Evidences from long-term monitoring of Italian forests. Tree radial growth as response index to disturbances and its relations with the stand structure
Forest@ - Journal of Silviculture and Forest Ecology, Volume 10, Pages 68-78 (2013)
doi: https://doi.org/10.3832/efor1010-010
Published: Sep 02, 2013 - Copyright © 2013 SISEF
Research Articles
Guest Editors: LIFE FUTMON Project
« Further Development and Implementation of an EU-level Forest Monitoring System »
Collection/Special Issue: Luca Salvati (CRA-RPS, Roma)
Abstract
The paper deals with the work undertaken since 1995 within the national level II network framed into the ICP-Forests ICP-IM programme. A synthesis of results from tree growth monitoring and relationships with stand structure and related parameters are reported. Current changes in the growth medium, i.e., physics and chemistry of atmosphere and soil (increase of average air temperature, rainfall shortage and drought, CO2 enrichment, ozone level, nitrogen fertilization, sulphate deposition) drive today the soil-tree-atmosphere relationships. The overall result of these concurrent and counteracting factors is recorded over each growing seasons by radial stem growth, providing a sensitive response index to disturbances. A few occurrences of disturbances to growth at regional and at case-study level, likely due to climate deviations, are discussed. Seasonal fluctuations and anomalous or extreme events are the major occurrences over the last decade. The heat wave 2003 is the main case occurred over a large part of Europe. Growth rate 2000-2004 showed reductions up to 50% in comparison to 1997-2000 on plots located within the southern continental border of the heat wave, in particular at low elevations and for pre-determined early growth species (beech and oaks). Over the following time-window 2005-2009, a significant growth decrease was vice versa detected within the coniferous spruce forests located at medium-high elevation in the Alps, where repeated seasonal anomalies both in air temperature and rainfall were recorded over the same time-span. The heavy effect of climate disturbance at a local scale was examined where two oak species with different auto-ecological features grow in sympatry. The rationale and awaited goals from protocols’ updating and the intensive surveys applied to core-areas in 2009-2010 under LIFE+FutMon are reported. Perspectives at short to medium term of monitoring programme at national and European level are also debated.
Keywords
Forest Ecosystem, Intensive Monitoring, Tree Growth, Stand Structure, Disturbance
Introduzione
Tutte le componenti biologiche di un sistema, ciascuna nel proprio habitat e ambito funzionale, interagiscono continuamente con l’ambiente di crescita. Qualsiasi variazione nello stato fisico-chimico di atmosfera e suolo produce una reazione dei meccanismi di controllo specifici e l’aggiustamento eventuale delle connessioni reciproche. Ciascuna popolazione genera retroazioni di resistenza, resilienza, adattamento entro i limiti delle proprie tolleranze auto-ecologiche. Entità, direzione, durata dei cambiamenti condizionano modi, tempi e capacità di risposta delle componenti biologiche, quindi anche degli alberi. Una rete di monitoraggio in continuo consente di esplorare la dinamica e la dimensione spazio-temporale del fenomeno dal livello locale a quello regionale fino a scala geografica attraverso contesti ambientali e specie diverse.
La rete nazionale di “monitoraggio intensivo degli ecosistemi forestali” di livello II, attiva dal 1995 nell’ambito del programma pan-europeo ICP-Forests ICP-IM ([1], [20], [21]), impostata secondo un disegno di tipo preferenziale, è formata da 31 casi di studio. I siti sono collocati entro superfici di vegetazione massale delle specie, da cui si assume che non pre-esistano particolari fattori limitanti. La risposta biologica attesa rimane così espressione prevalente del campo di variazione della fertilità naturale. La costruzione della rete su tipologie di bosco molto rappresentate e la ripetizione dei medesimi tipi attraverso il gradiente geografico di vegetazione, consentono di ricavare informazioni distribuite sul territorio.
Obiettivi della rete di monitoraggio intensivo sono la verifica della “salute e vitalità” (stato delle componenti biologiche) dei sistemi sotto osservazione e la determinazione di presenza, consistenza ed effetto di eventuali fattori di disturbo. Il protocollo di ricerca comprende i tre comparti, chimico, fisico e biologico su cui si articola il sistema. I primi due sono monitorati direttamente attraverso l’analisi dei fattori agenti secondo i rispettivi livelli, carichi o eventi critici: chimica delle deposizioni e delle soluzioni circolanti nel suolo, chimica fogliare e della lettiera, livello di ozono, andamento dei parametri del clima locale e registrazione di eventi anomali o estremi. La comunità biologica, che produce le reazioni agli input di tipo fisico-chimico, si avvale viceversa di indicatori e indici di risposta individuati in componenti diverse di una stessa popolazione (ad es., stato delle chiome e accrescimento radiale degli alberi) e di popolazioni diverse (ad es., flora vascolare e licheni epifiti).
Obiettivo di questo contributo è la sintesi dei risultati del monitoraggio dell’accrescimento legnoso radiale del fusto nel periodo 1996-2010. Si riportano le principali evidenze di disturbo a scala regionale e di caso di studio, si descrivono le azioni complementari svolte nel progetto LIFE+ FutMon (2009-10) e le prospettive del protocollo di monitoraggio.
Accrescimento e struttura del bosco
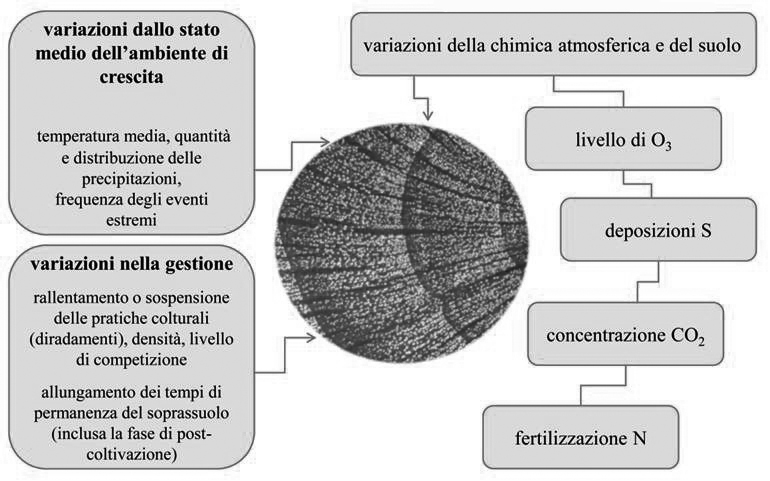
L’accrescimento è un fenomeno quantitativo misurabile a scala individuale, di strato, di popolazione. È il descrittore della fertilità del sito, della corrispondenza auto-ecologica ai caratteri della stazione e dell’abilità competitiva di ciascuna specie nell’ambiente di crescita relativo. L’accrescimento è condizionato da qualsiasi modifica dello stato medio, dalla chimica e fisica dell’atmosfera e del suolo, da mutamenti nella gestione colturale ed è indice di risposta sensibile della “salute e vitalità” del sistema (Fig. 1). Le modificazioni possono avere un effetto concorrente (ad es., innalzamento delle temperatura media dell’aria + arricchimento della CO2 atmosferica + sufficiente disponibilità idrica + fertilizzazione azotata → sinergia positiva); o opposto (ad es., carenza di pioggia e temperature elevate nella stagione di crescita → stress idrico). L’effetto di un qualsiasi fattore non è univoco ma contestuale agli altri presenti e all’ambiente di occorrenza del fenomeno. L’aumento delle temperature medie può risultare in un allungamento positivo della stagione vegetativa alle alte latitudini o in siti di quota dove non esistono carenze idriche, mentre diventa un fattore limitante in aree mediterranee dove una maggiore evapotraspirazione concorre ad accentuare una normale carenza estiva di acqua nel suolo.
Fig. 1 - Fattori influenti sull’accrescimento radiale (da: [3]).
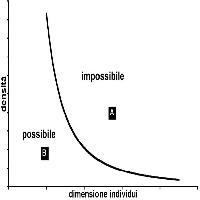
Obiettivo del monitoraggio dell’accrescimento arboreo è ottenere un “segnale” quanto più possibile filtrato dalle interferenze della struttura dimensionale e sociale. Qualsiasi popolazione arborea, naturale o modificata dalla selvicoltura applicata, è infatti strutturata in sotto-insiemi originati dal diverso risultato sociale di soggetti inizialmente simili. Questi vanno ad occupare in modo variabile lo spazio compreso tra il livello di esplorazione radicale e il piano delle chiome dominanti. La struttura arborea è la risultante fisionomica di questo processo. In popolazioni a ciclo di vita lungo quali quelle arboree, che prevedono l’aumento delle dimensioni unitarie con l’età in uno spazio definito che si sviluppa in senso verticale, la riduzione della densità iniziale è il postulato della vitalità del sistema. Il driver naturale di questo processo è la competizione per lo spazio e le risorse che realizza la progressiva selezione di alcuni individui, la mortalità di altri, la differenziazione dimensionale e sociale ([7]).
Accrescimento e mortalità sono quindi i feedback opposti che consentono lo sviluppo del bosco e determinano la formazione della struttura. L’auto-ecologia delle specie componenti può creare e mantenere complessità strutturali diverse. I sistemi colturali applicati semplificano di norma la struttura in modo evidente per finalità di produzione legnosa.
L’appartenenza dell’albero a una classe dimensionale o a uno strato verticale significa capacità di fotosintesi (massa fogliare e illuminazione relativa), di esplorazione del suolo (apparato radicale) e quindi di utilizzo delle risorse (energia, acqua nutrienti) asimmetrica verso altri soggetti di rango diverso. Questo si traduce in un accrescimento individuale differente.
La stratificazione dell’accrescimento per piani verticali o strati formati da individui simili per sviluppo, dimensioni e ambiente di crescita relativo è strumento efficace per ridurre l’interferenza strutturale. L’incremento nel piano dominante è per questa ragione utilizzato, insieme a quello di complesso, come “variabile di risposta” perché rappresentativo dei soggetti meno influenzati dal livello di competizione in atto. Gli individui dominanti sono inoltre l’interfaccia diretta con le deposizioni e lo stato dell’atmosfera (maggiore sviluppo e superficie di contatto delle chiome) e i maggiori utilizzatori delle risorse a livello radicale e quindi i soggetti più sensibili a variazioni nel mezzo di crescita ([8]).
Altri fattori specifici di ciascun soprassuolo che interferiscono con l’accrescimento potenziale sono età e densità unitaria.
L’etàè un determinante biologico comune a tutti gli esseri viventi; lo sviluppo degli alberi è approssimabile a una curva sigmoide con un primo tratto meno inclinato (età iniziale di affermazione e formazione), uno successivo più ripido che esprime la piena potenzialità di accrescimento (fase giovane e adulta), uno terminale di rallentamento e progressivo esaurimento incrementale (età matura e di senescenza). Le età osservate nei nostri boschi, per la quasi totalità coltivati e normalmente utilizzati nella fase adulta, sono ancora lontane dalla maturità biologica ed esprimono quindi, nella maggiore parte dei casi, un livello di accrescimento compreso entro il limite superiore della fase ottimale ([10]).
La riduzione del campo di applicazione della selvicoltura in atto da alcuni decenni nel nostro Paese per la marginalità economica di molti prodotti legnosi e lo sviluppo di funzioni complementari a quelle produttive, inizia a evidenziare casi di studio intermedi tra la fase adulta e quella di maturità. Ciò consente utili confronti nella prospettiva del consolidamento di un’area di post-coltivazione sempre maggiore che renderà molto più diffuse le classi di età mature ([9]).
La densità colma che determina un livello elevato di competizione interindividuale è viceversa una condizione già evidente e comune ai nostri soprassuoli. Se infatti i tagli di utilizzazione finale tendono in molte foreste ad essere posticipati indefinitamente, a questo si sovrappone l’effetto della sospensione dei tagli intercalari che avevano l’obiettivo di anticipare la mortalità naturale e di sostenere l’incremento legnoso del soprassuolo principale fino alla maturità prefissata (turno o diametro di recidibilità). Densità colme riducono i valori potenziali di accrescimento in tutta la popolazione e quindi anche nello strato dominante. Questo fattore può essere controllato confrontando le densità reali con il valore di riferimento SDI o indice di densità relativa.
A questo scopo sono confrontati range e valori medi di età, incremento corrente di volume e indice di densità (Fig. 2) rilevati sulla rete europea di livello II ([22]) con quelli dei siti italiani per le tre tipologie comuni: faggete, boschi di abete rosso, querceti.
Fig. 2 - Confronto del campo di variazione e del valore medio per i parametri: età del soprassuolo, incremento corrente di volume, indice di densità di popolamento (SDI) per tipologia sulla rete di livello II in Europa (ICP-EU) e in Italia (ICP-IT). Le età medie sono riferite all’anno 1995 sui valori riportati a 1.30 m ([22]). Le età attuali sono quindi maggiori di 15 anni più le annualità (10 in media) differenza tra l’età assoluta e quella alla quota di riferimento.
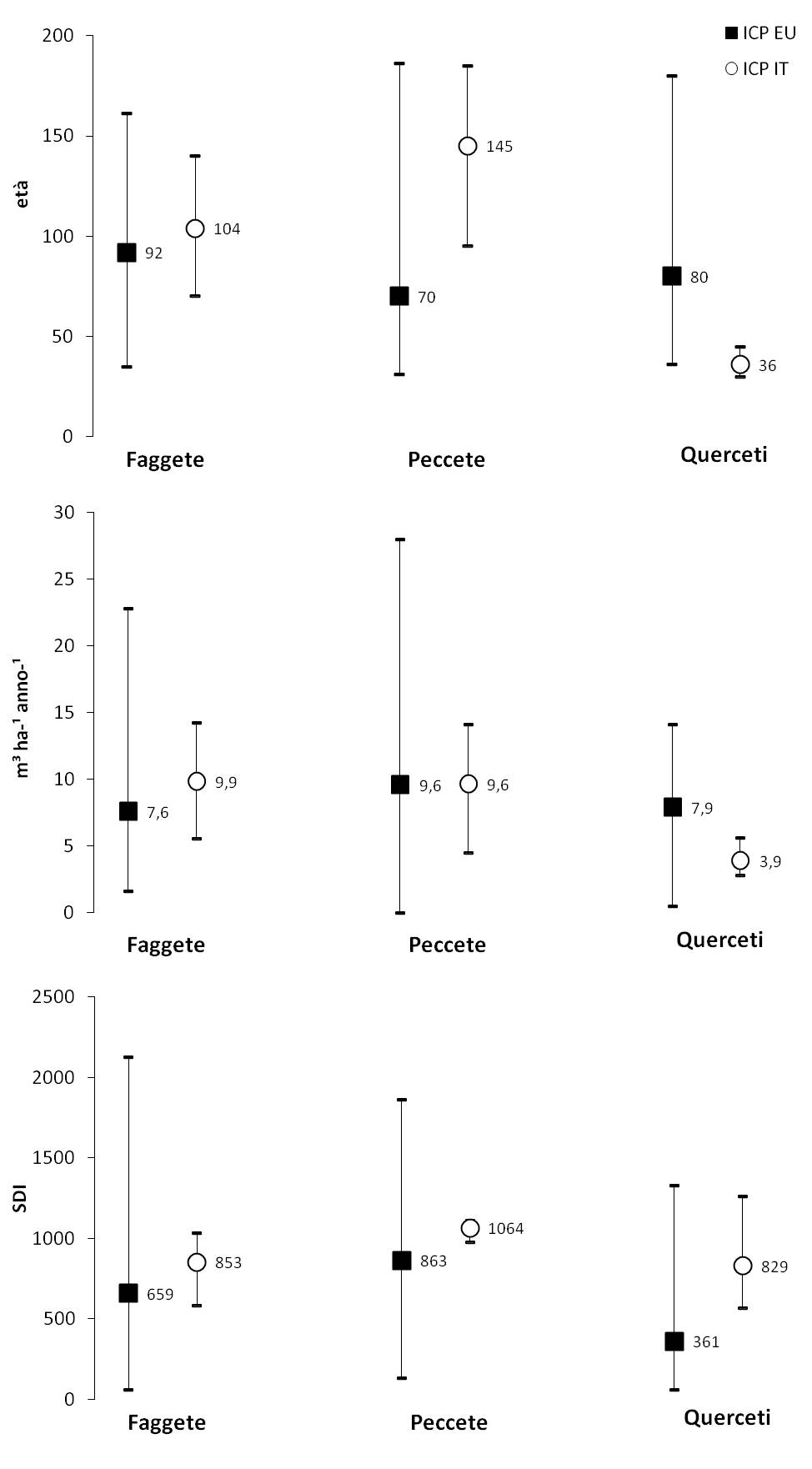
- Età: i casi di studio di faggete appenniniche e prealpina sono mediamente più adulti di quelli europei; le peccete alpine hanno un’età media circa doppia mentre i querceti caducifogli sono notevolmente più giovani. Il range europeo per i primi due tipi (faggete e peccete) è dovuto in prevalenza alle minori età rappresentate a livello continentale. Il caso dei querceti è opposto perché in Europa questa tipologia è generalmente presente in forma di fustaia, mentre in Italia (e altri Paesi mediterranei) lo è soprattutto come bosco ceduo oltre turno con soprassuoli mediamente molto più giovani. Il confronto conferma quanto detto sul rallentamento o sospensione in atto della gestione colturale nel nostro Paese e della tendenza quindi a prevalere dei soprassuoli adulti sulle classi giovani, come peraltro evidenziato dai dati dell’ultimo Inventario nazionale (INFC 2005).
- Incremento corrente di volume: la produttività legnosa è superiore di circa 2 m3 ha-1 anno-1 per i casi di studio di faggeta e valori eguali per quelli di abete rosso, nonostante le età medie superiori. La produttività dei querceti cedui è pari a circa la metà delle corrispondenti fustaie europee.
- Indice di densità di popolamento (SDI): i valori dei casi di studio italiani sono sempre mediamente superiori attraverso i tipi e riferibili alla applicazione ridotta o sospesa delle pratiche colturali, in particolare dei tagli intercalari. La differenza si amplia nei querceti, soprassuoli in post-coltivazione che crescono a livelli di elevata densità naturale per l’origine cedua. La rete europea comprende anche soprassuoli molto giovani e quindi naturalmente densi.
La misura della dimensione dei fusti e la sua ripetizione nel tempo sono la base per costruire la struttura dimensionale della popolazione, determinare l’accrescimento periodico (incremento), verificare la sua dinamica temporale.
Il collegamento tra accrescimento e struttura è descritto nelle Fig. 3 e Fig. 4. Sono rappresentati, per alcuni casi di studio di faggeta e di pecceta, la fisionomia del bosco, la curva di distribuzione delle frequenze dimensionali degli alberi e alcuni parametri di sintesi: età, massa in piedi, densità unitaria del soprassuolo. Sono riportati per piano (superiore, intermedio, inferiore) i valori di allocazione (G%) e quelli di incremento annuale (ΔG%) di area basimetrica, parametro utilizzato qui come proxy del volume arboreo.
Fig. 3 - Rappresentazione della struttura dimensionale (curva delle frequenze diametriche) e fisionomica, età, densità e massa in piedi del soprassuolo, parametri di allocazione e di incremento relativi ai piani individuati nei siti di faggeta distribuiti nel gradiente di latitudine osservato.

Fig. 4 - Rappresentazione della struttura dimensionale (curva delle frequenze diametriche) e fisionomica, età, densità e massa in piedi del soprassuolo, parametri di allocazione e di incremento relativi ai piani individuati nei siti di pecceta dell’arco alpino.
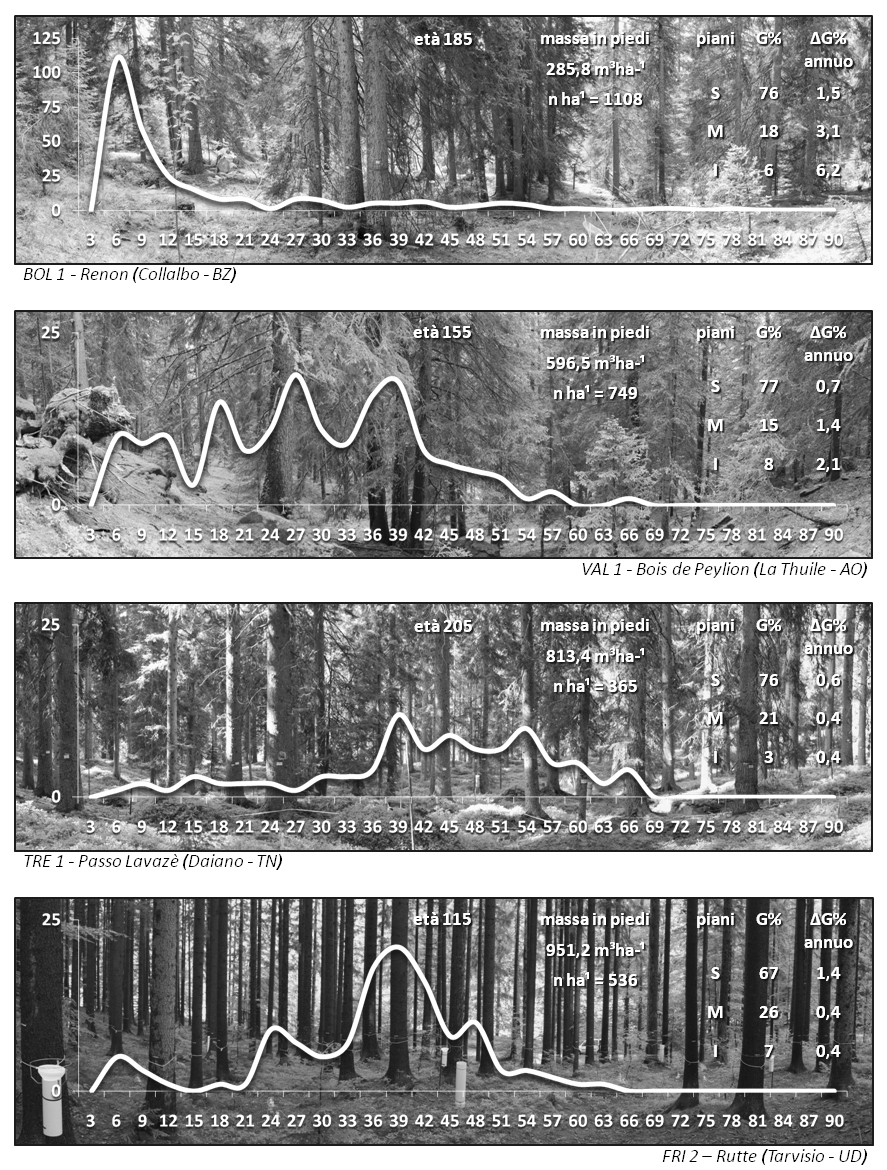
Si evidenzia il collegamento tra la fisionomia strutturale del bosco e l’andamento dimensionale (frequenze numeriche) che la rappresenta. Ciascuna curva, anche all’interno dello stesso tipo, presenta profili diversi che descrivono le aggregazioni spaziali, le densità e i dendrotipi originati. A queste strutture sono associate ripartizioni differenti dell’accrescimento legnoso allocato (G%) e della capacità corrente di incremento nei piani (ΔG%). La stratificazione consente così di monitorare sia la dinamica di complesso del soprassuolo che quella collegata a ciascun ambiente di crescita (piano), di verificare se esiste una diversa sensibilità di risposta in accrescimento per lo stesso o per tipi differenti di disturbo.
Le strutture coetanee di faggeta formate da dendrotipi e densità diverse e quelle disetanea (BOL1), irregolare (VAL1), coetanea su due classi di età (TRE1), coetanea (FRI2) di pecceta, presentano infatti caratteri differenziati di occupazione dello spazio verticale di crescita e di superficie esterna di contatto delle chiome con l’atmosfera (rugosità) e quindi una potenziale diversa sensibilità ai fenomeni di intercettazione e deposizione o a eventi anomali/estremi di tipo fisico-climatico.
Evidenze di disturbo dell’accrescimento nel periodo monitorato
Il livello geografico-regionale
Le evidenze più notevoli a oggi registrate a livello regionale e locale sono attribuibili al fattore climatico. A scala regionale, il primo fenomeno ha interessato le aree comprese entro il limite continentale meridionale dell’ondata di calore 2003 (Fig. 5). Nell’intervallo 1999-2004 sono riportati minori incrementi fino al 50% rispetto all’intervallo precedente su circa metà della rete.
Fig. 5 - Distribuzione dei livelli di riduzione dell’accrescimento attraverso la rete. Variazione nell’intervallo 2000-2004 rispetto al periodo 1997-1999 (da: [3]).
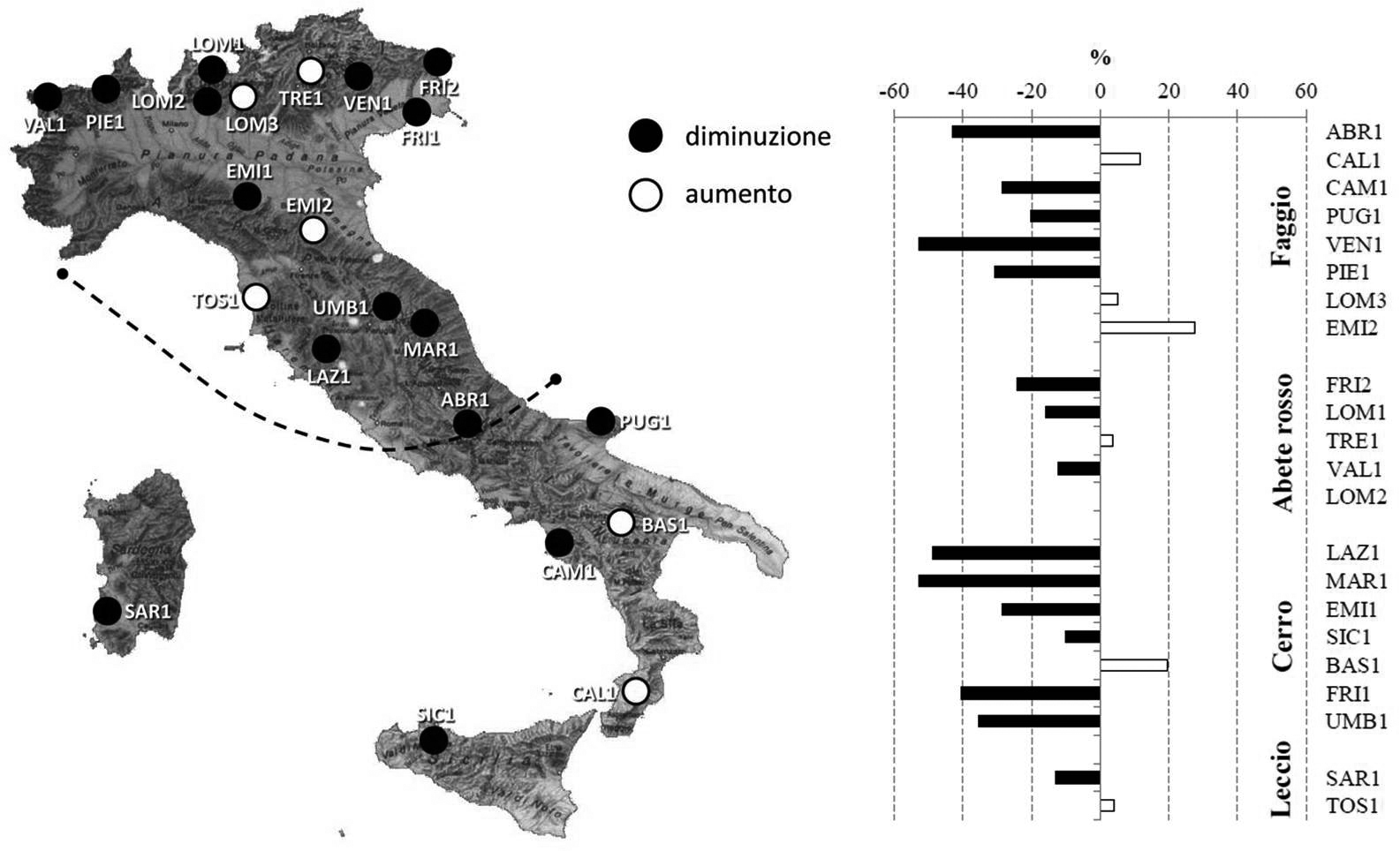
Carattere comune ai siti interessati sono la quota ridotta e le specie, querce decidue e faggio, ad accrescimento pre-determinato. Queste risentono del fattore di disturbo (elevate temperature associate a mancanza/carenza di precipitazione nel caso) anche nella stagione di crescita successiva (formazione della parte primaverile dell’anello legnoso). L’effetto si protrae quindi oltre l’anno di occorrenza del fenomeno e risulta così più marcato. Questo risultato è in accordo con quanto verificato per altre regioni europee e nell’ambito della stessa rete ICP da Jolly et al. ([17]) e da Dobbertin et al. ([6]) e in generale in Europa occidentale da Ciais et al. ([5]) e Leuzinger et al. ([18]).
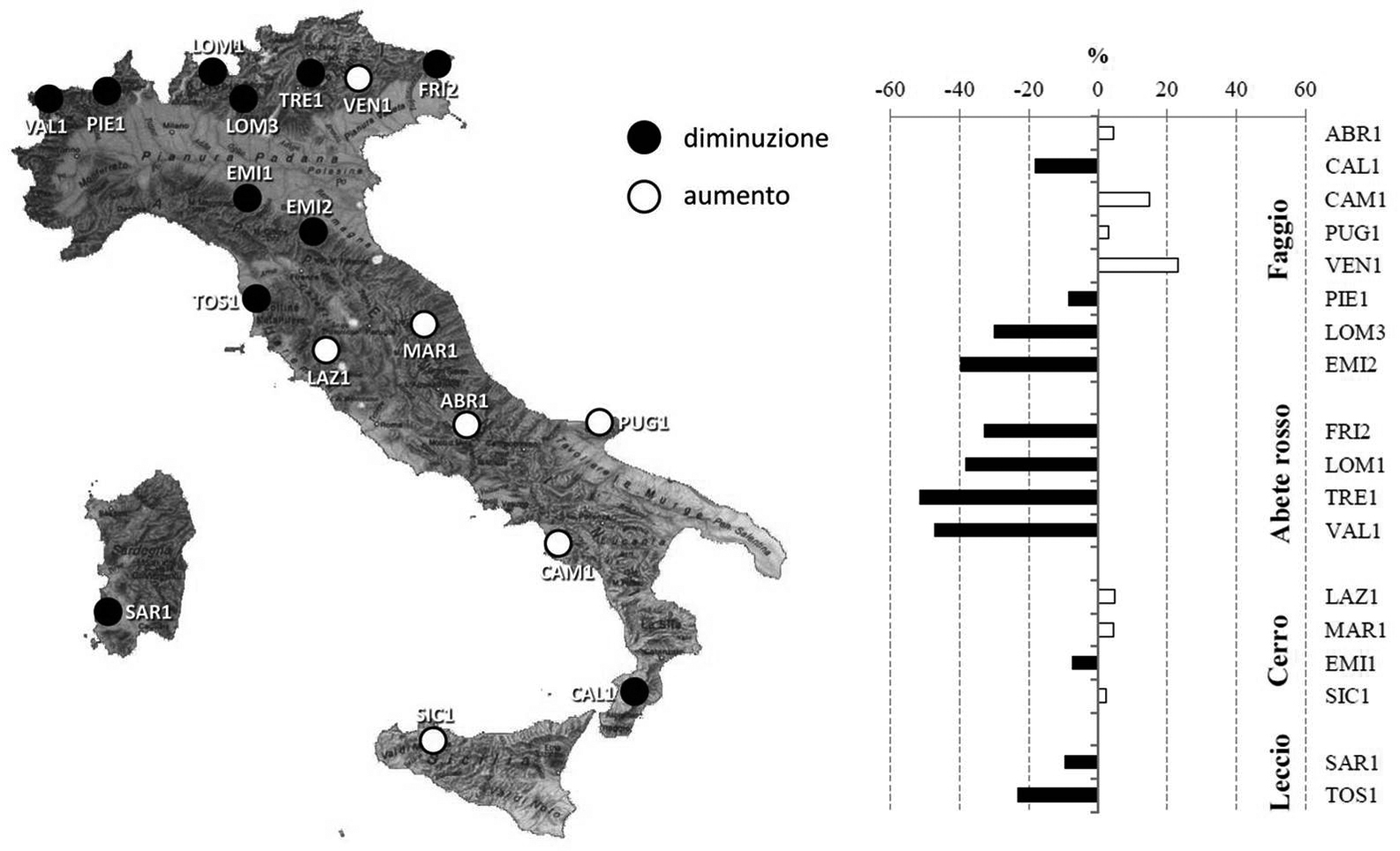
Il secondo fenomeno riguarda la regione alpina e le aree di conifere (abete rosso) collocate a quote medio-elevate nell’intervallo di monitoraggio 2005-2009 (Fig. 6). L’analisi climatica della regione (Tab. 1) ha evidenziato l’occorrenza nell’ultimo decennio di ripetute deviazioni nell’andamento stagionale delle temperature e della piovosità, in accordo con i modelli IPCC per la stessa area geografica.
Fig. 6 - Distribuzione dei livelli di riduzione dell’accrescimento attraverso la rete. Variazione nell’intervallo 2005-2009 rispetto al periodo 2000-2004 (da: [3]).
Tab. 1 - Deviazioni della temperatura media e della precipitazione rispetto alle medie del periodo 1996-2009 nella regione alpina. (+/-): aumento/diminuzione > 50%; (++/--): aumento/diminuzione > 100%; (+++/---): aumento/diminuzione > 150% (da: [3]).
| Variabile | Stagione | 2000 | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperatura | primavera | 0 | ++ | 0 | 0 | 0 | 0 | 0 | ++ | 0 | 0 |
| estate | 0 | 0 | 0 | +++ | 0 | 0 | 0 | 0 | 0 | 0 | |
| autunno | 0 | 0 | 0 | 0 | 0 | 0 | + | 0 | 0 | 0 | |
| inverno | 0 | 0 | 0 | - - | 0 | - | - | ++ | + | - | |
| Precipitazioni | primavera | 0 | +++ | ++ | - | ++ | 0 | 0 | 0 | 0 | 0 |
| estate | 0 | 0 | 0 | - | 0 | 0 | 0 | 0 | 0 | 0 | |
| autunno | +++ | 0 | +++ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| inverno | - | +++ | +++ | - | ++ | - | 0 | 0 | 0 | ++ |
Il livello locale
A questa scala, un caso interessante è relativo al sito EMI1, collocato sul margine meridionale della Valle Padana (Fig. 7). Qui convivono due querce decidue, rovere e cerro, con esigenze idriche diverse. Il cerro più xero-tollerante, la rovere più esigente di umidità atmosferica e sensibile allo stress idrico. La disponibilità in questo sito di una serie climatica trentennale ([2]) ha permesso di verificare ripetute deviazioni annuali nella temperatura media dell’aria e nella quantità di precipitazione. Le deviazioni di temperatura di segno positivo prevalgono nell’ultima decade e si traducono nella formazione di un trend significativo di aumento; quelle di precipitazione si distribuiscono con variazioni annuali marcate di segno opposto lungo tutto il periodo di osservazione, ma non costituiscono nessun trend. La risposta in accrescimento radiale per le due specie (Fig. 8) descrive il declino incrementale della rovere a partire dall’inizio degli anni ’90 ([4]). Le conseguenze a livello di popolazione nel periodo monitorato sono importanti: la rovere, specie prevalente al 1996 diventa specie secondaria al cerro in termini quantitativi per il ridotto accrescimento e per il livello di mortalità elevato (Fig. 9). La maggiore sensitività al ripetuto stress idrico estivo ha mutato in un intervallo breve un carattere relativamente stabile quale la composizione del bosco.
Fig. 7 - Collocazione geografica e struttura del soprassuolo (sito EMI1).

Fig. 8 - Cronologie anulari per rovere e cerro (sito EMI1).
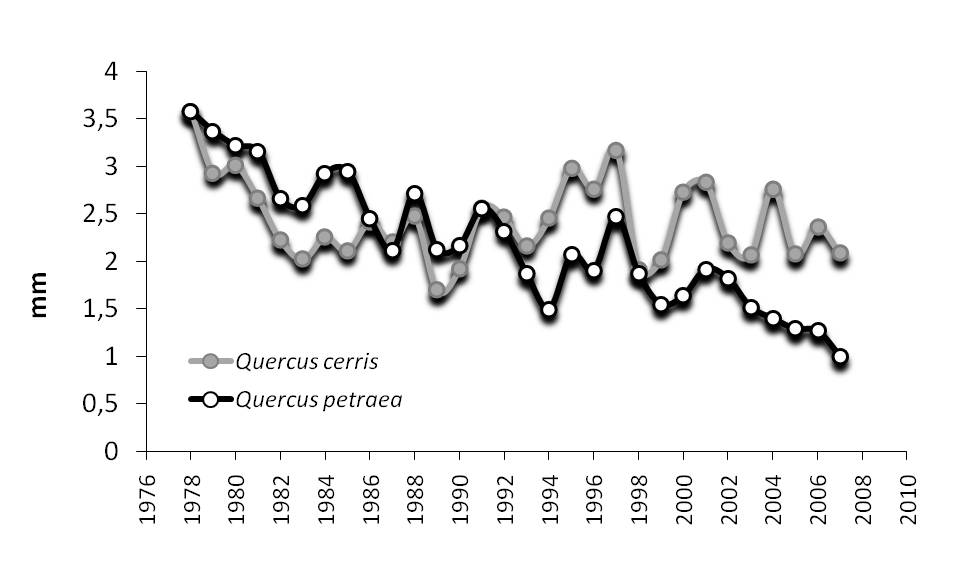
Fig. 9 - Andamento della mortalità (numero di individui) per rovere e cerro attraverso i successivi intervalli monitorati (sito EMI1).
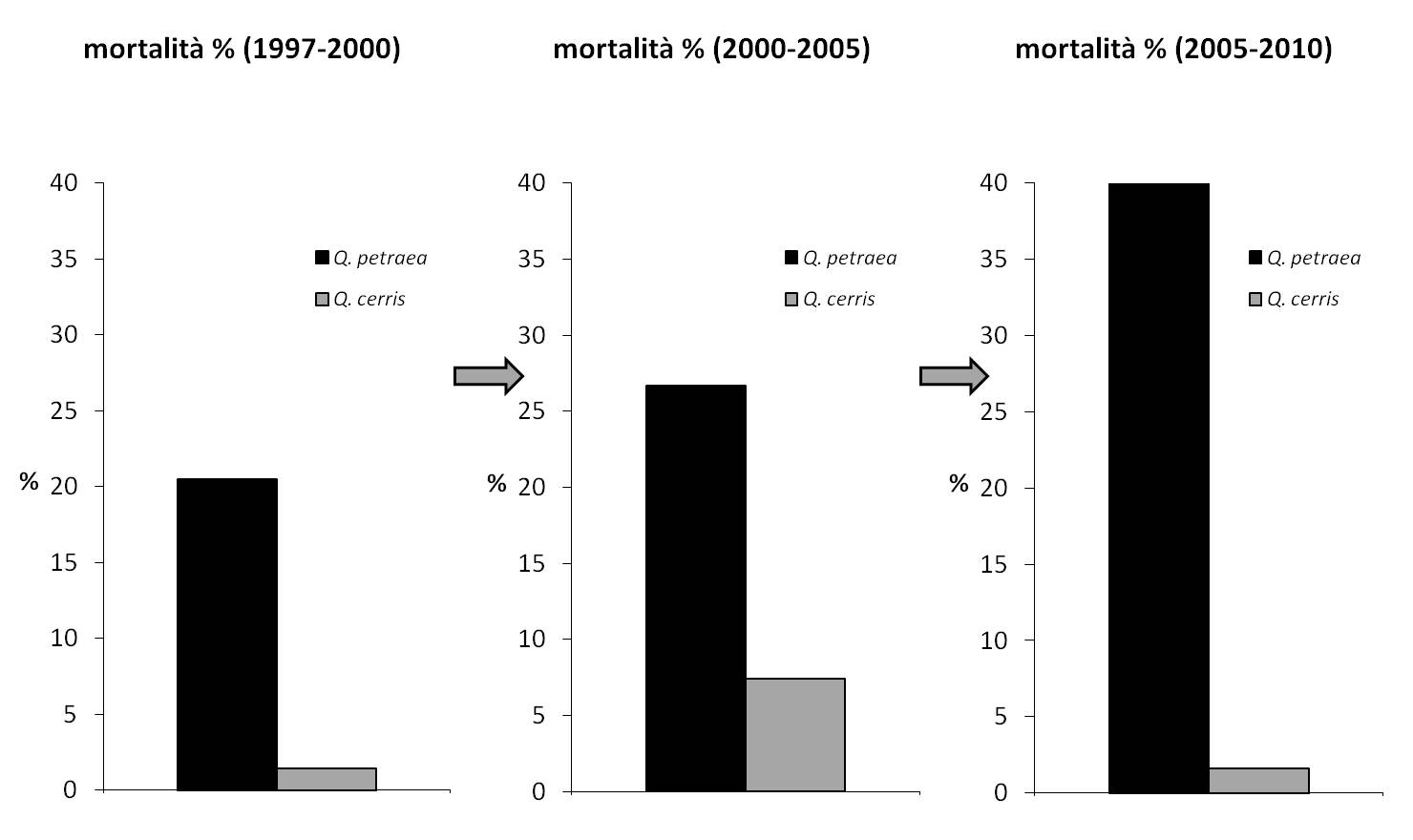
Questi casi evidenziano scale di lettura diverse dalla rete di livello II. I risultati delle analisi svolte per le singole Azioni e dell’analisi complessiva sono a oggi stati pubblicati sulle riviste di settore e in una serie di volumi dedicati al programma di monitoraggio ([11], [19], [12], [13], [14]).
Azioni complementari sviluppate nell’ultimo biennio 2009-2010
Il progetto FutMon ha aumentato le scale di rilievo (insieme-strato-individuo), introdotto intervalli di ripetizione più ridotti (annuale e intra-annuale), integrato protocolli complementari sui core-plot dove si è concentrato nell’ultimo periodo lo sviluppo dell’attività di monitoraggio.
Per l’Azione “accrescimento”, il rilievo ogni 5 anni a livello plot è stato integrato con quello annuale e intra-annuale su un campione random di quindici alberi della/e specie prevalente/i ubicati nel piano dominante. Il rilievo è svolto tramite bande di circonferenza con accuratezza di 0.01 cm e letture bi-settimanali lungo tutta la stagione vegetativa. Nella Fig. 10, è riportato il grafico relativo al plot EMI1-2010 (curve medie), dove entrambe le querce sono monitorate. Si evidenziano l’andamento sincrono dell’incremento radiale delle due specie e si conferma la ridotta capacità di accrescimento corrente della rovere rispetto al cerro.
Fig. 10 - Andamento incrementale intra-annuale per rovere e cerro (anno 2010 - sito EMI1).
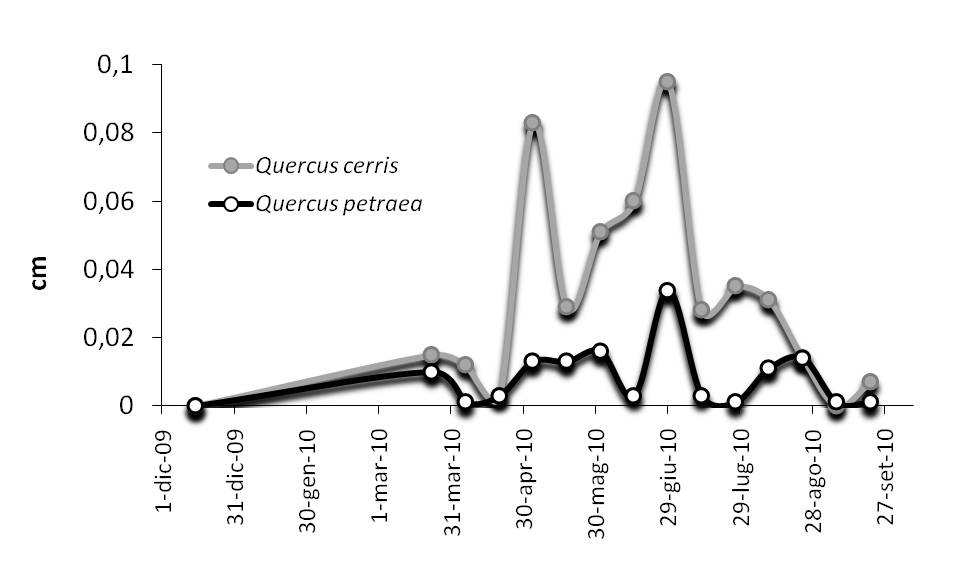
Questo protocollo è integrato, sullo stesso campione, dall’osservazione fenologica e di eventuali danni di tipo biotico ed abiotico alle chiome; a livello plot dalla determinazione di indici sintetici di produttività annuale: produzione di lettiera fogliare e totale, chimica della lettiera, indice di area fogliare. Le informazioni raccolte sullo stesso campione e sito nella medesima finestra temporale e le relazioni tra le variabili misurate consentono di definire la dinamica stagionale-annuale.
Prospettive del programma di monitoraggio
La serie dei dati raccolti dalla metà degli anni ’90 inizia ad essere abbastanza lunga per trarre alcuni risultati su stato e cambiamenti in atto di alcuni attributi chiave ([15], [16]). L’implementazione recente del protocollo su un sotto-insieme di aree target consente oggi una lettura di maggiore dettaglio. A livello nazionale, il finanziamento di queste Azioni da parte del Corpo Forestale dello Stato negli anni di interruzione del programma comune europeo ha consentito di mantenere la continuità delle misure e osservazioni, requisito fondamentale di un programma di monitoraggio intensivo. Soltanto la permanenza del tema “Monitoraggio delle Foreste” nell’Agenda europea potrà tuttavia, già nel breve periodo, garantire il finanziamento e quindi la prosecuzione del programma a livello continentale attraverso protocolli comuni.
Ringraziamenti
Si ringraziano i molti colleghi che partecipano a questa attività complessa di rilievo, ordinamento, validazione ed elaborazione dei dati a livello nazionale. In particolare, il Personale del CFS a livello centrale e quello operativo sui siti, il Personale delle Regioni, delle Province autonome, degli Enti Parco. Tutti, per la disponibilità, competenza e professionalità che hanno reso possibile in tutti questi anni la realizzazione e la continuità del programma.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar