Impact of wild ungulates on coppices from the Bisenzio Valley (province of Prato, Italy)
Forest@ - Journal of Silviculture and Forest Ecology, Volume 11, Pages 116-124 (2014)
doi: https://doi.org/10.3832/efor1259-011
Published: Jun 09, 2014 - Copyright © 2014 SISEF
Research Articles
Abstract
Similar to other Apennine areas, a considerable increase of wild ungulate density has been observed in the past decades in the Bisenzio valley (province of Prato), leading to a strong impact on coppice regeneration. In some felling areas, the prolonged browsing on young suckers turned the stumps into low and dense bushes, thus threatening the survival of the forest stand and triggering a loss of environmental stability. The aim of this work was to study the impact of deer on sprout regeneration after cutting in mixed coppices felled in the period 2007-2010 and characterized by the dominance of chestnut, hornbeam, Turkey oak, and beech. Some 36 plots were established in four municipal districts of the Prato province (Cantagallo, Vernio, Montemurlo, Vaiano). Damage and browsing intensity on each stump were evaluated using an index that allowed the comparison among different areas and fores stand types. Regardless of the species, over 90% of the sprouts revealed a very strong damage, whereas only 6% did not exhibit any damage. A significant difference in the sprout height was detected between browsed and non-browsed areas. Despite a high incidence of damage in all the studied areas, chestnut showed a greater resilience against browsing. In summary, overgrazing by wild game may determine severe contraints to the evolutionary dynamics of forest stands, frustrating the effects of silvicultural practices. The possible benefits of the implementation of a large-scale integrated wildlife plan, including hunting and forest-evironmental management are discussed.
Keywords
Coppice, Wild Ungulate Damage, Forest Dynamics, Silviculture, Sustainable Management
Premessa
L’eccessivo carico di fauna ungulata costituisce un fattore di disturbo in grado di apportare gravi limitazioni alle dinamiche evolutive dei soprassuoli forestali, al punto da vanificare gli effetti degli interventi selvicolturali ([27], [30]). È altresì dimostrato che la selettività dei danni di origine alimentare può modificare i rapporti di concorrenza e quindi limitare la presenza di alcune specie, mettendo a repentaglio, nel lungo periodo, la possibilità di sopravvivenza di quelle più appetite: possono risultarne cambiamenti nella composizione specifica e nella struttura degli ecosistemi forestali ([25], [17], [2], [32], [6]).
Per quanto riguarda in particolare la provincia di Prato, nella zona montana della parte settentrionale sono presenti consistenti popolazioni di ungulati selvatici, da anni sottoposti ad azioni di contenimento mediante caccia selettiva (capriolo e cervo) o di eradicazione (daino).
Particolarmente meritevole di attenzione è l’effetto che soprattutto il cervo (in ragione della vastità dell’areale, della numerosità della popolazione e delle esigenze alimentari) esercita sulle colture agrarie e forestali. Del danno sulle colture agricole si è oramai ben consapevoli; restano invece da chiarire molti aspetti di quello a carico dei boschi, con particolare riguardo agli effetti, nel breve e lungo periodo, sui processi di rinnovazione.
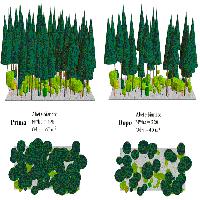
La situazione osservabile nei boschi della Provincia di Prato, ed in particolare nei boschi cedui di specie quercine della fascia boscata che comprende la Val di Bisenzio e la zona appenninica del comune di Montemurlo, indica una pressione molto forte. Boschi cedui tagliati da alcuni anni sono caratterizzati da ceppaie molto brucate che, a causa del morso ripetuto degli ungulati, si presentano come cespugli bassi e densi, dall’incerto avvenire (Fig. 1 e Fig. 2). In certe aree, poi, il livello di danneggiamento è talmente elevato che, qualora l’impatto si prolungasse nel tempo, potrebbe determinare l’innesco di fenomeni regressivi assai pericolosi in chiave di salvaguardia ambientale.
Fig. 1 - Ceppaia di cerro ripetutamente brucata in un ceduo della Val di Bisenzio.

Fig. 2 - Effetti della brucatura del cervo in un ceduo matricinato di cerro della Val di Bisenzio a tre anni dal taglio delle ceppaie.

Con questa ricerca ci si pone l’obiettivo di valutare l’incidenza della fauna ungulata sui processi di rinnovazione agamica nei cedui della Val di Bisenzio. In particolare lo scopo è di individuare i tipi di bosco più suscettibili al danno da brucatura (soprattutto in relazione a età del ceduo e alla specie dominante), sulla base di una metodologia originale speditiva per la valutazione del danno.
Ambiente di studio
Lo studio è stato condotto nella Val di Bisenzio, l’area della Provincia di Prato che segue il corso del fiume Bisenzio compresa tra i rilievi dell’ Appennino Tosco-Emiliano e lo sbocco del fiume nella pianura pratese. Il territorio di studio si estende nei Comuni di Montemurlo, Cantagallo, Vernio, Vaiano per un totale di 192.45 km2 con un’altitudine che varia da 100 a 1300 m s.l.m.
Il clima si presenta diversificato; infatti sui rilievi si riscontra un clima di carattere montano con inverni spesso nevosi, mentre nelle zone più pianeggianti la tendenza è verso una maggiore continentalità. La temperatura media mensile registrata raggiunge valori compresi tra 6 e 1 °C nel mese più freddo, mentre per il mese più caldo si ha una media compresa tra 24 e 17 °C. Scendendo dall’alta Val di Bisenzio il numero di mesi “freddi” (con T media < 10 °C) varia secondo le stazioni: Cottede (zona Montepiano) presenta 5 mesi freddi, Cantagallo 4, Vaiano 3. Le precipitazioni sono influenzate dai rilievi: vengono superati in media 1000 mm annui di pioggia. Di regola le precipitazioni sono più abbondanti in autunno e primavera, mentre il minimo si registra nei mesi estivi. Nelle stazioni di Cottede e Cantagallo non si registrano mesi aridi, mentre a Vaiano è arido il mese di luglio.
Le zone montuose della Val di Bisenzio presentano un’ossatura costituita da rocce sedimentarie e banchi di Arenaria, qua e là intercalati a Galestro; un caso a parte sono i rilievi della Calvana, che presentano rocce carbonatiche (Alberese). Nelle zone dominate dall’Arenaria i terreni hanno reazione tendenzialmente acida e sono più o meno impoveriti di sostanza organica; nelle zone che presentano scisti argillosi si originano terreni profondi e ricchi di humus, a reazione subacida-neutra. I suoli dei rilievi della Calvana, a pH alcalino, sono soggetti al fenomeno del carsismo ipogeo e epigeo, con formazione di grotte e cunicoli sotterranei per tutta la catena montuosa.
Per quanto riguarda la vegetazione, facendo riferimento alla Carta della vegetazione di Arrigoni ([5]), nella zona di studio si trovano le seguenti formazioni forestali: boschi di sclerofille sempreverdi, pinete mediterranee, boschi a dominanza di latifoglie decidue termofile, castagneti, cerrete, ostrieti, faggete.
La vegetazione della Val di Bisenzio è in effetti molto varia: la specie più comune nel territorio è il castagno, presente in boschi cedui e in castagneti da frutto. Il sottobosco è formato in questo caso principalmente da erbe a foglia laminare, esigenti o mediamente esigenti, con presenza di felci e rovi. Si possono però presentare anche castagneti di medio sviluppo a causa della minor fertilità del terreno: il sottobosco è composto in questo caso da calluna, ginestra dei carbonai e felce aquilina.
I querceti della zona sono a prevalenza di cerro: raramente si tratta di boschi puri, in quanto il cerro è misto, secondo i casi, a roverella, castagno, carpino nero, orniello, acero campestre e olmo campestre. Nelle zone a bassa quota, su terreni di natura arenacea, domina la mescolanza con il castagno. Le zone a maggior presenza di carpino nero sono prevalenti nei Monti della Calvana: in questi popolamenti, alla composizione specifica contribuiscono anche acero campestre, ciliegio e nocciolo. La faggeta è presente in molte zone della Val di Bisenzio, principalmente ad alta quota, sebbene siano presenti stazioni eterotopiche in cui il faggio scende fino a 400 m s.l.m.
Per quanto riguarda la presenza del cervo, secondo le stime al bramito effettuate nel corso del 2010 dal Comprensorio A.C.A.T.E.R. (Areale Cervo Appenninico Toscana Emilia Romagna) Centrale sono stati contati in Val di Bisenzio 112 maschi bramitanti su una superficie di 97.18 km2: ne risulta una densità di 1.15 esemplari per km2, che riferita all’areale riproduttivo arriva a un totale di 0.80. Riguardo alla consistenza complessiva della popolazione, per la Val di Bisenzio al 2012 si stima una cifra di 1047 capi ([1]), corrispondente a una densità di circa 10 esemplari per km2.
A decorrere dal 2000 si è cercato di limitare l’espansione del cervo con la caccia di selezione. Negli anni tra il 2008 e il 2011 vi è stato un aumento dei capi di cervo abbattuti, con un picco massimo di 127 capi nella stagione venatoria 2009-2010, soprattutto per quanto riguarda le classi dei piccoli dell’anno. Per quanto riguarda la classe delle femmine adulte, nelle stagioni venatorie 2008-2009 e 2009-2010 esse sono state abbattute in maggior numero. Nella stagione venatoria 2010-2011 si è registrato un aumento dei capi da prelevare ma una diminuzione degli effettivi prelevati, fenomeno dovuto all’aumento del numero sia di piccoli dell’anno, sia delle femmine adulte prelevabili.
Nell’area di studio è presente anche una consistente popolazione di capriolo, che nel 2012 ammontava a 2270 capi, corrispondente a una densità di 23 esemplari per km2.
Materiali e metodi
L’indagine è stata condotta partendo da un campione di 138 boschi cedui - accomunati dal fatto che i loro proprietari ne avevano fatto richiesta di taglio, presupposto per acquisire le informazioni necessarie per la successiva stratificazione del campione - distribuiti nei comuni di Cantagallo, Montemurlo, Vaiano e Vernio, censiti in base alle denunce di taglio avvenute negli anni dal 2007 al 2009. In realtà le dimensioni effettive delle tagliate non coincidono con quelle dichiarate, in quanto di dimensioni inferiori. Il motivo è che i proprietari tagliano la superficie dichiarata, un po’ alla volta, in più anni, magari lasciando in piedi il soprassuolo che, per provvigione, tipologia assortimentale, difficoltà di smacchio, risulta meno conveniente tagliare. Inoltre i confini reali della proprietà in fase di taglio non sono definiti con precisione e, spesso, nemmeno noti ai proprietari stessi: ne consegue che, per non sconfinare, si limita la superficie della tagliata.
Il campione è stato stratificato in funzione di:
- specie prevalente: cerro, carpino nero, castagno, faggio, misto (nel caso in cui non si registri dominanza da parte di alcuna specie).
- superficie utilizzata (A inferiore a 0.5 ha; B fra 0.51 e 1.5 ha, C fra 1.51 e 2.5 ha, D superiore a 2.51 ha).
- epoca di taglio (2007, 2008, 2009, 2010).
È stato quindi determinato un sottocampione di 36 tagliate, in modo casuale e ponderato in relazione alla frequenza in ciascun livello di stratificazione.
In ciascuna tagliata, nel tratto più ampio, è stato realizzato un transect lineare trasversale, lungo le isoipse, di lunghezza variabile in relazione all’ampiezza della tagliata e di larghezza variabile da 4 m per le ceppaie tagliate a 20 m per il censimento delle eventuali matricine.
Di ogni matricina presente all’interno della tagliata sono stati rilevati specie, diametro a 1.30 m (cm), altezza (m).
Per ogni ceppaia sono stati rilevati: specie, diametro maggiore e diametro ad esso ortogonale, numero dei ricacci (suddivisi in brucati e non brucati) e altezza dei 3 ricacci più alti per ceppaia.
Ciascuna ceppaia campionata è stata classificata “interna” alla tagliata se ubicata a una distanza dal margine della tagliata superiore all’altezza media (hm) delle matricine presenti, mentre in caso contrario la ceppaia è stata classificata di “margine”.
L’impatto della brucatura (I) sulle singole ceppaie è stato definito in relazione percentuale tra il numero di ricacci brucati e quello dei ricacci totali.
È stata poi definita una scala per la classificazione di ciascuna ceppaia in relazione al valore di I (Tab. 1).
Tab. 1 - Gradi di intensità di brucatura delle ceppaie e intervallo percentuale per ogni classe.
| Grado di I | Descrizione | Intervallo (%) |
|---|---|---|
| ND | Non Danneggiate | I= 0 |
| L | Brucatura Leggera | 1 < I < 20 |
| M | Brucatura Media | 21 < I < 40 |
| F | Brucatura Forte | 41 < I < 60 |
| MF | Brucatura Molto Forte | I > 61 |
Successivamente è stata classificata ciascuna tagliata in relazione all’impatto delle brucature (IT), tenendo conto della frequenza delle ceppaie per ciascun grado di I (Tab. 2).
Tab. 2 - Classificazione delle tagliate in base all’impatto delle brucature sulle ceppaie.
| Impatto sulla tagliata (IT) | Descrizione | Valori di riferimento di I |
|---|---|---|
| T | Trascurabile | (ND+L) > 80% |
| I | Intenso | (MF+F) > 60% |
| ME | Intermedio | tutti gli altri casi |
Al fine di verificare l’impatto del morso degli ungulati sull’accrescimento in altezza dei polloni di cerro (la specie più diffusa e, allo stesso tempo, quella per cui è disponibile il maggior numero di osservazioni) è stato condotto il test dell’ANOVA rispetto alla variabile “altezza media del pollone dominante”, ponendo come fonte di variazione l’età della tagliata.
Risultati
Impatto della brucatura sulle ceppaie in relazione alla specie
Complessivamente sono state censite 818 ceppaie, di cui 468 di cerro, 170 di castagno, 109 di carpino nero, 45 di orniello, 19 di faggio, 5 di robinia e 2 di carpino bianco.
Il cerro risulta la specie più danneggiata, col 91% delle ceppaie nella classe di danno MF, segue il carpino nero col 62% e il castagno con il 32%. Il 93% delle ceppaie di orniello, che risulta la specie meno danneggiata, rientra invece nella classe ND, mentre per le altre specie il 42 % rientra nella classe ND e il 58% in MF. Nell’insieme il 69% delle ceppaie censite rientra nella classe MF, il 25% nella classe ND, il 3% nella classe M, il 2% nella classe F e l’1% nella classe L (Tab. 3).
Tab. 3 - Impatto della brucatura sulle ceppaie in relazione alla specie.
| Specie | Grado di I (%) | ||||
|---|---|---|---|---|---|
| ND | L | M | F | MF | |
| Carpino nero | 28 | 2 | 5 | 3 | 62 |
| Castagno | 58 | 1 | 5 | 4 | 32 |
| Cerro | 4 | 1 | 2 | 2 | 91 |
| Orniello | 93 | 2 | 0 | 0 | 4 |
| Altro | 42 | 0 | 0 | 0 | 58 |
| Totale | 25 | 1 | 3 | 2 | 69 |
Impatto della brucatura in relazione al tipo di bosco
Il tipo di bosco più diffuso è quello a dominanza di cerro (15 tagliate), seguito da quello a dominanza di castagno (6 tagliate), carpino nero (2 tagliate), faggio (1 tagliata). Il ceduo misto (cerro, carpino nero e castagno) è rappresentato da 12 tagliate. Tutte le tagliate in cui la specie prevalente è il cerro o il carpino nero rientrano nella classe I (massima intensità di danno), quelle a dominanza di castagno per la metà risultano nella classe T (danno trascurabile), mentre i boschi misti per il 50% sono classificati come I, e per il resto sono equamente suddivisi fra le altre due classi. L’unico bosco di faggio analizzato rientra nella classe I (Tab. 4).
Tab. 4 - Impatto della brucatura in relazione al tipo di bosco.
| Tipo di bosco | Grado di IT (%) | ||
|---|---|---|---|
| T | ME | I | |
| Cerreta | 0.0 | 0.0 | 100.0 |
| Ostrieto | 0.0 | 0.0 | 100.0 |
| Castagneto | 50.0 | 16.7 | 33.3 |
| Faggeta | 0.0 | 0.0 | 100.0 |
| misto | 25.0 | 25.0 | 50.0 |
| Totale | 16.7 | 11.1 | 72.2 |
Complessivamente il 72% delle tagliate censite rientra nella classe I, il 17% in T e l’11% in ME.
Impatto della brucatura sulle ceppaie in relazione all’età dei ricacci
Il 79% delle ceppaie con ricacci di un anno rientra nella classe MF, il 15% nella classe ND (Tab. 5). I ricacci di 2 anni risultano leggermente meno danneggiati (65% nella classe MF, 30 % in ND). I valori dei ricacci di 3 anni non si discostano molto da quelli dei ricacci di 2 anni (nella classe MF si sale al 68%, nella ND ci si abbassa al 26%), mentre i valori dei ricacci di 4 anni tendono ad allinearsi con quelli di 1 anno (il 75% dei ricacci rientra nella MF, il 17% in ND).
Tab. 5 - Età dei ricacci e grado di impatto di brucatura (I).
| Età (anni) | Grado di I (%) | ||||
|---|---|---|---|---|---|
| ND | L | M | F | MF | |
| 1 | 15 | 1 | 4 | 1 | 79 |
| 2 | 30 | 1 | 1 | 2 | 65 |
| 3 | 26 | 1 | 3 | 3 | 68 |
| 4 | 17 | 1 | 5 | 3 | 75 |
Impatto della brucatura sull’accrescimento in altezza dei polloni di cerro
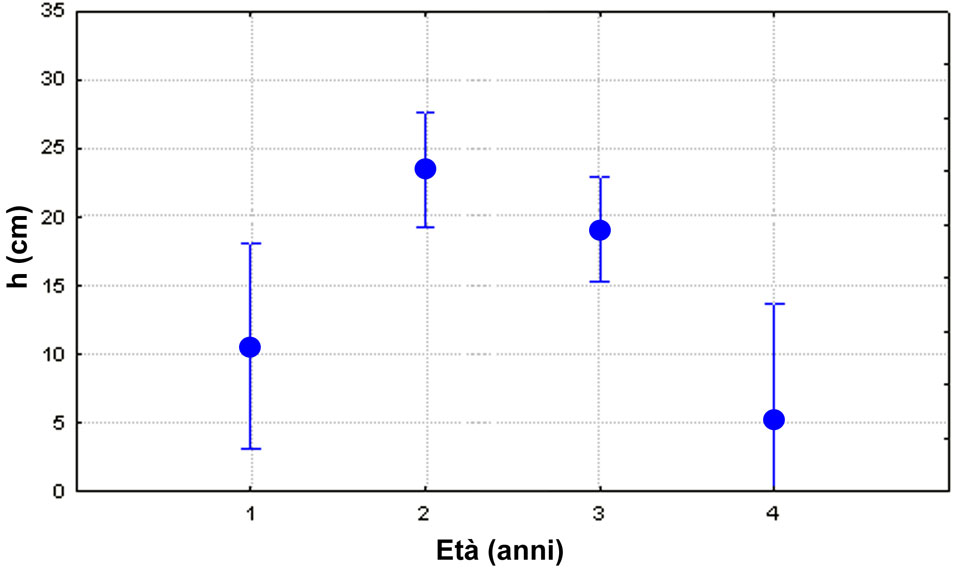
Il test ANOVA, condotto sull’altezza media dei polloni di cerro, evidenzia differenze molto significative (F[3.386] = 6.36; p<0.01) fra le diverse età messe a confronto. In particolare il test di Duncan indica che l’altezza dei polloni di 2 anni (23.5 cm) non è significativamente diversa da quella dei polloni di 3 anni (19.1 cm), a sua volta superiore ma non in misura significativa rispetto a quella dei polloni di 1 anno (10.6 cm) mentre l’altezza media più bassa si osserva nei polloni di 4 anni (5.1 cm), significativamente separata dalle altre (Fig. 3). La spiegazione - in accordo con quanto osservato sull’impatto della brucatura in relazione all’età dei ricacci - è che la ceppaia al secondo anno reagisca al danno da brucatura, ma che già dal terzo anno la ripetuta brucatura determini un progressivo deperimento.
Fig. 3 - Altezza media dei polloni dominanti di cerro, distinti per età (le barre verticali indicano gli intervalli fiduciari per p<0.05).
Impatto della brucatura in relazione alla superficie della tagliata
Si premette che la superficie media delle tagliate campionate è di 22 270 m2 (SE = 3254 m2).
Complessivamente 12 tagliate rientrano nella classe di superficie A, 9 tagliate nelle classi B e D, 6 tagliate nella classe C.
Le tagliate della classe A per il 75% appartengono al grado di impatto I. Per la classe di superficie B, il valore di I si abbassa al 67%, per salire al 100% nelle tagliate della classe di superficie C. Nelle tagliate della classe D il valore di I scende al 56% (Tab. 6). Ne risulta che le tagliate di maggiori dimensioni sono effettivamente quelle in cui il danno registrato è inferiore, anche se, nell’insieme, i dati non evidenziano una precisa relazione fra l’entità del danno e le dimensioni delle tagliate. Il dato si spiega con il fatto che la forte pressione alimentare, indipendentemente dalla dimensione, determina un danno molto forte.
Tab. 6 - Impatto della brucatura sulle tagliate, in relazione alla loro superficie.
| Classe di Superficie | Grado di It (%) | ||
|---|---|---|---|
| T | ME | I | |
| A | 17 | 8 | 75 |
| B | 22 | 11 | 67 |
| C | 0 | 0 | 100 |
| D | 22 | 22 | 56 |
Discussione
Prima dello spopolamento della campagna, le intense utilizzazioni dei boschi, la diffusione della zootecnia e una alta pressione venatoria avevano finito per ridurre la densità dei capi di molte specie di ungulati selvatici, determinando addirittura in certi casi estinzioni locali. Si aggiunga che anche i boschi aziendali erano importanti per l’alimentazione del bestiame: il sottobosco non solo era utilizzato direttamente in situ, ma era asportato periodicamente per ricavarne fascina per uso familiare. L’uomo era dunque un competitore naturale degli ungulati selvatici, in quanto ne limitava le disponibilità alimentari, sia col pascolo domestico sia con la raccolta di prodotti ([24]).
A partire dagli anni ’50, l’abbandono delle aree rurali, l’espansione del bosco, la cessazione delle utilizzazioni nei cedui, la forte riduzione del pascolo, il controllo della caccia e la reintroduzione di capi selvatici hanno determinato una inversione di tendenza. A questo si aggiunga la scomparsa dei grandi predatori (lupo, lince, orso). Il risultato è che i cervidi, in molte zone d’Italia, si sono espansi al punto da divenire responsabili di impatti negativi in aree sia agricole sia forestali. In condizioni ottimali di habitat e senza interventi di controllo, il tasso di incremento delle popolazioni di cervo può variare dal 10 al 30%, valore che porta al raddoppio di una determinata popolazione nel giro di pochi anni.
Per quanto riguarda la situazione dei boschi del Pratese, essa presenta molte analogie con quella osservabile nella confinante zona montana ed in quella collinare della provincia di Bologna ([31], [21]) dove l’elevato livello di danno prodotto dagli ungulati ha scatenato una serie di conflitti sociali che hanno determinato provvedimenti drastici da parte degli amministratori locali.
Le dinamiche del fenomeno, tuttavia, non possono essere valutate appieno se non alla luce delle trasformazioni paesaggistiche del nostro Paese, che hanno comportato importanti ricadute sulla fauna selvatica. Alcune specie, plastiche e opportuniste, hanno tratto vantaggio dall’espansione del bosco (cinghiale, capriolo). Molte altre specie, invece, hanno trovato nelle variazioni paesaggistiche un elemento sfavorevole alla loro diffusione (perdita di habitat specifici, come seminativi, prati-pascoli o pascoli alberati, adiacenti a boschi). Di fatto il cotico erboso è un elemento fondamentale di biodiversità, se non altro per l’alimentazione di selvatici: gli spazi aperti sono fondamentali per molti ungulati come per molte specie di uccelli. Ecco che oggi diviene determinante la salvaguardia (o il recupero) di aree aperte, fondamentali per il nutrimento di molti ungulati selvatici.
È un fatto che, in molti territori appenninici, la scomparsa di spazi aperti, oggi rimboschiti per via naturale, ha determinato una riduzione dell’offerta alimentare, con conseguenze negative sul prelievo in bosco ([23], [9], [28]). Ecco che la conservazione (o il recupero) degli spazi aperti viene sempre più spesso proposto come forma di gestione finalizzata a contenere la pressione alimentare del cervo sugli ecosistemi forestali: proposte in tal senso vengono fatte per il territorio appenninico ([26], [12], [4]) come per gli ecosistemi forestali di altri Paesi europei, fra cui il Regno Unito ([16], [20]).
Per quanto riguarda gli aspetti gestionali, un fattore che può influire sui danni da ungulati selvatici è rappresentato dalle dimensioni delle tagliate. In accordo con quanto detto a proposito degli spazi aperti, le tagliate dei cedui possono rappresentare un arricchimento in termini di diversità soprattutto a scala territoriale: la loro presenza arricchisce di habitat il paesaggio, con ricadute per gli ungulati ([30]) e per l’avifauna ([22]).
In realtà l’attrattiva di un habitat per la selvaggina non dipende solo dalla disponibilità di cibo, ma anche da altri fattori ambientali, fra cui gli effetti di margine. In più di un caso, le maggiori densità di selvaggina non si riscontrano nelle aree con maggiore disponibilità di cibo, ma in quelle più ricche di margini, che esprimono (per i cervidi, come per molte specie di uccelli) una grande forza attrattiva. È dimostrato che nelle tagliate di piccole dimensioni l’impatto è effettivamente maggiore, poiché la zona ecotonale presenta caratteristiche di habitat favorevoli agli ungulati, che in tali ubicazioni possono pascolare e allo stesso tempo godere della protezione del bosco (è nota la diffidenza dei selvatici a spingersi al centro di ampie superfici scoperte). Si aggiunga che la risorsa alimentare decresce al diminuire di superficie, per cui è possibile che in tagliate ampie l’offerta alimentare sia superiore alla domanda e, conseguentemente, l’impatto sia minore. Analogamente a quanto osservato nelle fustaie, anche nei cedui, a parità di superficie, un’ampia tagliata registra, di regola, minore intensità di brucatura da ungulati rispetto a un insieme di piccole tagliate, in cui l’effetto margine si fa spiccato ([29], [19]). Nel nostro caso di studio è vero che le tagliate di maggiori dimensioni sono quelle in cui il danno registrato è leggermente inferiore; tuttavia al diminuire delle dimensioni il danno cresce ma non in misura significativa, in conseguenza della forte pressione alimentare che, indipendentemente dalla dimensione, determina un danno di grande entità. Quando la densità dei cervi supera certi limiti, nemmeno la dimensione delle tagliate può rappresentare uno strumento per attenuare i danni da consumo alimentare.
Per quanto riguarda la selettività del danno, in Val di Bisenzio si è osservato che cerro e, in minore misura, carpino nero sono le specie più colpite. Il castagno appare il meno colpito e più resiliente nei confronti del morso degli animali, specialmente in boschi puri.
La selettività del consumo alimentare nei cervidi è documentata da molte ricerche. Fra queste vale la pena di segnalare le indagini condotte nel Parco della Maremma da Giovannini ([18]) e sull’Alpe di Catenaia (provincia di Arezzo) da Cutini et al. ([13]), entrambe focalizzate sul consumo alimentare del capriolo. Nel Parco Regionale della Maremma è stato evidenziato che la specie più appetita è l’orniello, mentre sull’Alpe di Catenaia la specie più colpita risulta il cerro (il castagno è il meno danneggiato). La selettività del consumo alimentare da parte dei cervidi è documentata anche da una serie di indagini condotte recentemente in molte località degli Stati Uniti ([14], [33], [15]), che hanno messo in luce come la pressione alimentare possa apportare, col tempo, modifiche alla struttura e alla composizione specifica dei popolamenti forestali interessati dal disturbo.
Per quanto riguarda gli effetti del brucamento sull’accrescimento dei ricacci, va premesso che un ceduo di cerro mediamente dovrebbe presentare, a un anno dal taglio, un’altezza media dei ricacci di circa 1 m ([7]), con alcuni polloni alti fino a 2 m (soprattutto nel caso di cerro e castagno). Le altezze rilevate in questo studio sono nettamente inferiori alle medie di un bosco in cui non si abbia danno da brucatura; il fenomeno, ovviamente, comporterà ricadute negative sull’accrescimento del bosco.
Per quanto concerne il danno ai ricacci in relazione alla loro età, sempre nel Parco della Maremma ([18], [11]) è stato dimostrato che il danno è maggiore nei primi anni dopo la ceduazione, quando vengono brucati i getti apicali, mentre si attenua nei cedui più vecchi (a 4 anni dalla ceduazione, nel caso dei cedui maremmani), dove prevale il consumo di getti laterali o epicormici. La presente indagine ha fornito, in questo senso, risultati diversi: sia i rilievi condotti sull’altezza dei polloni sia quelli relativi ai danni dimostrano che, al secondo anno, la ceppaia reagisce al danno da brucatura, sebbene si tratti di una reazione effimera. Già a partire dal terzo anno la ripetuta brucatura determina infatti un progressivo deperimento dei ricacci (i maggiori danni si registrano in corrispondenza di 1 e 4 anni di età). La spiegazione è attribuibile al fatto che il cervo, rispetto al capriolo e al daino, esercita una pressione alimentare ben superiore e, di conseguenza, maggiori danni a carico delle ceppaie.
Conclusioni
Gestire in maniera sostenibile i popolamenti forestali significa in molti casi fare i conti con la componente faunistica di tali ecosistemi: il problema dell’eccessivo carico di ungulati selvatici, che compromette la rinnovazione dei boschi, è spesso prioritario.
Da sottolineare come in Francia, Paese che ha visto una espansione delle popolazioni di ungulati molto forte, l’art. 1 del Codice Forestale stabilisce che “lo sviluppo sostenibile delle foreste implica un armonioso equilibrio tra le componenti forestali e faunistiche che permetta la rinnovazione dei popolamenti forestali in condizioni economiche soddisfacenti per il proprietario. Questo equilibrio è perseguito mediante l’applicazione del piano di caccia definito alla sezione 3 del capitolo V del libro IV del Codice per l’Ambiente…”. Non a caso, proprio in Francia, in un’area forestale del Donon, al fine di limitare i danni dei cervidi ai popolamenti forestali, si è creato un rapporto di collaborazione tra cacciatori e selvicoltori, basato su un’attività venatoria finalizzata alla riduzione del carico delle popolazioni di cervidi, da associare a interventi selvicolturali (diradamenti forti, apertura di corridoi) in grado di accrescere la disponibilità alimentare ([3]).
Al contrario di quanto si pensa comunemente, non si tratta solo di un problema di densità della fauna selvatica, anche se in condizioni di forte squilibrio il problema del carico appare determinante. Soprattutto, il problema non è solo selvicolturale, ma va affrontato su scala territoriale: attività agricole, caccia, turismo, apertura di strade, vicinanza di aree protette, scomparsa di spazi aperti con conseguente riduzione dell’offerta alimentare, sono tutti fattori che possono avere forte influenza sui danni da selvaggina di una zona boscata. Non si possono sottovalutare le migrazioni stagionali, come il fatto che se viene creata un’area protetta dalla caccia, gli ungulati tendono a concentrarsi proprio lì, con pesanti interferenze anche nei territori limitrofi ([10]).
In ultima analisi, la soluzione del problema, come ormai sostenuto da molti studiosi in Italia e all’estero ([23], [30], [8]), è una gestione faunistica integrata, che a misure faunistico-venatorie affianchi misure di gestione forestale e ambientale.
Nel caso specifico della Val di Bisenzio, è stato dimostrato che l’elevata densità della popolazione di cervo ha causato una riduzione della rinnovazione agamica nei cedui, tramite una persistente brucatura nelle zone in cui è stato effettuato il taglio. Se la pressione alimentare dovesse persistere nel tempo con questa intensità, in considerazione della selettività del consumo, sono prevedibili modificazioni nella struttura e nella composizione specifica dei popolamenti interessati dai danni (le indagini future potranno dare risposte più chiare in tal senso). Come si è visto, il problema dipende in parte dalla densità della popolazione di cervo, in parte dalla limitata presenza di spazi aperti, in cui potrebbe essere convogliata buona parte della domanda alimentare. Quindi, oltre a una caccia di selezione che contenga il numero dei capi presenti, è necessario affrontare il problema attraverso un’adeguata gestione del territorio. Per quanto riguarda gli aspetti selvicolturali, si tratta innanzitutto di creare, dove possibile, tagliate più ampie in modo tale da non amplificare l’effetto margine. Va da sé che l’ampiezza delle tagliate potrà avere efficacia solo a condizione che si riesca a contenere il numero di capi di cervo presenti.
Ringraziamenti
Lavoro svolto nell’ambito dell’Unità di ricerca “Gestione della fauna selvatica e rapporti di compatibilità con le attività agricole e l’ambiente”, afferente al Dipartimento GESAAF dell’Università di Firenze.
References
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar