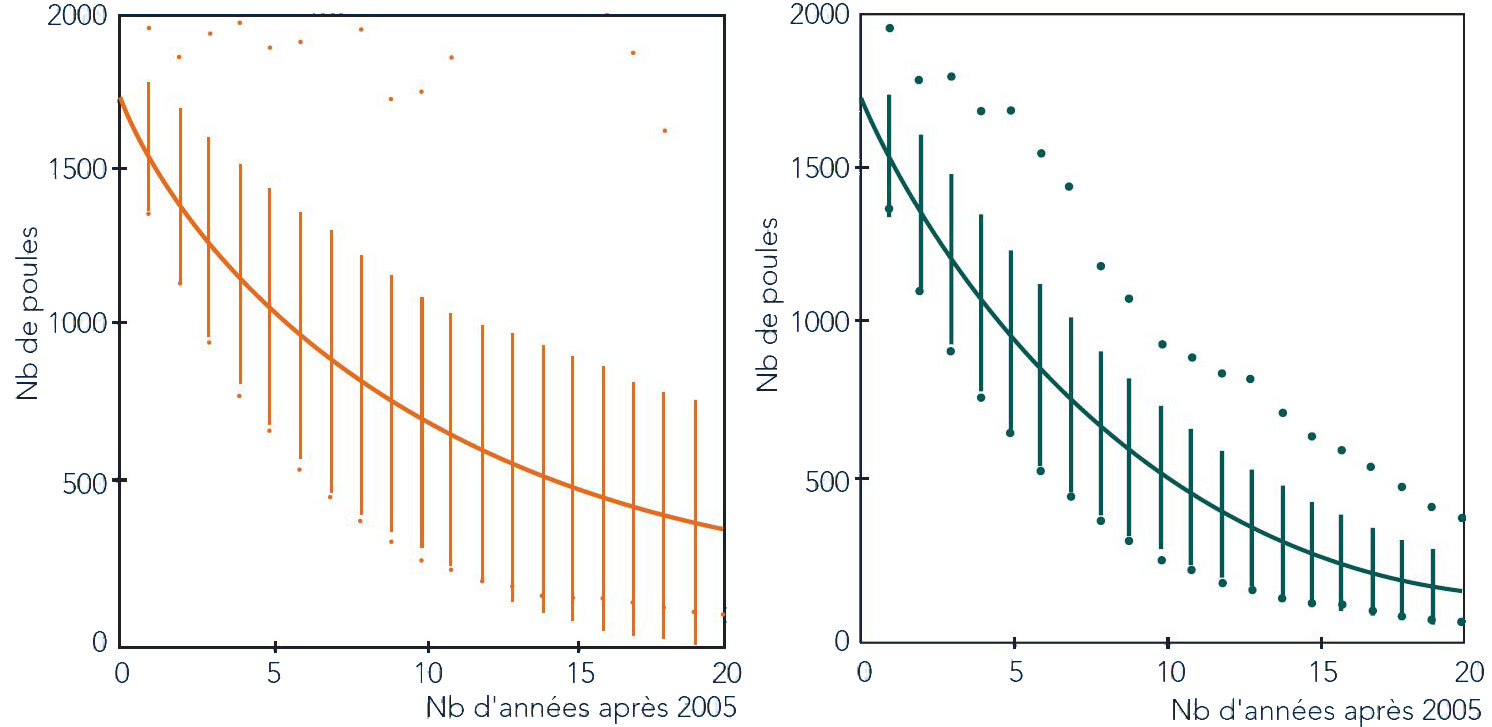
Fig. 4 - Evoluzione della consistenza del segmento femminile della popolazione di Gallo cedrone nei Pirenei per i 20 anni successivi al 2005 - sulla base di 1000 simulazioni - nello scenario a clima costante (a sinistra) e di proseguimento tendenziale all’aumento delle precipitazioni primaverili (a destra). Le barre verticali ricomprendono l’ampiezza della totalità delle simulazioni, mentre la curva rappresenta il valore medio dell’insieme delle simulazioni stesse (fonte: [11]).
Bollmann ([2]) rileva pure come un ulteriore fattore difficile da prevedere sia costituito dal tasso di utilizzazione delle foreste alpine, causato da una richiesta energetica in progressivo aumento. In senso generale è da prevedere un effetto positivo di tassi di utilizzazione più elevati, perché mediamente nelle Alpi in molte regioni la provvigione legnosa ha raggiunto un massimo storico alla fine del secolo scorso, deteriorando in effetti la vocazionalità ambientale di specialisti di foreste aperte come Francolino di monte e Gallo cedrone. È d’altro canto evidente che se si vuole generare una sinergia positiva tra l’utilizzazione delle foreste e la vocazionalità degli habitat forestali nel prossimo futuro, molti risultati ecologici della ricerca sui Tetraonidi forestali contemporanea debbono essere presi in considerazione ed attentamente trasfusi in sede applicativa ([3]).
Bollmann ([2]) afferma in conclusione che in ogni caso il cambiamento di uso del territorio da parte dell’uomo avrà una maggiore influenza sulla distribuzione e l’abbondanza dei Tetraonidi alpini di quanto non avrà il CC, almeno nella zona montana e subalpina (Tab. 1).
Tab. 1 - Impatto previsto del cambiamento climatico (CC) e di uso del suolo (CUS) sulle quattro specie di Tetraonidi alpine ed impatto integrale previsto sulla vitalità delle rispettive popolazioni nell’arco dei prossimi 50-100 anni (fonte: [2], modificata). (↑): aumento; (↓): diminuzione; (---): forte impatto negativo; (--): impatto negativo considerevole; (-): basso impatto negativo; (0): nessun impatto; (+): basso impatto positivo; (++): impatto positivo considerevole; (+++): forte impatto positivo; (-/+): impatto dipendente dal pattern stagionale.
| CC/CUS causa |
Caratteristiche ambientali | Francolino di monte | Gallo cedrone | Fagiano di monte | Pernice bianca |
|---|---|---|---|---|---|
| ↑ | Temperature e precipitazioni annue | -/+ | -/+ | -/+ | - |
| ↑ | Disturbo naturale (ad es. tempeste di vento, siccità, calamità da insetti) | +++ | ++ | + | + |
| ↑ | Superficie forestale | + | ++ | - | 0 |
| ↓ | Superficie di pascoli alpini | 0 | 0 | -- | - |
| ↑ | Disturbo antropogeno | (-) | - | - | - |
| Vitalità integrale prevista | ++ | + | -- | - |
Conclusioni
Le esigenze di ricerca sul tema dell’influenza del CC sulla biodiversità sono certo davvero pressanti: e a tal riguardo si fa nuovamente rilevare come il CC agisce in realtà sulle comunità di specie, le cui relazioni interspecifiche possono risultare modificate anche profondamente. Dal momento che risulta ancora piuttosto difficile disporre di scenari climatici completamente verosimili, tutte le conseguenze di queste modificazioni sono però ancora lontane dall’essere perfettamente comprese ([11]). D’altro canto, le stesse non possono che essere indagate attraverso simulazioni, che necessitano di una sinergia profonda e consapevole tra biologi, meteorologi ed ecologi modellisti (Fig. 5 - [24]). Inoltre, un ruolo importante di modulazione potrà essere assunto dalle modificazioni nelle interazioni biotiche, dai processi di espansione/contrazione delle componenti vegetali degli ecosistemi ([4]) e dalla perdita della connettività degli habitat. La prosecuzione delle ricerche e del monitoraggio sui Galliformi di montagna è in conclusione assolutamente necessaria per riuscire a proporre adattamenti nelle strategie di conservazione di queste specie che risultino efficaci nel medio-lungo periodo ([9]), tenendo conto delle modificazioni attese nelle condizioni ambientali in senso lato, e più specificamente, appunto, di quelle climatiche.
Fig. 5 - Consenso sulla presenza del Fagiano di monte in Svizzera per gli anni 2001 (in alto), 2050 (al centro) e 2100 (in basso), calcolato come la frazione di tutte le simulazioni (N=1212) che hanno previsto che la specie sia presente. Da notare che il consenso percentuale pari a 0 sulla presenza del Fagiano di monte equivale al consenso pari al 100% sull’assenza della specie (fonte: [24]).
Ringraziamenti
Questo lavoro è tratto da una relazione presentata alla 59a Assemblea AGJSO (Artbeitsgemeinschaft der Jagdverbände des Südostalpenraumes - Comunità di lavoro delle organizzazioni venatorie dell’area alpina sud-orientale), tenutasi a Feltre (BL) il 20-21 ottobre 2011. La versione iniziale della presente nota è stata rivista in seguito ai preziosi suggerimenti di Luca Rotelli: le sue osservazioni hanno consentito di rendere il lavoro più preciso e corretto. Desidero ringraziare inoltre Giorgio Carmignola per l’interessante confronto sulla situazione altoatesina, e infine Roberta Covi, che ha gentilmente curato la traduzione del riassunto.
References
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar