Evolutionary dynamics of birch (Betula aetnensis Rafin) coppices on the Mount Etna (Sicily)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 11, Pages 52-65 (2014)
doi: https://doi.org/10.3832/efor1191-011
Published: Apr 18, 2014 - Copyright © 2014 SISEF
Research Articles
Abstract
Evolutionary dynamics of birch (Betula aetnensis Rafin) coppices on the Mount Etna (Sicily). The aim of this paper is to evaluate the dynamics of Etna birch stands (Betula aetnensis Rafin) following the cessation of silvicultural activities in the Etna Regional Park (Sicily). We investigated forest structure, natural regeneration, vegetation and deadwood in different forest types. Our findings highlighted three different dynamics for birch populations: stable birch stands in the high mountain area which might represent an edapho-climax forest; progressive dynamic birch stands in the intermediate mountain area, showing a gradual depletion of birch and a concomitant replacement with monospecific stands (calabrian pine, beech, oaks) or mixed ones (with birch); pure birch stands (typical) that tend to be regressive - especially under stressful conditions - and to be replaced by xerophilous grasslands. Following the cessation of coppicing and with stand ageing, the stumps transformation into more homogeneous stand structures have been increasing. Within the context of protected areas the restoration of coppice selection system (with appropriate adaptations) could help to maintain the traditional forest landscape, acting as a silvicultural technique with low environmental and landscape impact.
Keywords
Betula Aetnensis, Mount Etna, Vegetation Dynamics, Coppice Selection System, Forest Restoration
Introduzione
Betula aetnensis Rafin. è una specie endemica a carattere relitto del Monte Etna, che caratterizza il paesaggio della fascia montana superiore ([50], [51], [62], [53]). Si tratta di una entità affine a Betula pendula Roth. che, come attestato da diversi caratteri morfologici ([43], [47]), istologici ([5]) ed ecofisiologici ([35]), denota una tendenza adattativa a condizioni ambientali di tipo montano-mediterraneo. Sotto il profilo tassonomico, si confrontano posizioni differenti: alcuni autori (Rafinesque-Schulmatz 1810 in [61], [47]) considerano la betulla dell’Etna una specie autonoma, altri come sottospecie o varietà di B. pendula (Gussone 1844 in [2], [43], [40]), altri ancora la fanno rientrare nella variabilità di B. pendula ([67]). Betula aetnesis entra nella costituzione di formazioni forestali di tipo temperato che ebbero, soprattutto durante il Quaternario, un ampio sviluppo anche nell’area mediterranea. Sul vulcano B. aetnesis non ha una distribuzione uniforme: in proposito Scuderi ([61]) riferisce che la betulla “vegeta nelle parti alte e settentrionali de’ fianchi del nostro vulcano”. Poli ([50], [51]), Poli Marchese ([52]), Poli Marchese & Patti ([53]) evidenziano come la betulla vegeti soprattutto sul versante di nord-est, e in minor misura su quelli nord e ovest a quote di (850) 1300-1900 m (2110). I boschi di betulla dell’Etna in passato erano sottoposti a sistematiche utilizzazioni; già Scuderi ([61]) affermava che i contadini usavano la betulla per cerchi per botti e per carbone ma non dava indicazioni selvicolturali. Sull’Etna la betulla è stata governata a ceduo. Santocono ([60]) parla di ceduo con rilascio di matricine (50-60 ad ettaro), Agostini ([2]) afferma che il soprassuolo “era governato a ceduo con riserva di matricine”. Secondo Fichera (2005, in verbis) invece si applicava il taglio a sterzo con un periodo di curazione di 12 anni, un diametro di recidibilità di 12 cm e con un basso numero di matricine (poche decine ad ettaro). Da questi cedui si ricavava carbone vegetale (apprezzato per gli antichi ferri da stiro in quanto non dava luogo ad emissione di scintille) e in minor misura legname da opera ([57]). Con l’istituzione del Parco Regionale dell’Etna nel 1987, non sono stati più eseguiti interventi selvicolturali (Fig. 1). Scopo del presente lavoro è quello di valutare le dinamiche evolutive dei popolamenti di betulla in seguito all’interruzione delle attività selvicolturali.
Fig. 1 - Ceduo di betulla invecchiato.

Materiali e Metodi
Descrizione dell’area di studio
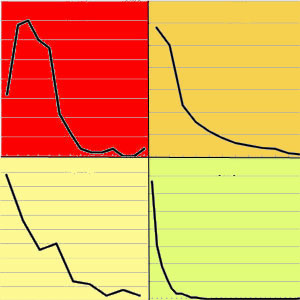
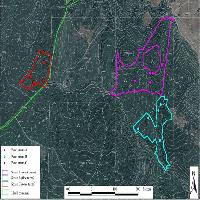
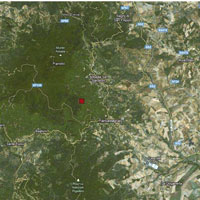
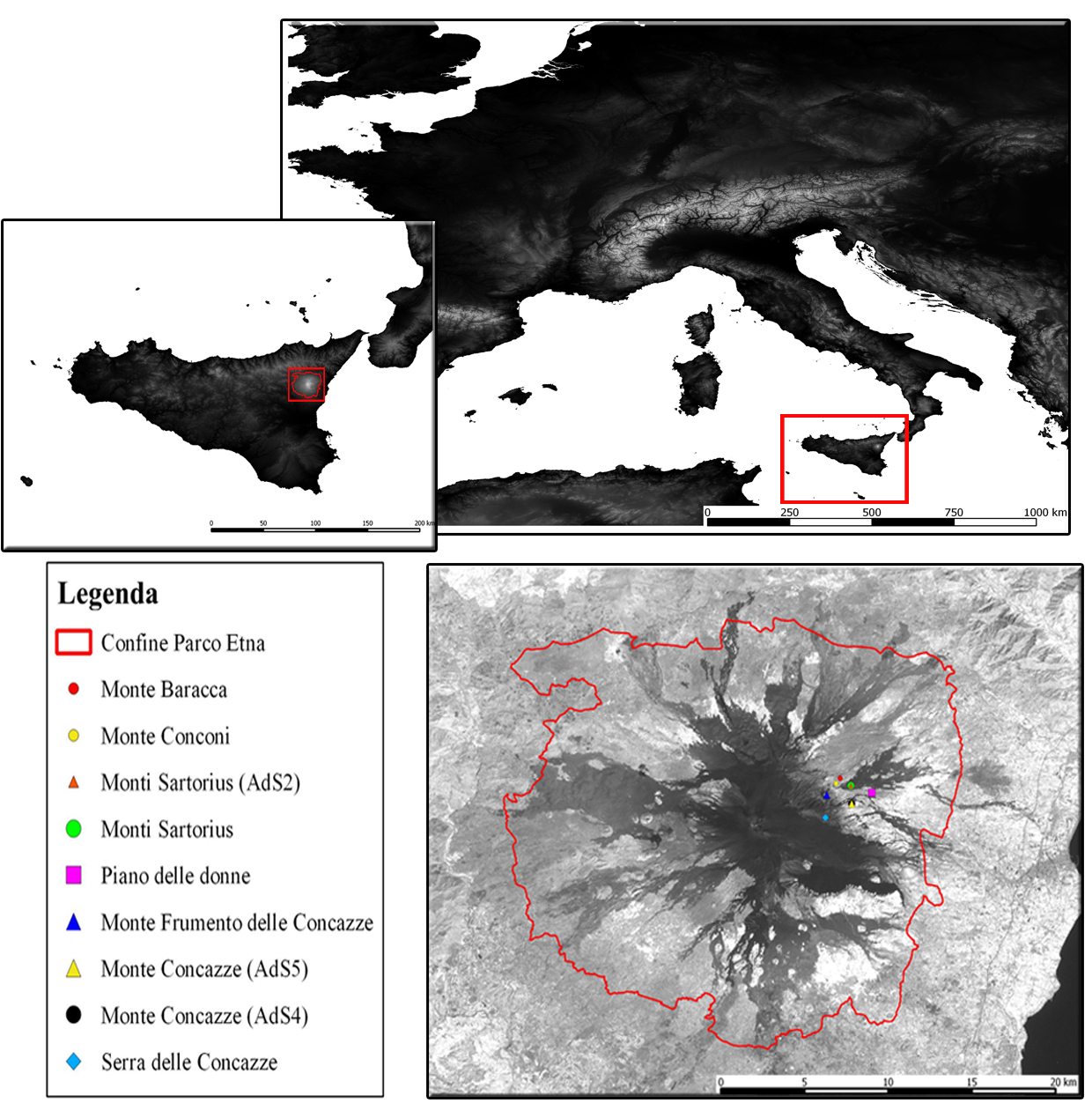
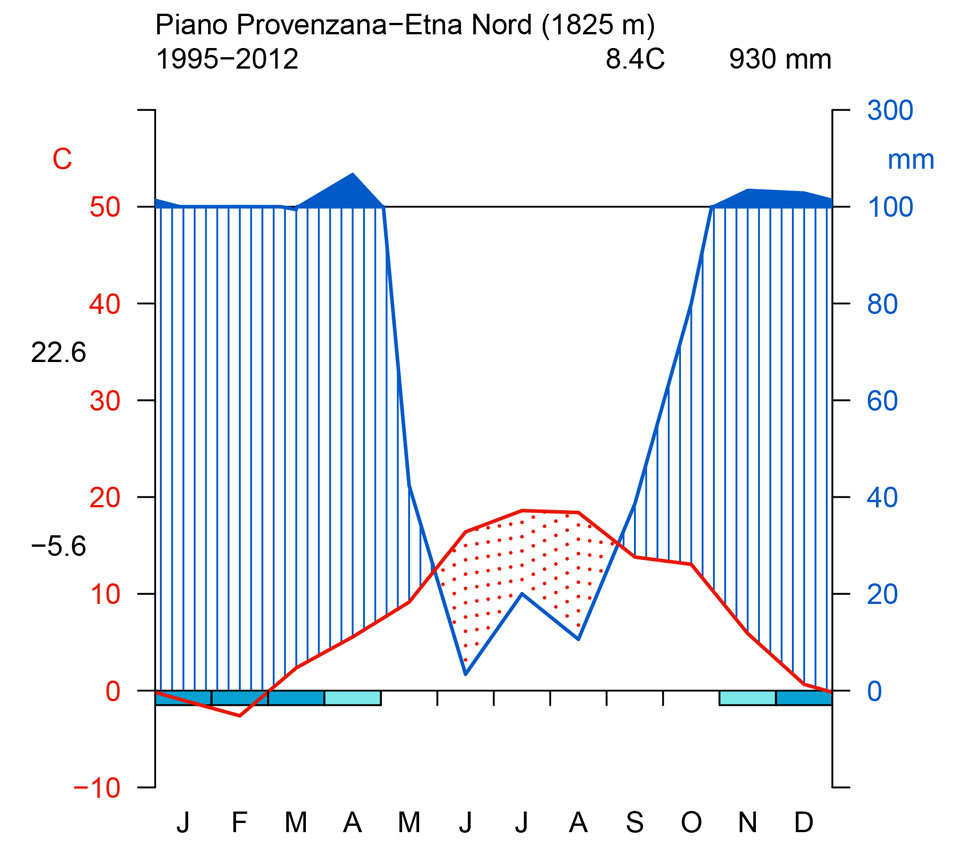
L’area oggetto di studio è situata sul versante orientale del Massiccio dell’Etna (Sicilia), alla latitudine 37° 44’ 59.47” N e longitudine 14 °59’ 44.23” E (Fig. 2). In particolare i betuleti studiati sono ubicati ad altitudini comprese fra 1530 m e 2100 m s.l.m. L’inquadramento tipologico e il confronto con altri sistemi di classificazione è riportato in Tab. 1. Per la caratterizzazione climatica si è fatto riferimento ai dati termopluviometrici relativi alla stazione di “Piano Provenzana - Etna Nord” (1825 m s.l.m.), l’unica che sull’Etna si trovi a quote paragonabili con quelle dell’area di studio. La temperatura media annua è di 8.4 °C e la precipitazione media annua è di 1008 mm. Le precipitazioni nevose interessano il periodo dicembre-marzo ed il periodo di aridità estivo si protrae per circa due mesi (Fig. 3). I betuleti rientrano, secondo la classificazione bioclimatica di Rivas Martinez ([55], [56]), nel bioclima supramediterraneo umido superiore ed in minor misura in quello oromediterraneo umido superiore ([8]). Il substrato geologico è costituito da colate laviche di natura basaltica e prodotti vulcanoclastici dell’Olocene e del Pleistocene/Olocene ([58]). I suoli dei betuleti sono ascrivibili all’associazione Roccia affiorante-Litosuoli e a quella dei Regosuoli-Litosuoli-Suoli Bruni Andici ([23], [24]). Si tratta di suoli pedologicamente poco evoluti con profilo di tipo A-C, tessitura variabile da franco-sabbiosa a sabbiosa, reazione subacida, permeabilità medio-alta, bassa capacità di ritenuta idrica, capacità di scambio cationico (CSC) medio-bassa, notevole grado di fertilità chimica (soprattutto potassio e fosforo), fatta eccezione per l’azoto, che risulta carente, e per il basso tenore di sostanza organica ([24], [35]). Questa area vulcanica si caratterizza per due peculiarità. La prima riguarda i frequenti apporti di ceneri e scorie che arricchiscono il suolo di elementi minerali. La seconda interessa le relazioni atmosfera-vegetazione: da una parte le concentrazioni, talvolta elevate, di zolfo ([36]) che deprimono la vegetazione, dall’altra quelle di anidride carbonica, che stimolano invece l’accrescimento delle piante ([11], [12], [34], [3]).
Fig. 2 - Localizzazione dell’area di studio. Monte Etna, immagine ripresa dal satellite Landsat 7. (Ads): area di saggio.
Tab. 1 - Tipo forestale e confronto tra i diversi sistemi di classificazione ([21], [22], [10]).
| Classificazione | Voce | Valore |
|---|---|---|
| Tipologia | - | Boschi di Betulla |
| EEA | Codice | 6.13.4 |
| Nome tipo | Other birch forest | |
| Corine Biotopes | Codice | 41.B6 |
| Nome unità | Mount Etna birch stand | |
| Tipi della Regione Siciliana | Codice | BS 10X |
| Nome Habitat | Betuleto a Betula aetnensis |
Rilievi dendrometrici e strutturali
I rilievi dendrometrici e strutturali sono stati eseguiti in 9 aree di saggio (AdS) di 20 x 40 m e solo in due casi di 20 x 30 m e 20 x 50 m (Tab. 2). Per ogni area di saggio è stato rilevato il diametro a 1.30 m (delle sole piante con diametro > 1 cm) e l’altezza dendrometrica degli individui. Per determinare l’etàè stato effettuato il prelievo di carotine mediante succhiello di Pressler con trivellazione eseguita a 1.30 m da terra in modo perpendicolare all’asse della pianta in senso radiale, avendo cura di disporsi da monte verso valle. Le carotine sono state scarificate e levigate con l’utilizzo di carta abrasiva in maniera tale da poter meglio evidenziare gli anelli. La lettura delle singole carotine è stata eseguita con un dendrocronografo LINTAB Rinntech, con una precisione di 0.01 mm, interfacciato ad un PC e gestito da uno specifico software (TSAP: Time Series Analysis and Presentation). La struttura si è ottenuta attraverso la realizzazione di un transect, tracciato nella parte mediana dell’area di saggio, largo 10 m. Di ogni pianta che ricadeva all’interno del transect, sono stati rilevati: la posizione topografica, il diametro a 1.30 m, l’altezza dendrometrica, l’altezza di inserzione della chioma (in corrispondenza del primo ramo verde), i 4 raggi della chioma misurati ortogonalmente (secondo le direzioni N-S ed E-W). Si è calcolato successivamente il grado di copertura e si è provveduto alla rappresentazione grafica con software SVS 3.31 (Stand Visualization System) della Pacific Northwest Research Station, USDA Forest Service, USA.
Tab. 2 - Elementi dendrometrico strutturali. (SDT): necromassa in piedi; (CWD): necromassa a terra; (*): i dati relativi a diametro e altezza riportano, rispettivamente, i valori medi e, fra parentesi, i minimi e i massimi. Betuleto tipico: (AdS 1) Monte Frumento delle Concazze; (AdS 2) Monti Sartorius; (AdS 4) Monte Concazza; (AdS 8) Monti Conconi; (AdS 9) Serra delle Concazze; (AdS 6) Piano delle Donne. Betuleto misto: (AdS 3) Monte Sartorius; (AdS 7) Monte Baracca; (AdS 5) Monte Concazza.
| Parametro | Betuleto tipico | Betuleto misto | |||||||
|---|---|---|---|---|---|---|---|---|---|
| AdS 1 | AdS 2 | AdS 4 | AdS 8 | AdS 9 | AdS 6 | AdS 3 | AdS 7 | AdS 5 | |
| Quota (m slm) | 1760 | 1700 | 1760 | 1740 | 2015 | 1560 | 1710 | 1725 | 1770 |
| Esposizione | E | NO | N | NE | NE | E | NE | E | NE |
| Inclinazione (%) | 20 | 20 | 15 | 15 | 30 | 5 | 15 | 15 | 25 |
| Età media (anni) | 43 | 47 | 37 | 32 | 55 | 42 | 40 | 25 | 51 |
| N ceppaie | 312 | 325 | 550 | 150 | 150 | 360 | 475 | 150 | 537 |
| N polloni/ceppaia | 5.5 | 6.1 | 9.1 | 6.5 | 43.3 | 4.2 | 4.2 | 6.5 | 5.2 |
| N polloni | 1787 | 2000 | 5016 | 975 | 6500 | 1510 | 1987 (+ 9% pino calabro) |
975 (+ 49% pino calabro) |
2787 (+30%faggio) |
| Grado di copertura (%) | 90 | 95 | 85 | 75 | 40 | 80 | 95 | 70 | 80 |
| Diametro* (cm) | 15.9 (1-32) |
12.2 (1-29) |
6.6 (1-18) |
9.8 (1-30) |
3.9 (1-18) |
10.5 (4-28) |
11.3 (1-28 ) |
9.1 (1-22) |
9.5 (1-27) |
| Altezza* (m) | 11.6 (1.8-15.5) |
10.6 (1.6-14.6) |
4.9 (1.6-8.2) |
8.8 (1.5-15.2) |
2.3 | 8.6 (3.2-14.1) |
9.4 (1.6-14.3) |
7.1 (1.8-12.4) |
7.4 (1.7-13.1) |
| Area basimetrica (m2/ha-1) | 35.8 | 23.5 | 14.1 | 16.7 | 7.5 | 13 | 19.9 | 6.5 | 19.9 |
| SDT (m3/ha-1) | 13.3 | 8.9 | 1.6 | 8.7 | 0.2 | 3.2 | 11.3 | 22.1 | 9 |
| Alberi morti in piedi (%) | 15.3 | 33.3 | 13 | 12.2 | 2 | 3 | 26 | 46 | 21 |
| CWD (m3 ha-1) | 3.4 | 8.4 | 2.7 | 5.7 | 0 | 0 | 4.3 | 8.8 | 12.6 |
Analisi della rinnovazione
E’ stata considerata rinnovazione naturale l’insieme delle piante arboree nate da seme con diametro < 1 cm a 1.30 m da terra. Il rilievo è stato eseguito mediante un campionamento sistematico di 3 subaree circolari di 1 m2 ogni 10 m, lungo 4 allineamenti. In ogni subarea è stato rilevato il numero dei semenzali; per ogni individuo sono state rilevate: la specie, l’altezza (espressa in cm), l’età (stimata). Inoltre le singole subaree sono state classificate in base all’ubicazione: coperto (c), scoperto (s) e margine (m).
Rilievi della necromassa
La necromassa è utilizzata come indicatore per la valutare la biodiversità e la naturalità di un sistema forestale ([38], [20], [30]). E’ stata distinta in: necromassa in piedi, comprendente le piante morte in piedi, intere o troncate, e necromassa a terra, comprendente tutti i rametti, rami e fusti di alberi e arbusti morti che sono caduti e che si trovano sul terreno ([31], [7]). Nel primo caso, sono stati misurati il diametro e l’altezza in tutti i soggetti presenti nell’area di saggio con diametro >2.5 cm a 1.30 m da terra. Nel secondo caso, è stato impiegato un campionamento per intersezione lineare ([17]). Di ogni singolo pezzo è stata rilevata la lunghezza, mentre è stato misurato il diametro dei soli pezzi con diametro >2.5 cm che intersecavano il segmento. Le elaborazioni hanno riguardato: (a) per la necromassa in piedi, il numero dei fusti morti in piedi, la percentuale di piante morte sul totale, la ripartizione in classi di diametro, il volume con la formula di Huber; (b) per la necromassa a terra, il volume di ogni segmento lineare, impiegando la seguente formula (eqn. 1):
dove n è il numero dei pezzi intersecati, d è il diametro dei pezzi (cm) nel punto di inserzione con il segmento e L è la lunghezza del segmento lineare campione (m).
Per il volume complessivo ad ettaro si è applicata la seguente formula (eqn. 2):
dove n è il numero dei segmenti campione, Vhaj è la massa ad ettaro dei residui legnosi stimata sul j-esimo segmento campione e Lj è la lunghezza del j-esimo segmento lineare campione.
Analisi fitosociologica
I rilievi sulla vegetazione sono stati eseguiti con il metodo di Braun-Blanquet ([6]), articolato nelle seguenti fasi: (i) individuazione della stratificazione secondo il seguente schema: strato arboreo superiore (A1, con altezza maggiore di 15 m), strato arboreo inferiore (A2, con altezza compresa tra 6 e 15 m), strato arbustivo superiore (a1, con altezza compresa tra 2 e 5 m), strato arbustivo inferiore (a2, con altezza compresa tra 1 e 2 m), strato delle nanofanerofite e camefite (a3, con altezza fino a 1 m), strato erbaceo (E); (ii) inventario completo delle specie presenti in ciascuno strato; (iii) stima ad occhio della superficie coperta dalle vegetazione e da ciascuna singola specie in ognuno degli strati considerati. La stima è stata effettuata secondo una scala convenzionale (+: copertura molto scarsa; 1: <5%; 2: <5-25%; 3: 25-50%; 4: 50-75%; 5: copertura > 75%). La nomenclatura delle specie segue Conti et al. ([15]) e Giardina et al. ([26]). L’insieme dei rilievi fitosociologici è stato sintetizzato in una tabella nella quale vengono poste sulle righe le diverse specie e sulle colonne i rilievi fitosociologici. Si è ottenuta così una tabella fitosociologica grezza che contiene, nell’intersezione tra righe e colonne, l’indice di copertura-abbondanza relativo a quella particolare specie (riga) e a quel particolare rilievo (colonna). Le tabelle fitosociologiche non strutturate costituiscono delle matrici elaborabili con i metodi dell’analisi statistica multivariata. Il primo passo del processo elaborativo è stato quello di togliere da ciascuna tabella tutte le specie sporadiche, cioè quelle che compaiono in un solo rilievo. Successivamente gli indici di copertura sono stati trasformati, secondo il metodo proposto da Van der Maarel ([65]), in modo da renderli compatibili con l’utilizzo dell’elaboratore elettronico secondo la seguente scala convenzionale: r: 1; +: 2; 1: 3; 2: 5; 3: 7; 4: 8; 5: 9.
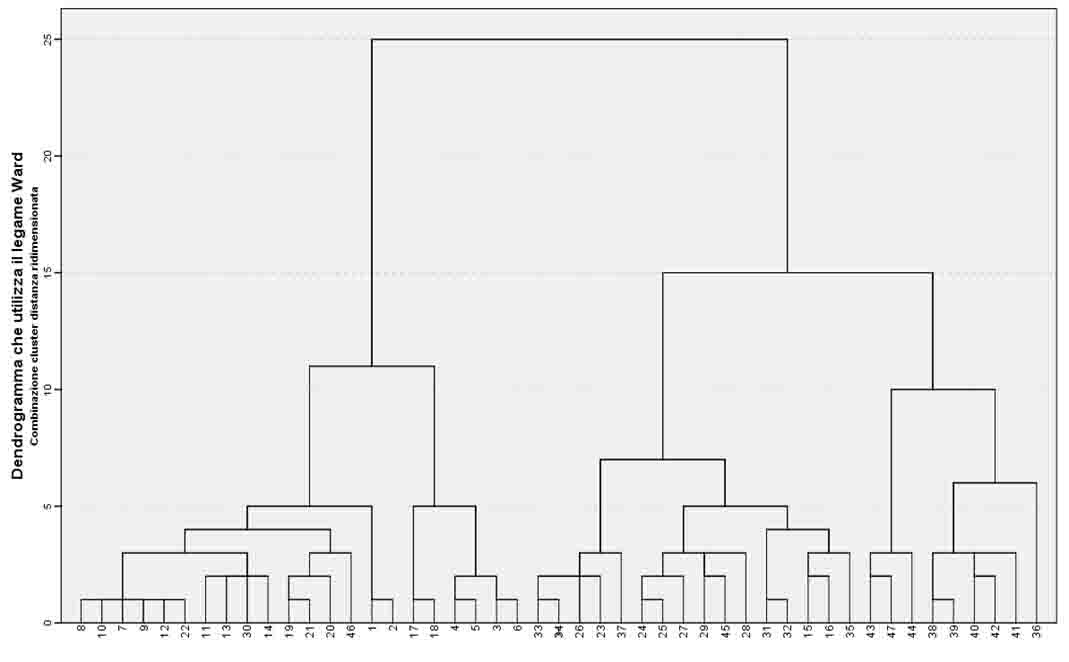
Le tabelle così definite sono state elaborate mediante il software IBM SPSS versione 20.0. Per meglio comprendere i rapporti tra i rilievi è stata fatta un’analisi di correlazione bivariata (correlazione di Pearson a 2-code) per verificarne la significatività. Successivamente, per giungere ad un raggruppamento dei rilievi in base alla similitudine nella composizione in specie, si è utilizzata la cluster analysis. Con questa tecnica, attraverso l’uso di algoritmi basati su funzioni di distanza e similarità, i rilievi sono stati rielaborati e riclassificati. La matrice di distanza è stata calcolata attraverso l’algoritmo della distanza euclidea, mentre per la regola di fusione dei gruppi è stato adottato il metodo di Ward ([68]) che realizza una classificazione gerarchica minimizzando la varianza all’interno dei gruppi. Ad ogni stadio, vengono pertanto fusi i gruppi che producono il minimo aumento della varianza totale entro i gruppi. Una partizione si considera tanto migliore quanto più le classi risultano omogenee al loro interno e differenti l’una dall’altra. Il risultato della cluster analysis viene espresso da un dendrogramma di classificazione, che riporta in ascissa il numero d’ordine distintivo di ciascun rilievo ed in ordinata la scala di distanza. Nel dendrogramma si osservano gruppi di rilievi di diverso grado gerarchico, alcuni più grandi che comprendono altri più piccoli, a seconda del livello di distanza considerato.
Risultati
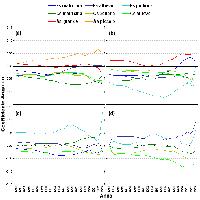
L’analisi multivariata dei rilievi fitosociologici ha permesso di raggruppare i singoli rilievi in più livelli gerarchici (Fig. 4). La classificazione di tutti i rilievi ha evidenziato due cluster principali definiti in termini fitosociologici e floristico-vegetazionali (Tab. S1 in Appendice 1, Tab. S2 in Appendice 2), che si analizzano separatamente.
Fig. 4 - Dendrogramma risultato dalla cluster analysis che mostra la disposizione ed il rapporto tra i rilievi fitosociologici. I numeri su l’asse x indicano il numero del rilievo fitosociologico.
Cluster 1: Betuleto della fascia altomontana
Tale cluster risulta costituito dai seguenti rilievi: 8, 10, 7, 9, 12, 22, 11, 13, 30, 14, 19, 21, 20, 46, 1, 2, 17, 18, 4, 5, 3, e 6. La categoria “betuleto della fascia altomontana” è rappresentata da popolamenti pressoché monospecifici di Betula aetnensis Rafin della fascia bioclimatica oromediterranea (Irano-Nevadense sensu [46]), situati a quote superiori ai 1850-1900 m s.l.m. circa, in corrispondenza dell’area Monte Zappinazzo-Vallone delle Quarantore e di quella di Serra delle Concazze. Si tratta di popolamenti dal portamento prevalentemente arbustivo caratterizzati principalmente dalla assenza di altre specie arboree (rari individui di pino calabro di ridotta statura sono presenti nella sua parte inferiore) e dalla estrema povertà floristica. Le specie degli strati nanofanerofitico-camefitico ed erbaceo (a3, E) sono principalmente appartenenti al contingente del Rumici-Astragaletea siculi ([9]). All’interno di questa tipologia è possibile riscontrare la presenza di Berberis aetnensis e Juniperus communis subsp. hemisphaerica (rilievi 1 e 2), con una fisionomia basso arboreo o alto-arbustiva, con elevata copertura, presenza nello strato a2 e a3 di Berberis aetnensis e Juniperus communis subsp. hemisphaerica, Cerastium tomentosum var. aetnaeum, maggiore presenza delle emicriptofite dell’Astragaletum siculi, quali Bellardiochloa variegata subsp. aetnensis, Tanacetum siculum ed una diminuzione di Astragalus siculus e Festuca circummediterranea. L’AdS 9 è rappresentativa dal punto di vista dendrometrico e strutturale di questo tipo di popolamenti. Essi hanno un basso grado di copertura e presentano un elevato numero di polloni di piccole dimensioni e di modesto sviluppo in altezza, espressione dei limiti stazionali in cui vegeta la betulla (Tab. 2). Non vi è presenza di rinnovazione naturale (Tab. 3).
Tab. 3 - Rinnovazione naturale. (AdS): area di saggio; (c): coperto; (s): scoperto; (m): margine. Betuleto tipico: (AdS 1) Monte Frumento delle Concazze; (AdS 2) Monti Sartorius; (AdS 4) Monte Concazza; (AdS 8) Monti Conconi; (AdS 9) Serra delle Concazze; (AdS 6) Piano delle Donne. Betuleto misto: (AdS 3) Monte Sartorius; (AdS 7) Monte Baracca; (AdS 5) Monte Concazza.
| Parametro | Betuleto tipico | Betuleto misto | |||||||
|---|---|---|---|---|---|---|---|---|---|
| AdS 1 | AdS 2 | AdS 4 | AdS 8 | AdS 9 | AdS 6 | AdS 3 | AdS 7 | AdS 5 | |
| Specie | - | pino calabro(c) | - | Pinocalabro(c) | - | querce (c)querce (s)pino calabro (s) | querce (m) | - | - |
| Densità (n/m2) | - | 0.08 | - | 0.08 | - | 2.6 2 1 | 0.16 | - | - |
| Altezza media (cm) | - | 30 | - | 30 | - | 20 13 1 | 5 | - | - |
| Età media (anni) | - | 7 | - | 7 | - | 3 3 14 | 2 | - | - |
Cluster 2: Betuleto della fascia montana
Tale cluster risulta costituito dai seguenti rilievi: 33, 34, 26, 23, 37, 24, 25, 27, 29, 45, 28, 31, 32, 15, 16, 35, 43, 47, 38, 39, 40, 42, 44, 41 e 36). La categoria “betuleto della fascia montana” individua i popolamenti di Betula aetnensis Rafin. della fascia bioclimatica supramediterranea (di tipo subatlantica sensu [46]), situati a quote comprese tra 1530 m e 1850-1900 m s.l.m. Si tratta di popolamenti dal portamento tipicamente arboreo, caratterizzati sia da una più accentuata dinamica evolutiva - diversi tratti del betuleto si presentano interessati da processi successionali, più o meno progrediti, con pino calabro e in minor misura con faggio e querce spp. - sia da una maggiore diversità floristica, avendosi difatti un deciso aumento sia quantitativo che qualitativo delle specie dei Querco-Fagetea ed una corrispondente diminuzione delle specie del contingente floristico del Rumici-Astragaletea siculi. La maggior parte dei rilievi si riferisce a popolamenti monospecifici di betulla della fascia montana superiore, che si presentano caratterizzati da una dominanza pressoché assoluta della specie, valori medi di copertura dello strato arboreo oscillanti da poco più del 50% fino all’80% e comunque spesso superiori al 70%, strato arbustivo con una discreta partecipazione della Genista aetnensis, strato a3 abbastanza eterogeneo con la partecipazione anche cospicua di Juniperus communis subsp. hemisphaerica e di Adenocarpus bivonae, strato erbaceo eterogeneo ma sempre dominato da Festuca circummediterranea. In generale si tratta dei popolamenti di betulla meglio conservati, che presentano una situazione evolutiva meno accentuata essendo scarsamente presenti le altre specie arboree. Le AdS 1, 2, 4, 6 e 8 rappresentano questa tipologia dominata dalla sola betulla (Tab. 2). La rinnovazione è assente o molto bassa, solo nell’AdS 6 diviene più consistente con prevalenza di querce e pino calabro (Tab. 3 - Fig. 5a). Ad un’analisi di maggior dettaglio si nota come la cluster analysis divida il cluster 2 in tre sub-cluster ad un livello più basso di dissimilarità, che si differenziano tra loro da un punto di vista floristico e strutturale. Un primo sub-cluster comprende popolamenti di quota generalmente compresa tra 1600 m e 1700 m s.l.m., caratterizzati da una presenza sporadica di Pinus nigra Arnold subsp. calabrica (Loud.) A.E. Murray nei diversi strati arborei (rilievi 23, 24, 25 e 26), valori medi di copertura elevati e compresi tra l’80% e il 90%, strato arbustivo scarsamente rappresentato, strato a3 quasi assente, strato erbaceo caratterizzato dalla discreta presenza di specie nemorali quali Crepis leontodontoides e Cephalanthera longifolia; ben rappresentate sono però anche le specie dei Rumici-Astragaletea siculi; notevole è la presenza della rinnovazione di pino calabro.
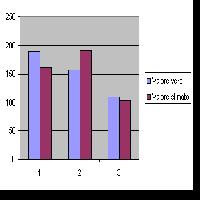
Fig. 5 - Transect di struttura e distribuzione diametrica. Vista prospettica del transect e distribuzione delle piante in classi di diametro con errore standard per ciascuna area di saggio. (a): betuleto puro; (b): betuleto misto; (AdS): area di saggio.
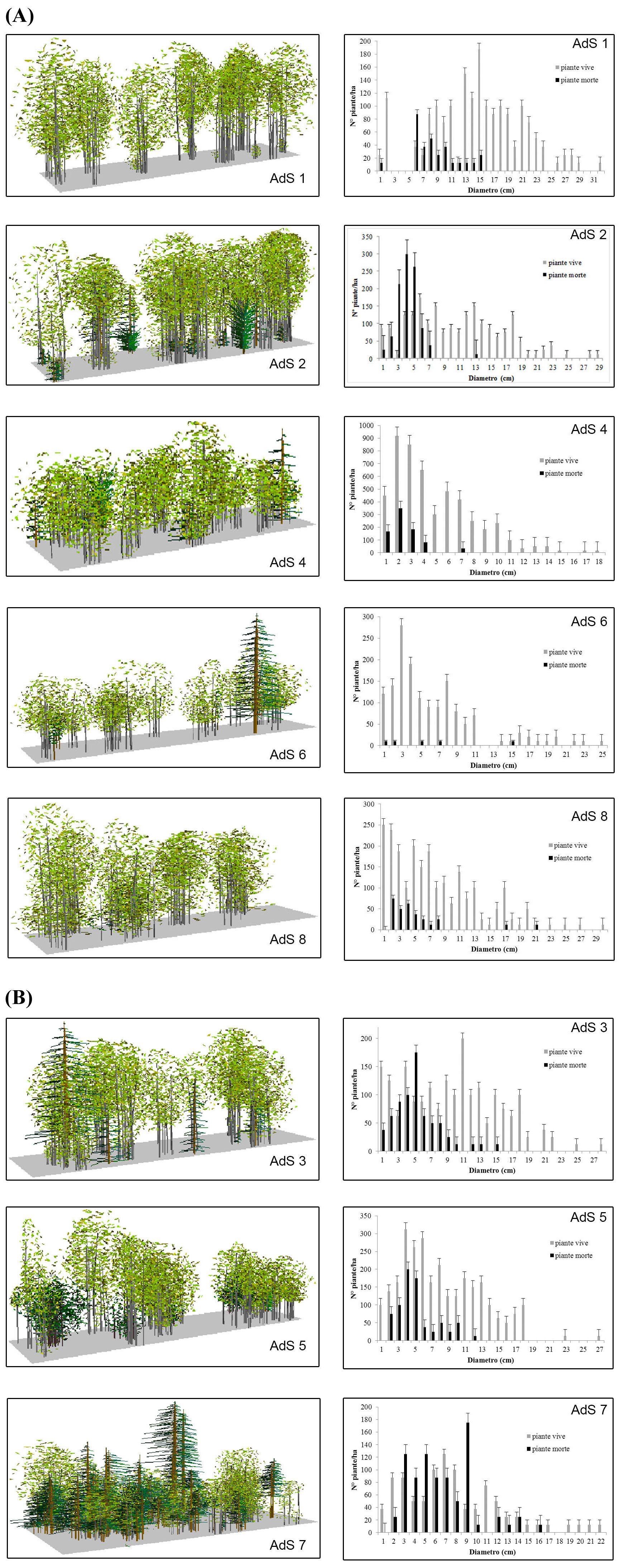
Le AdS 3 e 7 descrivono questi popolamenti dal punto di vista dinamico-strutturale. In essi la presenza del pino calabro varia in numero di individui dal 10 al 50%, segno di una tendenza a sostituire la betulla su suoli meno evoluti (Tab. 2). La rinnovazione naturale è molto scarsa e rappresentata dalle sole querce (Tab. 3). Nella Fig. 5b è riportata la distribuzione in classi diametriche delle piante vive e morte. Un secondo sub-cluster riunisce rilievi (27, 28, 29, 31 e 32) caratterizzati dalla costante presenza del faggio nei diversi strati arborei (si riscontra anche una rara presenza del pino calabro), valori medi di copertura generalmente elevati, che in qualche caso raggiungono valori del 100%, strato arbustivo a1 e a2 poco rappresentati che vede in alcuni casi una discreta presenza della Genista aetnensis, anche lo strato a3 è poco rappresentato con valori medi di copertura compresi tra 15 e 30%, dati da Adenocarpus bivonii, Astragalus siculus e Juniperus hemisphaerica alternativamente dominanti; lo strato erbaceo è anch’esso caratterizzato dalla presenza di specie nemorali.
L’AdS 5 evidenzia una presenza del faggio che si esprime con circa il 30% del numero di individui: con un suolo maggiormente evoluto, la rinnovazione naturale di faggio si è affermata nel betuleto (Tab. 2) e si è evoluta in individui più adulti. Il fenomeno evolutivo è evidente nonostante risulti assente la rinnovazione naturale (Tab. 3).
Questi popolamenti tendono a evolvere in direzione di popolamenti misti, con una importante partecipazione del faggio e in minore misura di pino calabro. Questo trova riscontro nell’analisi dei rilievi del terzo sub-cluster, appartenenti per lo più a popolamenti di tipo misto (rilievi 33, 34, 37, 45, 35, 43, 47, 38, 39, 40, 42, 44, 41, 36) dove la betulla si mescola con il pino calabro a quote generalmente comprese tra 1600 - 1700 m s.l.m., con il faggio a quote superiori a 1700 m s.l.m. e con querce (in particolare Q.congesta C.Presl) intorno ai 1600 m s.l.m., localizzate in Contrada Cerrita.
Un aspetto di non secondaria importanza è lo stato fitosanitario dei popolamenti di betulla dell’Etna. Si registra una diffusa morìa che interessa la betulla etnea da circa 10 anni, dovuta ad attacchi di funghi patogeni del genere Armillaria e Heterobasidion ([63]). L’attacco è particolarmente aggressivo sulle piante di betulla che vivono in condizioni di stress vegetativo. È stato inoltre evidenziato come la moria di Betula aetnensis si manifesti quando è associata a pino calabro ([64]), probabilmente per una trasmissione del fungo dall’una all’altra specie.
Discussione
Le dinamiche evolutive: età, densità e distribuzione dei diametri dei polloni
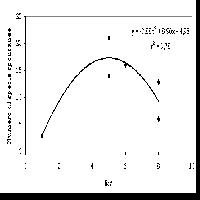
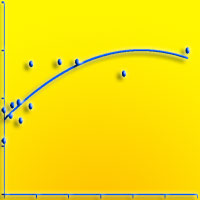
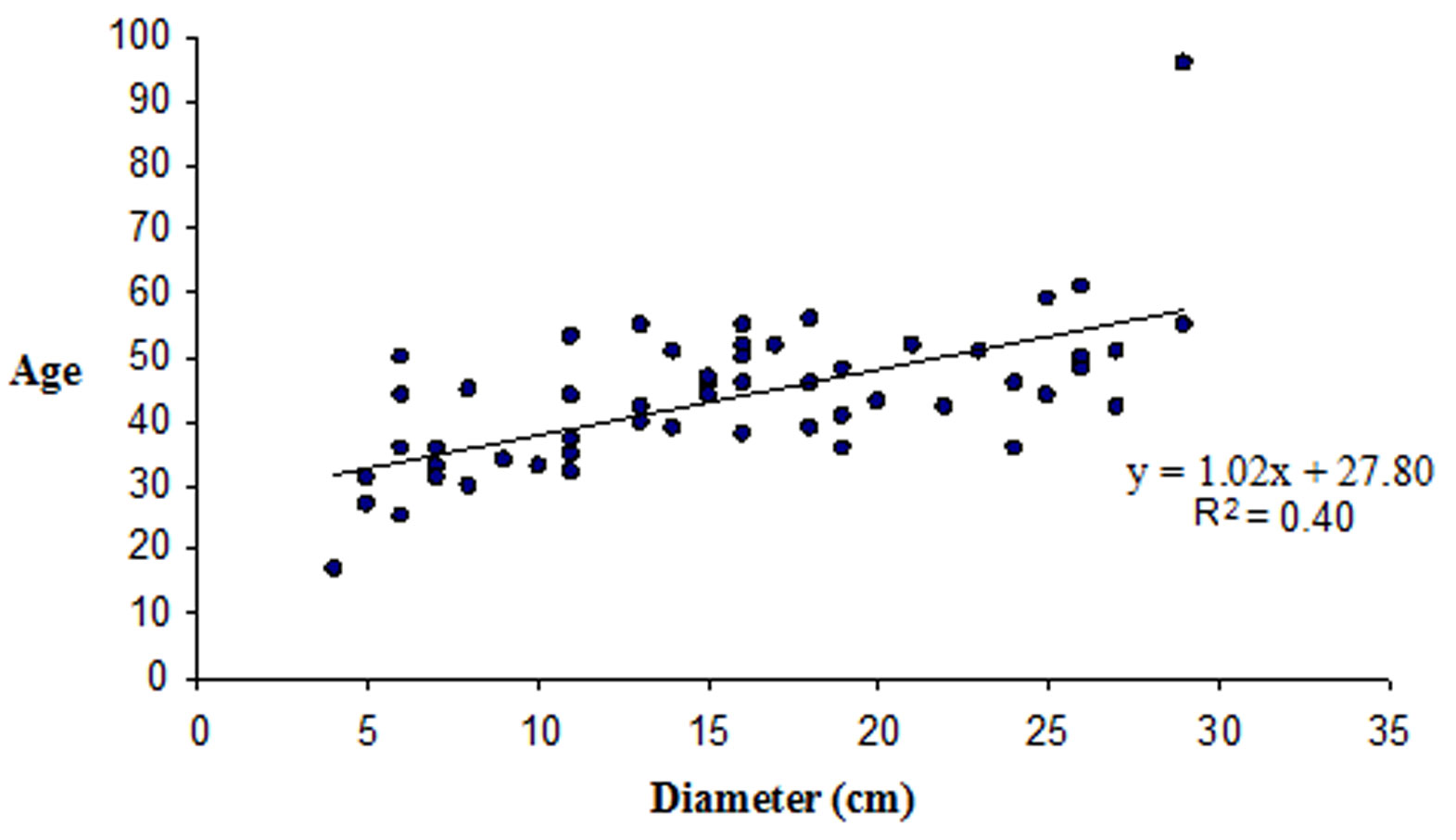
L’età media dei polloni è di 41 anni (42.6 anni per i betuleti puri e 38.6 per i misti); le età più elevate si ritrovano nell’AdS 9 con 67 e 71 anni e nell’AdS 1 in una matricina di 96 anni. La frequenza maggiore si riscontra nella classe 45-50 ma in generale le piante analizzate hanno un’età compresa tra i 35 e i 55 anni. La relazione diametro-età (Fig. 6) non è molto marcata (R2 = 0.40); infatti, ad esempio, i soggetti di 40 o 50 anni possono avere sia 5 che 25 cm di diametro. Il numero di polloni per ceppaia è nei popolamenti puri di 4.2-9.1 (43.3 nell’AdS 9) e di 4.2-6.5 in quelli misti: si tratta di valori bassi che confermano la peculiarità della specie di non riuscire a sviluppare un numero elevato di polloni. Nei betuleti puri, in seguito alla sospensione delle ceduazioni, è possibile valutare meglio le modalità di riorganizzazione strutturale dei polloni: nelle AdS 4 e 8 permane una distribuzione delle classi diametriche di tipo esponenziale, espressione di un passato trattamento a sterzo; nell’AdS 2 si ha una distribuzione coetaneiforme con asimmetria a sinistra così come nell’AdS 1. In questi due casi, che corrispondono ai popolamenti più adulti, è più evidente il processo evolutivo in seguito alla sospensione del trattamento a sterzo. L’AdS 6 non è significativa in quanto sono stati eseguiti sfollamenti qualche anno fa. Nelle AdS di età maggiore i polloni adulti occupano lo strato superiore e ombreggiano i polloni giovani dello strato sottostante, che muoiono come si nota anche dai valori di necromassa (Tab. 2).
Fig. 6 - Distribuzione delle età in funzione del diametro ad esclusione dell’area di saggio 9.
I processi di rinnovazione naturale della betulla
Nei popolamenti puri e misti vi è assenza di rinnovazione da seme di betulla. La fruttificazione avviene nella betulla in giovane età, a 10-15 anni sui polloni e a 15-25 sulle piante da seme, in modo abbondante con cadenza annuale o biennale. L’età media dei betuleti studiati è di 41 anni. Si tratta di una fase ancora relativamente giovanile, se si considera che la betulla raggiunge i 100-120 anni di età ([18]). Il seme matura a fine estate-autunno quando comincia la disseminazione anemocora che può raggiungere i 192 m dalla pianta madre ([44]). Ne consegue che nella betulla il potenziale di produzione di seme è alto. La mancanza di rinnovazione di betulla deve essere ricercata in altre cause:
- bassa facoltà germinativa (20-30 % per [49], 40% per [42]) e bassa vitalità del seme, che decresce rapidamente: 6% dopo il primo anno, 3% al secondo e 1% al terzo anno ([42]);
- bassa sopravvivenza dei semenzali durante il primo anno, inferiore al 5-10% ([33]);
- assenza di micorrize ([19], [32]), per quanto Napoli ([41]) segnali proprio nei betuleti dell’Etna la presenza di due micorrize (Leccinum scabrum, Russula nitida);
- alto grado di copertura dei popolamenti (>80%), che impedisce la rinnovazione sotto copertura per una specie molto esigente di luce in fase giovanile ([45], [48]);
- elevata erosione idrica ed eolica del seme;
- carenza di N nel suolo ([44]);
- sensibilità allelopatica e alla competizione erbacea ([14]).
Situazioni dinamiche
Nella zona alto-montana la composizione monospecifica e l’assenza di fenomeni evolutivi in atto (senza spostamenti in senso altitudinale) sembrano avvalorare l’ipotesi di una apparente stabilità del betuleto. Ciò conferma le tesi di Gamisans ([25]) e Spada et al. ([62]) secondo le quali i popolamenti residui presenti nell’area mediterranea (Penisola Iberica, Appennino, Corsica, Turchia), dove Betula pendula Roth. è relegata in ambienti più umidi, con uno spostamento verso quote sempre più alte, tendono alla stabilità. Del Favero ([18]) aggiunge che si tratta di formazioni durevoli in quanto bloccate nell’evoluzione dalla povertà del suolo. Betula pendula Roth. è capace di adattarsi a suoli molto acidi, con valori di pH fino a 3.3 ([13]), non tollerate da altre specie come il pino calabro. In questo senso, Betula pendula risulta avvantaggiata dall’assenza di competitori. Le caratteristiche istologiche e anatomiche dell’apparato conduttore della betulla consentono, inoltre, un eccellente adattamento alle condizioni di siccità estiva ([5], [35]) così come l’apparato radicale molto profondo ([4]). Si aggiunga che nella fascia montana la permanenza della betulla può essere stata favorita anche dal taglio a sterzo. Giordano ([27]) precisa che nel caso del taglio a raso della ceppaia la rinnovazione agamica si mantiene elevata fino a 15 anni, anche se dopo 3 ceduazioni la ceppaia si spossa. La facoltà pollonifera declina dopo i 40-60 anni. Quindi sembra importante sottolineare che la modalità di ceduazione a sterzo ha un’azione meno stressante per la ceppaia e consente il rinfoltimento del soprassuolo attraverso la facilitazione dell’emissione di polloni radicali. La betulla dell’Etna si potrebbe conservare anche grazie a fenomeni di rinnovazione agamica naturali, confermando quanto osservato da Spada et al. ([62]), alle quote più alte in popolamenti appenninici. Anche i betuleti relativamente più stabili della fascia montana, soprattutto in assenza di disturbi esterni, vengono nel lungo periodo sostituiti da altre specie ([18]). La lettiera migliora le condizioni del suolo consentendo la formazione di humus di tipo mull ([28]) e quindi l’affermazione di specie più esigenti ([14]). Ciò è evidente nei betuleti misti (AdS 3, 5 e 7) dove si sono avviati da tempo fenomeni evolutivi risalenti presumibilmente all’ultima ceduazione. Essi consistono in una prima colonizzazione di pino calabro e faggio, generalmente localizzata all’interno dell’area di insidenza delle ceppaie di betulla, dove si hanno condizioni microclimatiche più favorevoli e dove la copertura leggera favorisce le specie di fasi successionali più mature ([1]). Questi processi dinamici sono messi in evidenza anche da un maggiore quantitativo di necromassa, i cui valori sono pari a 22.7 m3 ha-1 in quelli misti e a 9.1 m3 ha-1 in quelli puri (Tab. 2).
Conclusioni
I betuleti dell’Etna presentano tre diverse dinamiche evolutive:
- Stabilità. È il caso dei betuleti della fascia alto-montana, almeno nel medio periodo, se non si accentuano fenomeni dovuti ai cambiamenti climatici previsti anche in ambiente mediterraneo ([37]); essi costituiscono quindi un edafo-climax.
- Dinamica evolutiva progressiva. I betuleti della fascia montana presentano una dinamica ben più complessa e articolata in funzione delle caratteristiche pedologiche, che in buona parte dei casi è indice di una progressiva rarefazione e di una concomitante sostituzione con popolamenti arborei monospecifici (pino calabro, faggio, querce) o misti (con betulla - Fig. 7, Fig. 8). Nel caso della pineta, sempre a seconda delle caratteristiche pedologiche, è da prevedere una ulteriore evoluzione verso la faggeta.
- Dinamica evolutiva regressiva. I popolamenti puri di betulla (betuleto tipico) tendono - soprattutto a causa dell’azione congiunta di condizioni climatiche estreme per la specie (dovute all’altitudine) ed un substrato povero - ad essere sostituiti da formazioni erbacee o basso arbustive dei Rumici-Astragaletea, soprattutto se si accentuano annate caratterizzate da un andamento climatico particolarmente sfavorevole (elevate temperature, prolungati periodi di siccità, ecc.). Questa dinamica regressiva è probabilmente da mettere in relazione anche con la morìa della betulla dovuta agli attacchi dei funghi patogeni del genere Armillaria e Heterobasidion. A livello strutturale, in seguito alla sospensione delle ceduazioni e con l’invecchiamento, si sono accentuati i fenomeni di trasformazione delle ceppaie, che tendono a una maggiore uniformità (coetaneiformi). Queste indagini confermano che il paesaggio forestale è destinato, anche nell’arco di pochi decenni, a cambiare sia per il diverso uso delle risorse forestali dovute ai cambiamenti economici e sociali, sia per gli effetti dei cambiamenti climatici ([39]).
Fig. 7 - Colonizzazione di pino calabro all’interno di ceppaia di betulla. Monte Baracca, 1730 m s.l.m.

Fig. 8 - Colonizzazione di faggio all’interno di ceppaia di betulla. Contrada Serracozzo, 1760 m s.l.m.

Il ripristino del ceduo a sterzo, secondo gli adattamenti proposti da Coppini & Hermanin ([16]) - basati sul trattamento a sterzo semplificato alternato, in cui la componente dei polloni è costituita da due sole classi cronologiche/dimensionali - potrebbe contribuire a conservare questo paesaggio, ricadente nell’ambito di un’area protetta. Si pensi, in tal senso, che il ceduo di betulla ha un valore dal punto di vista storico e culturale e che il ceduo a sterzo è una tecnica selvicolturale di basso impatto ambientale e paesaggistico ([59], [16]).
References
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar