Foliar nutrients in Italian forests: results from the 1995-2009 monitoring network sites CONECOFOR
Forest@ - Journal of Silviculture and Forest Ecology, Volume 10, Pages 113-121 (2013)
doi: https://doi.org/10.3832/efor1019-010
Published: Nov 13, 2013 - Copyright © 2013 SISEF
Research Articles
Guest Editors: LIFE FUTMON Project
« Further Development and Implementation of an EU-level Forest Monitoring System »
Collection/Special Issue: Luca Salvati (CRA-RPS, Roma)
Abstract
Foliar nutrients in Italian forests: results from the 1995-2009 monitoring network sites CONECOFOR. In 1995, the Italian Forest Service (CFS) established the National Network for Forest Ecosystem Monitoring (CONECOFOR) with the aim to study the ecological interactions among structural and functional components of forest stands. In the 31 permanent plots of the CONECOFOR Network, surveys are carried out including vegetation, trees crown condition, chemistry of leaves and soils, tree growth, atmospheric deposition, climate, microclimate, ozone and biodiversity. These surveys are carried out by CFS in collaboration with several scientific institutions. In such context, IBAF/CNR is specifically involved in the investigation of foliage chemical content. The foliar nutritional status was analyzed in 25 sites, taking as a reference the values published by the ICP-Forests and those published in the literature. The study involved 7 forest species (Quercus cerris, Q. petraea, Q. robur, Q. ilex, Fagus sylvatica, Carpinus betulus and Picea abies) investigating the concentrations of the major macronutrients as nitrogen (N), phosphorus (P), potassium (K), magnesium (Mg), sulfur (S) and calcium (Ca) and the N/P ratio. In beech, N, S and K were found in quite high concentrations, though without showing imbalances due to concentration excesses. Ca and Mg are present in very high concentrations, highlighting the possibility of imbalances, especially in relation to the other nutrients. For these two nutrients, spruce shows substantially balanced conditions as also for N/P ratio. For deciduous Oaks nutrients arewithin the parameters of normality, with the exception of Ca. This has been observed in 4 out of the 6 studied sites, with potential indication of nutritional imbalances related to Calcium. Nutrient concentrations in Holm oak are within the average of published data, except for Ca and Mg of site TOS-1, the former being slightly low, the latter being unusually high, in relation to the peculiar soil Mg content of the above site. Considering the coefficient of variation (CV), the variability of the different elements, taking in account all species, presents the following ranking: K ≈ P > Ca> Mg ≈ N / P > S> N. On those sites presenting the longer time-series (12 out of the 25 studied sites), a more detailed analysis has been performed in order to verify the existence of trends along the monitoring period for each nutrient. Calcium, Mg and N/P show a steady decrease during the considered time span. Such trend is less marked for Ca than for Mg and N/P. Phosphorus and K show an increasing trend while no common trends have been highlighted for N and S. Overall, the study provides relevant data on status and dynamics of nutrients in Italian forest ecosystems.
Keywords
Foliar Nutrients, Macronutrient, Nutritional Status, Nutrient Classes, Nutrient Trends, Foliar Chemistry
Introduzione
La composizione chimica delle foglie è un indicatore importante dello stato nutrizionale dell’albero. La disponibilità di ogni singolo elemento è fondamentale per la vita della pianta, perché ognuno di questi svolge un ruolo importante nel metabolismo: l’azoto (N) è per esempio il principale costituente delle proteine ([25]) ed, insieme al fosforo (P), è l’elemento maggiormente limitante per la crescita della vegetazione terrestre ([19]). Il P è presente anche negli acidi nucleici, nell’ATP e nelle membrane cellulari ([26]) ed ha inoltre funzioni specifiche nella sintesi proteica. Il calcio (Ca) è presente nelle componenti strutturali delle foglie, principalmente come costituente delle pareti cellulari ([26]). Il potassio (K) ha un ruolo nelle funzioni stomatiche, di assorbimento e trasporto fra comparti. Questo elemento è anche attivo nel determinare alcune importanti attitudini delle piante come la resistenza al gelo ed alla siccità ([27], [13], [11], [22], [3]). Alcune funzioni importanti della pianta come la fotosintesi sono strettamente collegate al magnesio (Mg) ed alla disponibilità di N, mentre i processi di produzione del legno sono influenzati dalla disponibilità nel terreno di macro e micronutrienti e dalla loro concentrazione nelle foglie ([11], [22], [3]).
È ampiamente dimostrato che il contenuto di molti micro e macroelementi fogliari è proporzionale alla loro disponibilità nel terreno. ([16], [14], [17]). Tuttavia la concentrazione di nutrienti nelle foglie non è comunque influenzata unicamente dalla disponibilità edafica, ma da diversi altri fattori. La capacità di assorbimento delle piante per esempio svolge un ruolo importante. Questa è influenzata da fattori quali le caratteristiche del sito, l’intensità della gestione forestale, l’entità delle deposizioni atmosferiche: l’aumento della disponibilità di un elemento può indurre uno squilibrio tra esso e gli altri elementi, che possono diventare relativamente insufficienti alle esigenze della pianta. Questo è frequentemente osservato tra N e P, K e Mg, a seguito di un aumento di deposizione di N ([9]).
Un eccesso, una carenza o uno squilibrio di nutrienti fogliari può influenzare lo stato di salute delle foreste, compromettendo il loro equilibrio ([28], [10], [6]).
Dal 1986, l’International Co-operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests (ICP-Forests) collabora a stretto contatto con l’Unione Europea al fine di rilevare l’influenza dei maggiori fattori di stress degli ecosistemi forestali. Un monitoraggio intensivo si svolge su 860 aree permanenti collocate in foreste che rappresentano gli ecosistemi forestali più importanti del continente. Su 767 di queste si effettua biennalmente un campionamento fogliare al fine di monitorare il contenuto dei maggiori nutrienti (The Condition of Forests in Europe 2008 Executive Report). Grazie a questo programma di monitoraggio su larga scala, un database dei nutrienti fogliari è ormai disponibile per i più importanti Paesi europei.
In Italia le prime venti aree sono state installate nel 1995. Tra il 1999 e il 2003 la copertura geografica ed ecologica della rete è stata integrata mediante l’inserimento di undici nuove aree nel Nord e Centro Italia, aumentando il numero di siti per specie (leccio) e nuovi ambienti (formazioni planiziali, ecosistemi di alta quota di larice e peccio). Al momento la rete è costituita da 31 aree ([5]).
In questo lavoro, basato sull’analisi dei principali nutrienti fogliari, presentiamo i dati sullo stato nutrizionale degli ecosistemi forestali in Italia. Molti dei siti studiati sono stati anche inclusi nella rete di monitoraggio intensivo del progetto “LIFE+ FutMon” (⇒ http://www.futmon.org/). Il progetto, coordinato dal Thunen Institute tedesco, ha coinvolto 38 beneficiari (rappresentanti quasi tutti gli Stati membri dell’UE) e ha contribuito al UN/ECE International Cooperative Programme of Forests (ICP-Forests).
Il presente studio, basato su dati di medio/lungo termine (1995-2009), si pone come un punto di riferimento per quanto riguarda i nutrienti fogliari in ecosistemi forestali nel nostro paese.
Gli obiettivi specifici sono: (a) lo studio dello status nutrizionale delle foreste monitorate; (b) l’analisi degli andamenti nel periodo studiato. Per quanto riguarda lo status nutrizionale si valuterà: (i) la concordanza tra i valori italiani e le soglie per la nutrizione fissate a livello europeo ([24]); (ii) se eventuali differenze siano legate a squilibri nutrizionali o a caratteristiche sito-specifiche (i valori soglia si basano prevalentemente su dati provenienti dal centro Europa); (iii) l’opportunità di inserire nel confronto un’importante specie mediterranea quale il leccio (Quercus ilex L), non contemplata nelle tavole europee dei valori soglia ([24]). Riguardo invece il punto (b) verrà indagata l’esistenza di andamenti e l’eventuale loro comportamento.
Metodi
L’analisi dei nutrienti fogliari è tra le indagini obbligatorie nei siti di livello II della rete ICP-Forests ed è svolta ogni due anni. Il programma di monitoraggio in Italia è iniziato nel 1995 su 20 siti e, durante un periodo di 14 anni, il numero di aree di monitoraggio permanenti e di specie esaminate è aumentato ([2]). Le aree di monitoraggio permanenti attualmente attive in Italia sono 31 per un totale di 11 specie (Fig. 1).
Fig. 1 - Anni campionati per i siti della rete di monitoraggio CONECOFOR/FutMon. In blu i siti scartati dal presente studio (anni di campionamento non sufficienti); in verde quelli presi in considerazione per il solo studio dello stato delle concentrazioni (limitato numero di anni di campionamento); in rosso quelli presi in considerazione sia per le concentrazioni che per lo studio degli andamenti.
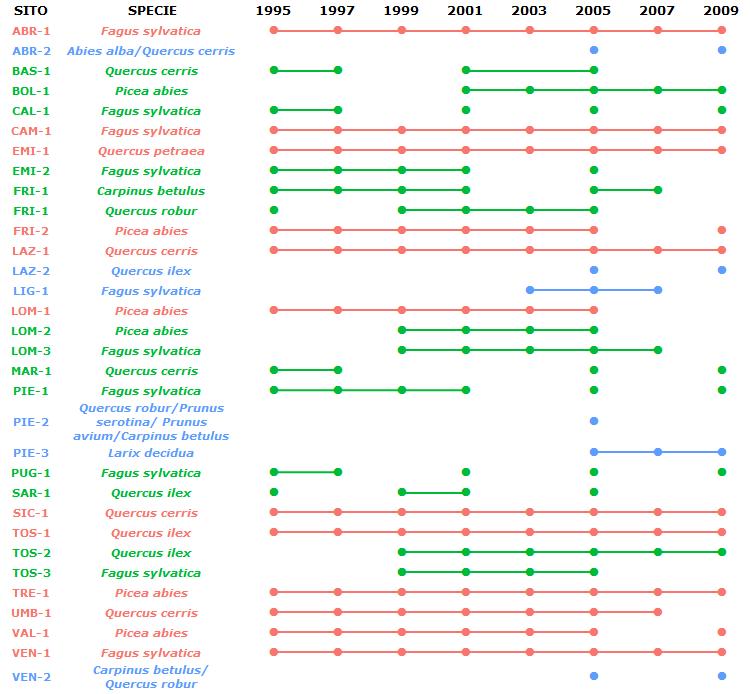
In questo lavoro è stato considerato il periodo di campionamento 1995-2009. Sono stati analizzati i sei principali nutrienti, ossia N, P, Ca, Mg, K e S. La procedura di campionamento e di analisi fogliare segue i protocolli descritti nel manuale adottato a livello europeo ([18]).
Una rivisitazione del protocollo in oggetto è stata eseguita a seguito di quanto discusso nel Forests Expert Meeting di Varsavia (Ottobre 2009) ed in quello finlandese di Tampere, (Febbraio 2010). Su proposta del gruppo Eco.Di.S.F-IBAF-CNR (responsabile della raccolta e delle analisi fogliari) è stato apportato un proficuo aggiornamento volto a standardizzare le metodologie di prelievo, conservazione e spedizione del materiale campionato ([18]). Una difformità di campionamento fra paese e paese potrebbe infatti rendere meno confrontabili i risultati ottenuti da ogni nazione e, ancor più grave, indurre anomalie nel dato che, in sede di interpretazione dello stesso, potrebbero essere attribuite ad altri fattori.
Il campione viene prelevato dal terzo superiore della chioma. Si raccolgono foglie sviluppatesi in condizioni di piena luce da piante appartenenti al piano dominante e codominante del popolamento.
Le specie decidue vengono campionate durante il picco della stagione di crescita, prima dell’inizio dei fenomeni di senescenza autunnale, mentre il prelievo nelle conifere avviene durante il periodo di dormienza. Nel leccio il campionamento può avvenire indifferentemente in piena estate come nel tardo autunno.
Il campionamento avviene su un numero minimo di cinque alberi e, quando possibile, interessa le stesse piante per tutto il periodo di campionamento.
In laboratorio i campioni vengono trattati secondo il seguente protocollo: (i) asciugatura in stufa a 80 °C per 48 ore; (ii) macinatura; (iii) mineralizzazione (per P, Ca, Mg e K); (iv) analisi mediante uno dei metodi approvati, proposti dal manuale ICP-Forests, rielaborato e integrato durante il progetto LIFE+ FutMon.
I laboratori che nel periodo monitorato hanno effettuato le analisi, hanno aderito annualmente ad un Ring Test allo scopo di testare e certificare la qualità dei propri strumenti e delle proprie procedure di analisi ([8]).
Dei 31 siti afferenti alla rete per lo studio delle concentrazioni, sono stati presi in considerazione 25 siti: quelli con almeno tre campionamenti e/o ritenuti di maggiore interesse (riportati in verde ed in rosso nella Fig. 1). Per lo studio degli andamenti sono stati utilizzati i dati di 12 siti ovvero quelli con almeno 5 campionamenti nell’arco del periodo di studio (riportati in rosso in Fig. 1).
I principali elementi nutritivi (N, P, K, Ca, Mg, S) sono stati scelti come indicatori dello stato nutrizionale delle aree di monitoraggio permanenti. Poiché la concentrazione assoluta di nutrienti può non indicare l’effettiva esistenza di eventuali squilibri nutrizionali ([12], [7]), abbiamo anche considerato il rapporto N/P. Sono stati scelti questi due elementi in quanto sono generalmente considerati quelli maggiormente limitanti lo sviluppo della vegetazione terrestre e, dato questo loro importante ruolo, sono disponibili in letteratura database di riferimento riportanti i valori di questo rapporto ([19]). Le concentrazioni dei nutrienti ottenute dallo studio dei siti italiani, sono state confrontate con i valori soglia proposti a livello europeo per il monitoraggio delle condizioni fogliari delle foreste ([24]).
Tali valori stabiliscono tre classi di concentrazione: bassa, media ed alta. In generale la classe media si riferisce a condizioni di equilibrio mentre le altre due classi indicano possibilità di sbilanci nutrizionali dovuti a carenza od eccesso (possibile tossicità) del nutriente ([24]).
Tali classi contemplano le più importanti specie dell’areale centro europeo. Specie con areali più marginali (seppur importanti per le regioni in cui vegetano), come ad esempio il leccio, non sono contemplate. Lavori precedenti a questo ([14]) hanno già evidenziato come non sia prudente considerare per il leccio i valori soglia relativi al gruppo delle querce caducifoglie. Nel presente lavoro, per questa specie il confronto viene effettuato facendo riferimento ai dati presenti in letteratura.
Riguardo infine l’approccio statistico usato per la verifica della presenza di andamenti temporali dei nutrienti, sono state utilizzate regressioni lineari (ordinary least squares) rispetto al tempo per le quali, qualora significative, sono stati determinati i coefficienti angolari. L’analisi statistica è stata realizzata con il programma SYSTAT (versione 11).
Risultati e discussione
Il presente studio interessa (Fig. 1) nove siti di faggio (Fagus sylvatica L.) distribuiti lungo un gradiente latitudinale Nord-Sud che va dal sito VEN-1 (Veneto) a quello di CAL-1 (Calabria). L’abete rosso (Picea abies Karst.) è rappresentato da sei siti distribuiti lungo l’arco alpino con VAL-1 sito più ad Ovest e FRI-2 quello più ad Est. I siti di querce caducifoglie sono in totale sette e nello specifico cinque di Cerro (Quercus cerris L.), uno di Rovere (Quercus petraea L.) ed uno di Farnia (Q. robur L.). Il leccio (Quercus ilex L.) è presente in tre siti: due in Toscana ed uno in Sardegna.
Studio delle concentrazioni
Nella Fig. 2 vengono mostrati i box plot delle concentrazioni, espresse in mg g-1 e del rapporto N/P per ogni sito e per singolo elemento. Questi si riferiscono ai valori medi dei cinque alberi per sito e degli anni del periodo di studio.
Fig. 2 - Box plot delle concentrazioni (mg g-1) e del rapporto N/P per sito per il periodo di studio (1995-2009). La simbologia proposta in figura è in accordo con quanto proposto da Tukey ([15]): i limiti destro e sinistro del box corrispondono al primo e terzo quartile (rispettivamente 25 ° e 75° percentile). La barra a destra si estende dal limite al valore più alto che è pari a 1.5 · IQR del limite, dove IQR è il range inter-quartile. I dati oltre la fine delle barre sono valori anomali e vengono indicati come punti.
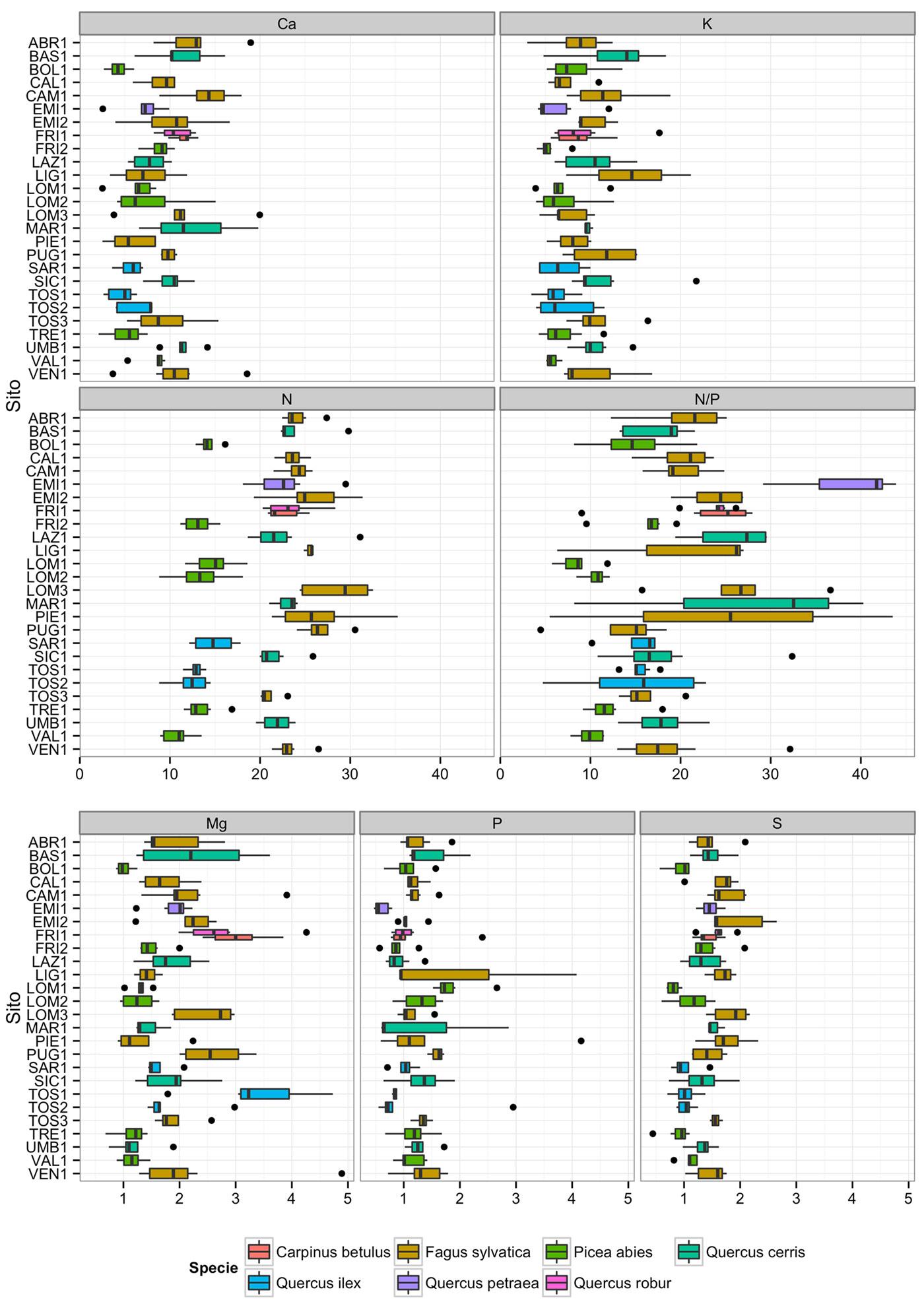
Dall’analisi del contenuto analizzando la Fig. 2 per specie o per gruppi affini, si possono evidenziare i seguenti gradienti di concentrazione dei nutrienti:
- Faggio: N > Ca ≈ K >> Mg > S ≈ P
- Querce caducifoglie: N > Ca ≈ K >> Mg > S > P
- Abete rosso: N > K ≈ Ca >> P ≈ Mg > S
- Leccio: N > K > Ca > Mg > S ≈ P
Da questo tipo di osservazione, finalizzata ad un confronto fra le diverse specie, emerge come, tranne il leccio, gli altri gruppi presentino concentrazioni di Ca e K molto vicine fra loro.
Mettendo a confronto quanto in media osservato nel periodo di indagine con i valori soglia di riferimento proposti a livello Europeo per il monitoraggio delle condizioni fogliari delle foreste, emerge quanto riportato in Tab. 1.
Tab. 1 - Percentuale di siti CONECOFOR le cui concentrazioni fogliari, per ciascun elemento e ciascuna specie (Fig. 2), che ricadono nelle classi soglia proposte da Stefan et al. ([24]). Classe di concentrazione: (1): bassa; (2): media; (3): alta.
| Elemento | Faggio - Classe soglia | Abete rosso - Classe soglia | Querce decidue - Classe soglia | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | |
| N | - | 56 | 44 | 17 | 83 | - | - | 100 | - |
| S | - | 56 | 44 | 34 | 66 | - | - | 100 | - |
| P | 56 | 33 | 11 | - | 83 | 17 | - | 86 | 14 |
| Ca | - | 11 | 89 | - | 33 | 67 | - | 29 | 71 |
| Mg | - | 11 | 89 | - | 83 | 17 | - | 71 | 29 |
| K | - | 44 | 56 | - | 100 | - | - | 57 | 43 |
| N/P | - | 78 | 22 | - | 100 | - | - | 57 | 43 |
Per il faggio si può osservare come la concentrazione di tutti gli elementi, salvo il P, sia equilibrata ma con un’importante percentuale di siti nella classe alta (classe 3). Per Ca, Mg e K la percentuale di siti in classe 3 è decisamente elevata il che potrebbe determinare un rischio di squilibrio. La situazione per l’abete rosso è più equilibrata seppure anche per questa specie la concentrazione di Ca sia molto elevata. Per quanto riguarda le querce decidue si può osservare ancora una concentrazione di Ca particolarmente elevata mentre risulta equilibrata per il resto degli elementi.
Nel caso del Ca è stata rilevata una concentrazione fogliare sensibilmente più elevata in siti con suoli di matrice carbonatica rispetto a quelli di altra natura. Un’osservazione mirata sui suoli dei siti con faggio ha rilevato che quattro siti, dei cinque che presentavano le più alte concentrazioni fogliari di tale elemento, presentano matrici di natura carbonatica (dati non mostrati).
I valori soglia Europei non forniscono dati ufficiali di riferimento per il leccio. Vista l’importanza di questa specie nel contesto Mediterraneo è stata comunque operata una comparazione con quanto osservato in letteratura.
Nel range di dati provenienti principalmente da Spagna e Grecia ([4], [23], [20], [1], [21]) abbiamo osservato che i siti di SAR-1 e TOS-2, per quanto riguarda il N, sono rispettivamente il sito con più elevata e con minore concentrazione. Entrambi questi siti presentano valori di P vicini alle stazioni che in tale range si configurano come le più ricche di tale elemento, mentre il sito TOS-1 presenta , per quanto riguarda il N, valori in linea con la media.
Tutti e tre i siti italiani presentano valori di Ca bassi (tutti e tre vegetano su substrato non calcareo).
Riguardo il Mg, infine, il sito TOS-1 presenta una concentrazione particolarmente elevata (la concentrazione di tale elemento nel suolo è pari a 20-30 meq l-1), ben al di sopra di ogni altro sito osservato, mentre le concentrazioni per gli altri due siti si collocano nella fascia bassa del range considerato.
Studio degli andamenti
I valori interannuali di ciascun sito evidenziano un andamento nel tempo. Nella Tab. 2 si illustra come in molti casi questo sia significativamente interpretabile tramite una retta (colonna 2) la cui pendenza ed andamento vengono riportati nella colonna 4.
Tab. 2 - Andamenti nel tempo delle variazioni di concentrazione dei singoli elementi, indicazione della loro significatività, entità e direzione delle tendenze di lungo termine (1995-2009). (*): P < 0.1; (**): P < 0.05; (***): P < 0.01.
| Sito | Elemento | Regressione lineare | |
|---|---|---|---|
| Significatività | Valore Coefficiente | ||
| ABR1 | N | - | -0.079 |
| S | - | 0.006 | |
| P | * | -0.035 | |
| Ca | ** | -0.436 | |
| Mg | *** | -0.155 | |
| K | - | 0.056 | |
| N/P | - | 0.390 | |
| CAM1 | N | *** | -0.286 |
| S | *** | 0.057 | |
| P | - | 0.013 | |
| Ca | *** | -0.647 | |
| Mg | *** | 0.222 | |
| K | *** | 0.247 | |
| N/P | *** | -0.494 | |
| EMI1 | N | * | 0.423 |
| S | ** | -0.025 | |
| P | ** | 0.015 | |
| Ca | *** | -0.583 | |
| Mg | *** | -0.068 | |
| K | - | 0.126 | |
| N/P | - | -0.187 | |
| FRI2 | N | - | -0.029 |
| S | - | -0.032 | |
| P | ** | 0.038 | |
| Ca | - | -0.144 | |
| Mg | - | -0.020 | |
| K | *** | 0.207 | |
| N/P | *** | -0.654 | |
| LAZ1 | N | ** | -0.510 |
| S | *** | -0.590 | |
| P | - | 0.011 | |
| Ca | - | 0.015 | |
| Mg | - | 0.011 | |
| K | - | 0.093 | |
| N/P | *** | -0.832 | |
| LOM1 | N | ** | 0.354 |
| S | ** | 0.016 | |
| P | *** | 0.090 | |
| Ca | - | -0.149 | |
| Mg | * | 0.028 | |
| K | *** | 0.589 | |
| N/P | - | -0.097 | |
| SIC1 | N | - | -0.072 |
| S | *** | -0.090 | |
| P | *** | 0.086 | |
| Ca | - | -0.098 | |
| Mg | - | -0.034 | |
| K | * | 0.177 | |
| N/P | *** | -0.123 | |
| T0S1 | N | - | 0.031 |
| S | - | 0.014 | |
| P | - | -0.004 | |
| Ca | - | -0.130 | |
| Mg | *** | -0.161 | |
| K | - | 0.096 | |
| N/P | - | 0.071 | |
| TRE1 | N | - | 0.002 |
| S | - | -0.008 | |
| P | * | 0.029 | |
| Ca | *** | -0.471 | |
| Mg | - | 0.013 | |
| K | - | 0.095 | |
| N/P | *** | -0.501 | |
| UMB1 | N | *** | -0.374 |
| S | * | 0.017 | |
| P | - | -0.009 | |
| Ca | - | 0.187 | |
| Mg | - | -0.016 | |
| K | *** | 0.407 | |
| N/P | - | -0.097 | |
| VAL1 | N | - | 0.084 |
| S | *** | 0.050 | |
| P | - | 0.015 | |
| Ca | *** | -0.434 | |
| Mg | *** | -0.054 | |
| K | - | 0.088 | |
| N/P | - | -0.048 | |
| VEN1 | N | - | -0.056 |
| S | *** | 0.047 | |
| P | *** | -0.056 | |
| Ca | *** | -0.733 | |
| Mg | ** | -0.134 | |
| K | - | 0.183 | |
| N/P | *** | 0.997 | |
Il faggio è sicuramente la specie che maggiormente evidenzia chiari andamenti temporali delle concentrazioni fogliari e, in linea di massima, per ogni sito almeno un elemento presenta un andamento significativo nel periodo monitorato. In particolare nel sito CAM-1 (faggio) si delineano andamenti per sei elementi, mentre nei siti EMI-1 (cerro-rovere) e VEN-1 (faggio) ciò avviene per cinque elementi e così via decrescendo fino al sito TOS-1 (leccio) che presenta un andamento significativo per il solo Mg.
Se si osserva la situazione dal punto di vista dell’elemento, S e P evidenziano un andamento significativo rispettivamente in otto e sette siti, mentre Ca, Mg ed il rapporto N/P in cinque.
Nei casi in cui è emersa la presenza di andamenti, sarà possibile improntare studi mirati a comprendere se il fenomeno derivi da disturbi o da altri fattori. Importante in questi casi è stabilire se la concentrazione oggetto di studio stia aumentando o diminuendo nel tempo.
Dalla Tab. 2 si osserva che i siti che presentano concentrazioni con andamento crescente (20) e quelli con andamento decrescente (24) sono approssimativamente equivalenti.
A livello di elemento: per l’N e lo S non si evince una chiara tendenza come risulta invece per gli altri elementi. In particolare si osservano i seguenti andamenti:
- crescita per il Ca il Mg ed il rapporto N/P;
- diminuzione per P e K.
Conclusioni
Il monitoraggio ha consentito di fornire lo stato dei principali nutrienti fogliari in alcune delle tipologie forestali italiane. Nell’arco del periodo monitorato si è osservata una situazione eterogenea.
La comparazione dei dati ottenuti con i valori soglia europei e di letteratura (per il leccio) ha evidenziato condizioni mediamente equilibrate con alcune peculiarità quali la situazione del faggio che è risultata, per alcuni nutrienti, piuttosto ricca con possibilità di rischio di squilibrio da eccesso. Degna di nota è inoltre la situazione del Ca che presenta valori sempre molto alti per faggio, abete rosso e querce caducifoglie e bassi per il leccio. A tal proposito, per alcuni nutrienti risulta chiara la relazione con la disponibilità degli stessi nei suoli (osservato per il Ca ed il Mg).
Si sono osservati andamenti interannuali anche se non è stato evidenziato un andamento comune per tutti i siti monitorati.
Ulteriori approfondimenti, finalizzati a mettere in relazione le condizioni nutrizionali dei vari siti con altre indagini strutturali e funzionali (ad esempio soluzioni circolanti nei suoli, accrescimento degli alberi, ecc.), saranno oggetto di prossimi studi per ampliare la comprensione delle relazioni ecologiche delle foreste italiane.
Ringraziamenti
Si ringraziano: Il Corpo Forestale dello Stato ed i responsabili delle aree di monitoraggio permanente della rete di monitoraggio CONECOFOR per il supporto fornito alle operazioni di campo. Il Sig. Giorgio Poggio (ISE-CNR, Pisa), il Sig. Domenico Mastroianni (IRSA-CNR, Monterotondo Scalo - Roma), la Dott.ssa Valentina Iori, la Dott.ssa Lucia Pietrosanti ed il Sig. Pierangelo Bertolotto (IBAF-CNR, Monterotondo Scalo - Roma) per il supporto alle operazioni analitiche. Il Dott. Mario Cammarano, il Sig. Pierangelo Bertolotto ed il Dott. Domenico Serafini (IBAF-CNR, Monterotondo Scalo - Roma) per il prezioso contributo fornito alla fase di campionamento in chioma come tree-climbers.
References
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar