Biological, ecological and genetic aspects of Narrow leaved ash (Fraxinus angustifolia Vahl) in the northeastern Italy
Forest@ - Journal of Silviculture and Forest Ecology, Volume 12, Pages 25-40 (2015)
doi: https://doi.org/10.3832/efor1628-012
Published: Aug 08, 2015 - Copyright © 2015 SISEF
Research Articles
Abstract
Narrow leaved ash (Fraxinus angustifolia Vahl) is an important forest tree belonging to peculiar and threatened ecosystems, where it forms mixed populations with other deciduous trees. Its range partly overlaps with that of European ash (Fraxinus excelsior) and interspecific cross is possible. These habitats are of the utmost importance due to their high level of biodiversity, but highly threatened by human activities. Since climate change is expected to favor the diffusion of narrow leaved ash, a study was carried out with the aim of enhancing our knowledge of the biological, ecological and genetic aspects of this species. Twelve populations of narrow leaved ash from north-eastern Italy were sampled and their genetic variability was analyzed by means of six molecular markers (nuclear microsatellites), of which 4 gave consistent and repeatable results. Overall, 113 alleles were observed, of which 38 were present in only one population (private alleles). Consistent levels of genetic variability was found (mean number of alleles per locus = 7.08, effective number of alleles per locus = 4.37, expected heterozygosity = 0.613), though lower than those reported in the literature. Fixation index was very close to zero, indicating a good agreement with Hardy-Weinberg’s expectations. The genetic differentiation among populations was significant (both GST e FST as high as 0.102), although it was not possible to correlate the distribution of genetic variability with ecological or geographical factors. Genetic pools of F. excelsior and F. angustifolia were distinct, though several individuals showed intermediate genetic characteristics, especially in the population from Grotta di Farra where both species are present and many individuals showed intermediate morphological features. The presence of individuals possibly originated by interspecific cross should be taken into account in the case of seed collection. Our results may contribute to a rationale management of genetic resources of narrow leaved ash, also considering the current legislation on forest reproductive material.
Keywords
Narrow Leaved Ash, nSSR Markers, Genetic Variability, Genetic Differentiation, Interspecific Hybridization, Regions of Provenance, Seed Stands
Introduzione
Le foreste sono soggette a numerosi fattori di stress, per la maggior parte dovuti all’azione antropica: deforestazione o gestione irrazionale, frammentazione degli habitat, introduzione di specie alloctone, ampliamento di superfici urbanizzate o destinate ad attività agricole, ecc. La situazione è resa più problematica dai cambiamenti climatici in atto, i quali possono accentuare le difficoltà di crescita e sopravvivenza degli alberi, nonché alterare i livelli di biodiversità e la struttura genetica propri dei popolamenti forestali. Tale aspetto riveste particolare importanza nelle aree periferiche e/o marginali di diffusione delle varie specie, quali in particolare l’area mediterranea ([18], [33], [21]). La più importante strategia per la salvaguardia delle foreste risiede nella conservazione del loro potenziale adattativo, il quale è strettamente legato ai livelli di variabilità genetica presenti nell’ecosistema e tra individui appartenenti alla stessa specie ([38], [19]). La differenziazione genetica tra popolamenti deriva a sua volta dall’azione di numerose forze evolutive, tra cui la pressione selettiva a differenti condizioni ecologiche e la deriva genetica sono le più importanti. Il flusso genico tende a contrastare tale processo, riducendo l’eterogeneità delle frequenze alleliche tra le varie popolazioni.
La caratterizzazione della variabilità genetica all’interno e tra popolazioni di specie forestali rappresenta quindi un presupposto fondamentale per la definizione di strategie volte a salvaguardare la loro biodiversità. I marcatori genetici rappresentano uno strumento di grande importanza per ricavare dati e informazioni sui livelli di variabilità e sulla struttura genetica delle popolazioni ([28]), nonché per fornire indicazioni relative alla gestione dei materiali forestali di propagazione, soprattutto nel caso in cui l’obiettivo principale degli interventi sia quello della ricostituzione di ecosistemi naturaliformi ([9], [39]).
Il frassino ossifillo (Fraxinus angustifolia Vahl) è un costituente importante delle foreste ripariali delle pianure alluvionali, ove forma popolamenti misti con altre latifoglie. Si tratti di ambienti preziosi dal punto di vista della biodiversità in essi presente, ma ormai rari a seguito dell’impatto negativo delle azioni antropiche. Poiché le modificazioni climatiche in atto potrebbero favorire la diffusione delle specie a carattere maggiormente termofilo, appare importante approfondire le conoscenze sulle risorse genetiche del frassino ossifillo.
La specie oggetto di analisi
Il genere Fraxinus appartiene alla famiglia delle Oleaceae e comprende 43 specie distribuite nelle aree temperate e subtropicali dell’emisfero settentrionale, di cui 24 in ambito europeo ([40]). Si tratta quasi esclusivamente di alberi decidui, sebbene siano presenti anche alcuni esempi di cespugli sempreverdi, diffusi soprattutto in zone semiaride. La maggior parte delle specie (circa i due terzi) è caratterizzata da impollinazione anemofila: le rimanenti (entomofile) sono diffuse prevalentemente in Asia.
In Italia crescono tre specie di frassini: il frassino maggiore o comune (Fraxinus excelsior L.), il frassino ossifillo o meridionale (F. angustifolia Vahl) e l’orniello (F. ornus L.). Mentre le prime due appartengono alla sezione Fraxinaster e presentano una impollinazione anemofila, l’orniello (sezione Ornus) presenta fiori molto attrattivi per gli insetti. Frassino maggiore ed ossifillo sono strettamente correlati e non sempre la loro distinzione è facile, soprattutto nell’ambito di boschi in cui si trovano entrambe le specie. La situazione è complicata dal fatto che le due specie possono dare origine ad individui ibridi ([32], [17], [11]). Benché si tratti di una specie naturalmente molto variabile, nell’ambito del frassino ossifillo sono state individuate alcune sottospecie, le cui differenziazioni si basano soprattutto sulla forma dei frutti e delle foglie e che presentano anche una distribuzione geografica differenziata: angustifolia (bacino mediterraneo occidentale), oxycarpa (Europa centrale e meridionale) e syriaca (dalla Turchia fino al limite orientale di diffusione della specie). La classificazione intraspecifica è tuttavia resa difficoltosa dalla presenza di numerose forme intermedie.
Il frassino ossifillo è comune nella penisola Iberica e lungo tutte le aree costiere del Mediterraneo. È diffuso anche in Marocco, Algeria e Tunisia, mentre ad est si spinge fino al mar Caspio (Fig. 1).
Fig. 1 - Distribuzione naturale del frassino ossifillo (F. angustifolia) in Europa (da [13]).
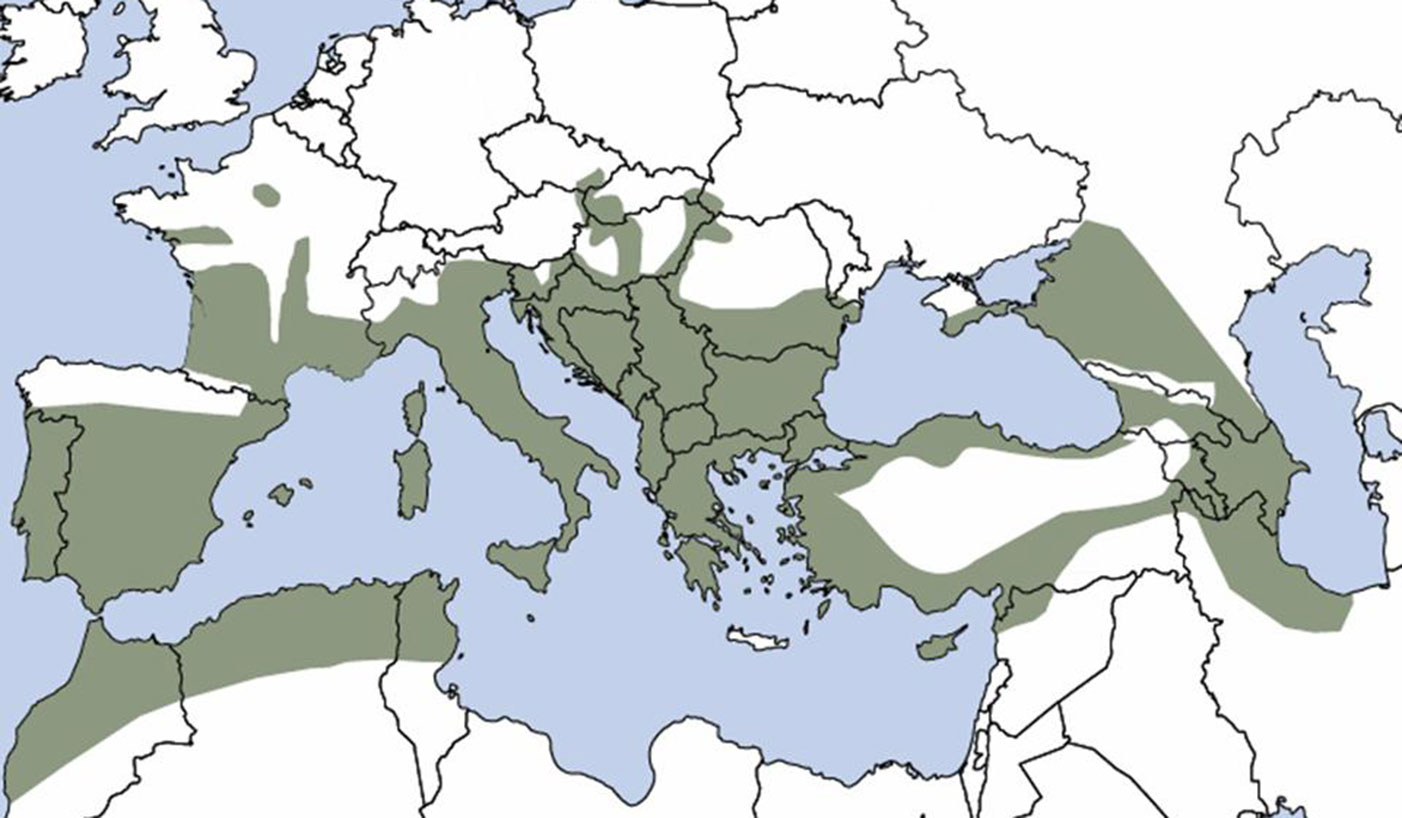
Nell’Italia settentrionale il frassino ossifillo è componente dei boschi planiziali relitti, frequente solo nella parte orientale e meridionale della Pianura Padana (Friuli Venezia Giulia, Veneto, Emilia Romagna); diviene più sporadico a partire dall’Emilia Romagna e dalla Lombardia occidentali verso nord-ovest, mentre ad ovest del Ticino è da considerare non autoctono. Importante componente delle selve umide dei litorali tirrenici, questa specie può risalire fino a quote submontane nell’Italia centro-meridionale, soprattutto lungo impluvi con condizioni pedo-climatiche favorevoli ([4]).
Criteri di distinzione tra frassino maggiore e ossifillo
Sebbene le due specie siano strettamente correlate, esistono alcuni caratteri che consentono la loro distinzione ([13]). Il più efficace riguarda il tipo di infiorescenza, che è composta (panicolo) nel frassino maggiore, mentre è semplice e priva di diramazioni laterali (racemo) nell’ossifillo. Ne consegue che le infiorescenze della prima specie contengono un numero di fiori molto maggiore rispetto a quelle della seconda. Questa caratteristica può essere facilmente osservata anche quando i semi sono maturi o addirittura già parzialmente staccati. Come detto, si tratta del carattere distintivo più affidabile, con l’eccezione di alcuni individui subandroici di frassino maggiore, nelle cui infiorescenze si trovano fiori ermafroditi lungo l’asse principale e fiori maschili sulle ramificazioni laterali. Dopo l’impollinazione questi ultimi cadono, per cui l’infruttescenza assume la forma di un racemo, facilmente confondibile con quella del frassino ossifillo.
Il colore delle gemme è normalmente nero nel frassino maggiore e bruno nell’ossifillo: tuttavia è presente (soprattutto nella seconda specie) una certa variabilità, che non sempre consente diagnosi precise. Nonostante l’elevato livello di variabilità ostacoli la distinzione delle due specie sulla base delle sole caratteristiche fogliari, si riportano di seguito le differenze più costanti nell’aspetto delle foglie ([30], [36], [1]):
- Frassino maggiore: foglioline di norma in numero di 7-15, con nervature secondarie irregolarmente ramificate, non sempre raggiungenti il margine fogliare. Dentelli del margine fogliare regolari, in numero maggiore rispetto alle nervature secondarie, con apice solitamente rivolto verso l’interno della foglia.
- Frassino ossifillo: foglioline di norma in numero di 3-11, con nervature secondarie perlopiù non o poco ramificate, raggiungenti il margine fogliare, il quale in corrispondenza delle nervature è dentellato, con dentelli irregolari, in numero pari al numero di nervature secondarie ed apice frequentemente rivolto verso l’esterno della foglia.
Risorse genetiche
Attualmente nel Registro Europeo dei Materiali di Base, previsto dalla Direttiva CEE n. 105 del 1999 sui materiali forestali di riproduzione, recepita nel nostro Paese con il Decreto Legislativo 386 del 2003 (⇒ http://ec.europa.eu/food/plant/plant_propagation_material/eu_marketing_requirements/forest_material_en.htm), sono registrate 561 provenienze di frassino ossifillo, di cui 491 nella categoria “Identificati alla Fonte” (430 Fonti di Seme e 61 Soprassuoli), 37 in quella “Selezionati” (tutti Soprassuoli) e 33 in quella “Qualificati” (4 Arboreti da Seme e 29 Genitori); la maggior parte di tali provenienze (370) è localizzata in Spagna, cui segue come importanza la Croazia.
In Italia sono stati catalogati complessivamente 32 popolamenti forestali idonei alla raccolta di seme di frassino ossifillo, di cui uno solo appartenente alla categoria “Selezionati” (Tab. 1).
Tab. 1 - Elenco dei materiali forestali di base per il frassino ossifillo in Italia. Tutti i popolamenti appartengono alla categoria “Identificato alla fonte”, eccetto l’ultimo che appartiene alla categoria “Selezionati”). (MdB): Materiali di Base; (SO): soprassuolo; (FS): fonte di semi.
| Regione | Popolamento | Codice Regionale | Tipo di MdB | Riferimento legislativo Regionale |
|---|---|---|---|---|
| Lombardia | Magenta/Robecco sul Naviglio (MI) | fan/B-BP/LOM-MI003 | SO | DGR 8/6272 del 21/12/2007, DD 4380 del 21/5/2012 |
| Marmirolo (MN) | fan/B-BP/LOM-MN001 | SO | ||
| Marcaria (MN) | fan/B-BP/LOM-MN006 | SO | ||
| Sicilia | B. Ficuzza | Sicilia 2 | SO | Piano Forestale Regionale DP 158/S.6/SG del 10/04/2012 |
| V. Anapo | Sicilia 16 | FS | ||
| B. Isnello | Sicilia 27 | FS | ||
| FriuliVenezia-Giulia | San Vito al Tagliamento (PN) | B/FVG (125) | FS | LR 11/2002, DD 1917/2010 |
| Bosco Baredi - Selva d’Arvonchi(Muzzana del Turgnano, UD) | B/FVG (123) | FS | ||
| Buja, ponte sul Corno (UD) | B/FVG (001) | FS | ||
| Muzzana del Turgnano, Bonifica di Muzzana (UD) | B/FVG (015) | FS | ||
| Sesto al Reghena (PN) | B/FVG (126) | FS | ||
| Buja, Case Rafael (UD) | B/FVG (067) | FS | ||
| Palude di Sablici (Doberdò del Lago, GO) | A/FVG (124) | FS | ||
| Isola della Cona (R.N. Foce Isonzo) - Staranzano, GO | A/FVG (122) | FS | ||
| Bosco "Baredi" - Selva d’Arvonchi (Muzzana del Turgnano, UD) | B/FVG (121) | FS | ||
| Veneto | Lison (Portogruaro, VE) | B/VEN (B/VE 013) | FS | DGR 3263 del 15/10/2004 |
| Lama Polesine (Ceregnano, RO) | B/VEN (B/VE 106) | FS | ||
| Torretta/Tartaro (Legnago, VR) | B/VEN (B/VE 131) | FS | ||
| S. Martino di Colle Umberto (Colle Umberto, TV) | B/VEN (B/VE 018) | FS | ||
| Isola d.Certosa/Laguna di Venezia (Venezia) | B/VEN (B/VE 105) | FS | ||
| Alvisopoli (Fossalta di Portogruaro, VE) | B/VEN (B/VE 148) | FS | ||
| Villa Furlanis/Portovecchio (Portogruaro, VE) | B/VEN (B/VE 149) | FS | ||
| Belfiore (Pramaggiore, VE) | B/VEN (B/VE 150) | FS | ||
| Emilia-Romagna | Boschi di Carrega (PR) | 330-fox-0023 | SO | LR 10/2007 DD 5205/2008 |
| Bosco della Mesola (FE) | 220-fox-0040 | SO | ||
| Santa Giustina (FE) | 220-fox-0041 | SO | ||
| Bosco Panfilia FE) | 120-fox-0042 | SO | ||
| Pineta di Classe (RA) | 220-fox-0045 | SO | ||
| Carpi Nord (MO) | 120-fox-0102 | FS | ||
| Gorgo Antonelli (FE) | 210-fox-0145 | SO | ||
| Bolognina di Crevalcore (BO) | 120-fox-0148 | FS | ||
| Toscana | San Rossore - San Giuliano Terme (PI) | Fan/C/TOS-0026 | SO | - |
All’interno del progetto europeo EUFGIS (⇒ http://portal.eufgis.org/), inoltre, sono monitorate e oggetto di studio 18 provenienze europee, in parte coincidenti con i materiali di base iscritti nei Registri Nazionali ed Europeo, di cui due in Italia (San Rossore, in provincia di Pisa, e Grotticelle, in provincia di Potenza).
Aspetti biologici
Il frassino ossifillo è una specie anemofila. Le sue infiorescenze possono essere costituite da fiori solo maschili, solo ermafroditi oppure da un insieme dei due. Non sono mai state osservate infiorescenze costituite unicamente da fiori femminili, nemmeno con residui di stami. Poiché ciascun individuo porta sia fiori maschili che bisessuati, si può affermare che le piante risultano essere ermafrodite. La fioritura del frassino ossifillo è più precoce rispetto a quella del maggiore. Le infiorescenze fuoriescono da gemme laterali in tardo inverno (tra la fine di gennaio e l’inizio di marzo), da 2 a 11 settimane prima dell’espansione delle foglie: tale intervallo risulta più accentuato in ambito mediterraneo. Il frassino ossifillo è protogino, cioè la recettività degli stami dei fiori ermafroditi precede la maturazione del polline. Tuttavia, poiché vi è una considerevole sovrapposizione tra le due fioriture, l’autofecondazione è di fatto possibile. In presenza di stagioni autunnali ed invernali molto miti, la fioritura può essere anticipata al tardo novembre, subito dopo la caduta delle foglie. In tal caso, però, le infiorescenze possono essere danneggiate dai freddi invernali, per cui la produzione sementiera risulta spesso nulla. Le gelate sono infatti considerate il più importante fattore limitante la diffusione della specie: tuttavia, si ritiene che il frassino ossifillo trarrà vantaggio dai mutamenti climatici ampliando il suo areale nei prossimi anni ([13]).
Le infiorescenze presentano mediamente 15 fiori (leggermente di più nel caso di strutture miste, costituite cioè sia da fiori maschili che ermafroditi), quindi in numero nettamente inferiore rispetto al frassino maggiore. Lo sfasamento temporale di fioritura ostacola la formazione di ibridi tra frassino ossifillo e maggiore, anche laddove le due specie coesistono: tuttavia, soprattutto in determinate annate, la fine della fioritura della prima specie può sovrapporsi all’inizio di quella della seconda, per cui in realtà individui ibridi possono effettivamente originarsi, anche in numero consistente. È comunque più probabile trovare ibridi su portaseme appartenenti al frassino maggiore, in quanto il periodo di sovrapposizione tra la recettività dei fiori femminili del F. excelsior e il rilascio del polline di F. angustifolia è maggiore rispetto alla situazione inversa.
Aspetti ecologici
Il frassino ossifillo è una specie tendenzialmente eliofila, più termofila rispetto al frassino maggiore, come testimoniano il baricentro distributivo meridionale e la diffusione a quote minori, di temperamento mesoigrofilo. Predilige suoli alluvionali con reazione sub-neutra, fertili e profondi, con granulometria sottile, mostrando tolleranza per contenuti in argilla anche elevati. Il frassino ossifillo presenta una resistenza a situazioni di temporanea sommersione superiore rispetto a quella del frassino maggiore: rientra infatti nella composizione delle comunità forestali ripariali delle pianure e del tratto terminale dei corsi d’acqua (Populion albae), ed è specie diagnostica dei boschi paludosi con Quercus robur, Ulmus minor ed Alnus glutinosa, propri dei substrati alluvionali fini con livello di falda mediamente elevato ([35]) Dal punto di vista conservazionistico, si tratta di comunità vegetali importanti in quanto presenti in forma relittuale e fortemente frammentate come conseguenza dell’antropizzazione. Gli habitat sensu direttiva 92/43 sono il 91F0 [Foreste miste riparie di grandi fiumi a Quercus robur, Ulmus laevis e Ulmus minor, Fraxinus excelsior o Fraxinus angustifolia (Ulmenion minoris)] e 92A0 (foreste a galleria di Salix alba e Populus alba).
Aspetti genetici
Il frassino ossifillo è stato oggetto di un limitato numero di studi genetici, a differenza del frassino maggiore. Nell’ambito del progetto europeo FRAXIGEN sono stati osservati complessivamente 8 aplotipi nell’ambito di 34 popolazioni appartenenti all’Europa mediterranea ed orientale. Per quanto riguarda la situazione italiana, sono stati individuati almeno due aplotipi, uno più meridionale e l’altro settentrionale (con la pianura padana come possibile zona di compresenza), cui potrebbe essere aggiunto un terzo aplotipo individuato al confine tra Slovenia ed Italia. La maggior parte degli aplotipi è stata osservata nella penisola balcanica, suggerendo come tale area possa essere stata un rifugio glaciale della specie. Non è tuttavia possibile escludere che anche le penisole iberica ed italiana abbiano rappresentato altre zone di rifugio, stante la presenza di aplotipi assenti nei Balcani.
La differenziazione genetica tra popolamenti è risultata modesta, sia nell’ambito di popolamenti che si trovano nello stesso Paese (ad es., Grecia - [27]) che tra popolamenti appartenenti ad ambiti geografici più lontani. La situazione presenta quindi grandi analogie con quanto già evidenziato a proposito del frassino maggiore, quanto meno nel nostro paese ([12]).
Scopo del lavoro
Lo scopo dello studio è stato quello di analizzare la variabilità genetica presente nel frassino ossifillo nell’ambito dell’Italia nord-orientale. Il mutamento climatico attualmente in atto potrebbe infatti favorire la diffusione spontanea di specie tendenzialmente termofile ([13]), sebbene su tale previsione il dibattito sia tuttora aperto e non manchino opinioni contrastanti ([8]). Le stesse attività selvicolturali, soprattutto se a prevalente finalità di ricostituzione naturalistica, potrebbero in futuro prevedere un maggior ricorso a specie caratterizzate da più elevata tolleranza alle alte temperature. Il quadro complessivo viene reso ancor più complesso dalla recente diffusione del patogeno Chalara fraxinea, individuato in Italia nord-orientale a partire dal 2009 ([26]) ed in grado di colpire tutte le specie italiane del genere Fraxinus. Gli effetti di tale fitopatia non sono tuttavia ancora del tutto chiari, sebbene le aree caratterizzate da climi freschi appaiono essere quelle più a rischio ([7]).
Nell’ambito del presente studio sono stati valutati 12 popolamenti mediante il ricorso a marcatori molecolari di tipo microsatellite ([37]). Sono anche stati approfonditi i rapporti intercorrenti tra la specie ed il frassino maggiore, cercando di evidenziare eventuali fenomeni di introgressione genetica tra le due specie. Infine, i risultati ottenuti sono stati discussi alla luce della Direttiva Europea 1999/105/CE, che si occupa dell’utilizzazione di materiali forestali di propagazione e stabilisce principi e criteri sui quali la filiera vivaistica deve basarsi. Tra questi uno degli aspetti più rilevanti è la definizione di Regioni di Provenienza, cioè aree caratterizzate da elevata omogeneità dei parametri ecologici e sulle quali si trovano popolazioni geneticamente omogenee ([3]).
Materiali e metodi
I popolamenti oggetto di studio
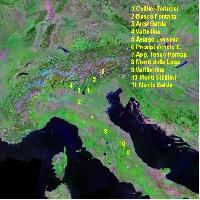
Il lavoro ha preso in considerazione 12 popolamenti di frassino ossifillo (Fig. 2), sulla cui origine autoctona si può essere ragionevolmente certi. Cinque di tali popolamenti sono localizzati nell’ambito della Regione Veneto (Fig. 3, Fig. 4, Fig. 5), 3 in Friuli Venezia Giulia (Fig. 6), altrettanti in Emilia Romagna ed uno in Lombardia. Alcune caratteristiche geografiche dei popolamenti di frassino ossifillo oggetto di studio sono riportate nella Tab. 2.
Fig. 2 - Localizzazione geografica delle popolazioni analizzate nel presente studio (in rosso: Fraxinus angustifolia; in giallo: Fraxinus excelsior).
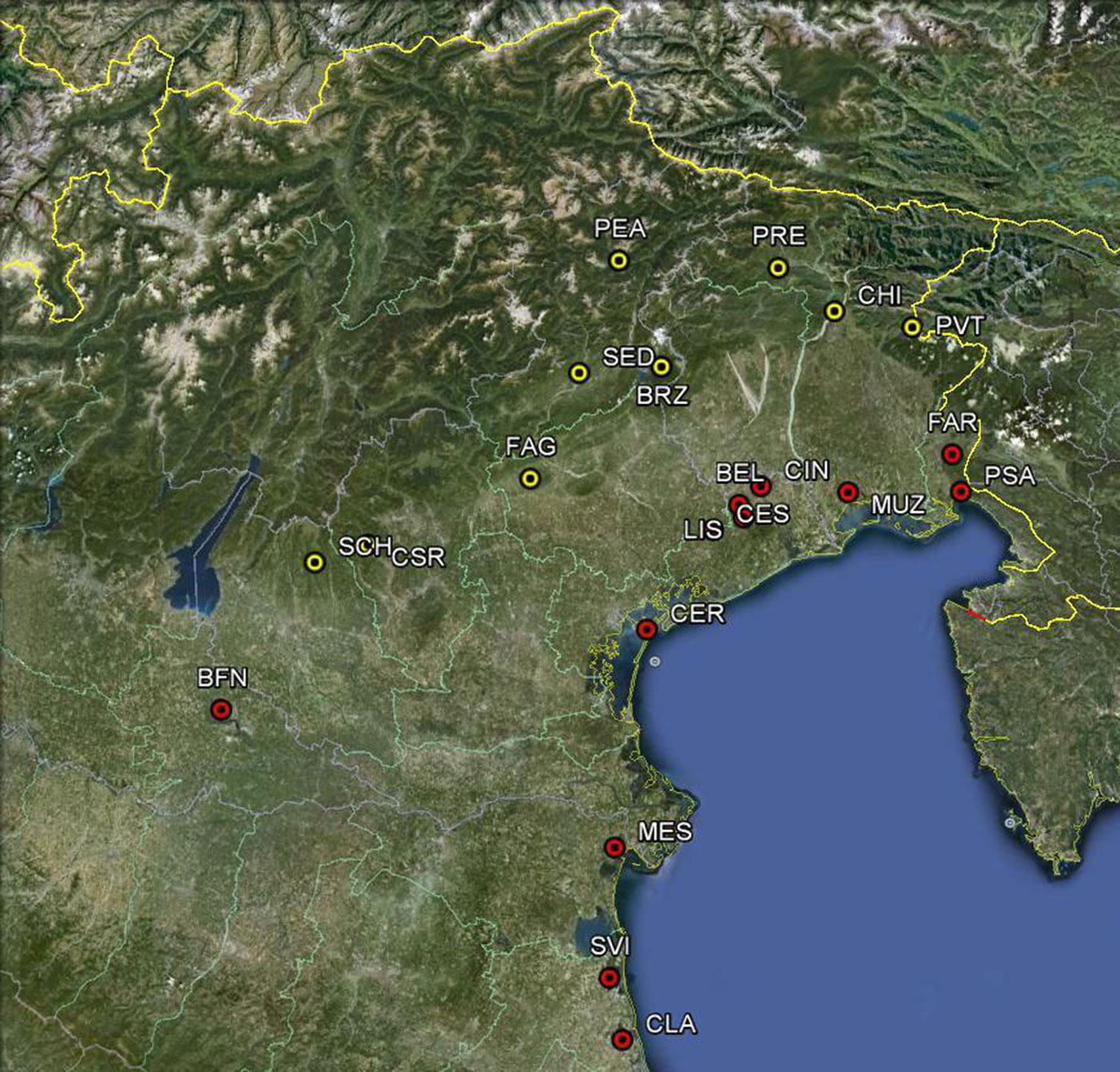
Fig. 3 - Maestoso esemplare di frassino ossifillo a Belfiore (Pramaggiore, VE).

Fig. 4 - Giovani frassini ossifilli nel popolamento dell’Isola della Certosa (VE).

Fig. 5 - Il popolamento dei Laghi di Cinto (Cinto Caomaggiore, VE).

Fig. 6 - Il popolamento di Palude Sablici (Doberdò del Lago, GO).

Tab. 2 - Popolamenti di frassino ossifillo analizzati nello studio e loro localizzazione geografica.
| Sigla | Popolamento (Comune) | Provincia | Lat. N | Long. E | Quota(m s.l.m.) |
|---|---|---|---|---|---|
| BFN | Bosco Fontana (Marmirolo) | Mantova | 45° 12’ | 10° 45’ | 25 |
| SVI | S. Vitale (Ravenna) | Ravenna | 44° 30’ | 12° 14’ | 5 |
| MES | Bosco della Mesola (Goro, Codigoro, Mesola) | Ferrara | 44° 51’ | 12° 15’ | 2 |
| CLA | Pineta Classe (Ravenna) | Ravenna | 44° 20’ | 12° 17’ | 2 |
| CER | Isola Certosa (Venezia) | Venezia | 45° 26’ | 12° 22’ | 1 |
| CES | Olmè (Cessalto) | Treviso | 45° 42’ | 12° 37’ | 2 |
| BEL | Belfiore (Pramaggiore) | Venezia | 45° 47’ | 12° 43’ | 5 |
| LIS | Lison (Portogruaro) | Venezia | 45° 44’ | 12° 44’ | 1 |
| CIN | Laghi di Cinto (Cinto Caomaggiore) | Venezia | 45° 49’ | 12° 48’ | 9 |
| MUZ | Muzzana (Muzzana del Turgnano) | Udine | 45° 48’ | 13° 08’ | 10 |
| FAR | Grotta (Farra di Isonzo) | Gorizia | 45° 54’ | 13° 32’ | 30 |
| PSA | Palude Sablici (Doberdò del Lago) | Gorizia | 45° 48’ | 13° 34’ | 7 |
Si riportano di seguito alcune brevi caratteristiche ecologico-vegetazionali dei popolamenti studiati.
- BFN: querco-carpineto della bassa pianura padana orientale.
- SVI, MES, CLA: pinete di pino domestico associate a querco-carpineti.
- CER: isola della laguna veneziana in parte occupata dal complesso di edifici di una vecchia caserma abbandonata, attualmente in fase di recupero (bosco costiero dei suoli idrici).
- CES: bosco misto caducifoglio tipico della pianura padano-veneta, con fustaia prevalentemente coetanea interessata da recenti interventi di diradamento e irrigazione al fine di migliorare il rinnovamento della farnia (querceto misto vegetante su falda sospesa).
- BEL: relitto di bosco planiziale inserito all’interno di un parco storico (ulmo-frassineto).
- LIS: relitto di bosco planiziale (querceto) parzialmente allagato in ambito di bassa pianura intensamente coltivata (querco-carpineto planiziale).
- CIN: cave senili rinaturalizzate cui si associano boschi umidi e corsi d’acqua di origine sorgiva, lungo i quali si sviluppano fasce boscate e siepi campestri (saliceto di Salix alba, querco-carpineto planiziale, alneta di Alnus glutinosa).
- MUZ, FAR: boschi umidi su substrati minerali dei terrazzi fluviali più evoluti.
- PSA: bosco palustre su substrati torbosi o minerali con prolungata inondazione.
Molti di tali popolamenti (BFN, MES, CLA, CER, LIS, MUZ e PSA) sono ufficialmente riconosciuti dalle competenti Amministrazioni Regionali quali fonte di semi certificati.
Per meglio analizzare i rapporti esistenti tra il frassino ossifillo e quello maggiore, vengono riportati anche alcuni dati di analisi effettuate in precedenza su popolamenti di quest’ultima specie, presenti nella stessa area geografica. Alcune caratteristiche di tali popolamenti sono riportate nella Tab. 3 e in Fig. 2 (per ulteriori dettagli si veda [2], [22], [12])
Tab. 3 - Popolamenti di frassino maggiore considerati nello studio e loro localizzazione geografica.
| Sigla | Popolamento (Comune) | Provincia | Lat. N | Long. E | Quota(m s.l.m.) |
|---|---|---|---|---|---|
| SCH | Schivazzi-Campe-Croce (Velo Veronese) | Verona | 45° 36’ | 11° 06’ | 950 |
| CSR | Contrada Sorto (Valdagno) | Vicenza | 45° 39’ | 11° 18’ | 740 |
| FAG | Fagarè (Cornuda) | Treviso | 45° 50’ | 11° 55’ | 200 |
| SED | Sedico (Sedico) | Belluno | 46° 07’ | 12° 06’ | 350 |
| PEA | Paeaio-Vinigo (Vodo di Cadore) | Belluno | 46° 25’ | 12° 15’ | 1.000 |
| BRZ | Broz (Tambre) | Belluno | 46° 08’ | 12° 25’ | 1.020 |
| PRE | Preone (Ampezzo) | Udine | 46° 24’ | 12° 52’ | 1.000 |
| CHI | Chianei (Trasaghis) | Udine | 46° 17’ | 13° 05’ | 500 |
| PVT | Ponte Vittorio (Taipana) | Udine | 46° 15’ | 13° 30’ | 500 |
Il campionamento
I campionamenti in bosco hanno coinvolto alcune foglie prelevate durante la stagione autunnale da non meno di 24 piante adulte per popolamento, sufficientemente distanziate tra di loro da rendere improbabili eventuali vincoli di parentela. Le piante campionate sono state ritenute appartenenti alla specie Fraxinus angustifolia sulla base di caratteri morfologici (numero e forma delle foglioline, colore delle gemme e, ove possibile, tipo di infiorescenza). Per alcuni popolamenti (SVI, MES, CES e MUZ) si è utilizzato direttamente il DNA già estratto, predisposto e fornito dal Centro Nazionale per lo Studio e la Conservazione della Biodiversità Forestale del Corpo Forestale dello Stato di Peri (Verona).
Analisi genetiche
Il DNA è stato estratto dal materiale campionato in bosco (foglie) mediante macerazione in azoto liquido ed utilizzo di specifici prodotti reperibili in commercio (nel caso specifico il QIAGEN® DNeasy plant mini kit), adottando le procedure suggerite dal produttore. Per ogni campione è stata ottenuta una concentrazione del DNA pari a 20 ng μl-1.
L’analisi genetica è stata condotta analizzando 6 loci nucleari polimorfici scelti tra i 16 microsatelliti isolati da Brachet et al. ([5]) e Lefort et al. ([20]) per il frassino maggiore, in quanto polimorfici e caratterizzati da una buona risoluzione del loro pattern elettroforetico, nonché sulla base dei risultati reperiti in bibliografia ([15], [23], [16]). Le caratteristiche di tali marcatori sono riportate in Tab. 4.
Tab. 4 - Caratteristiche descrittive dei marcatori microsatelliti nucleari usati per lo studio. I marcatori FEM(SATL)11 e FEM(SATL)12 sono stati successivamente esclusi dalle analisi, a causa della scarsa risoluzione e ripetibilità delle bande da loro originate. (pb): paia di basi.
| Microsatellite (nucleare) |
Primer utilizzati | Sequenza ripetuta |
Numero di alleli |
Peso molecolare (range in pb) |
|---|---|---|---|---|
| FEM(SATL)04 | 5’-TTCATGCTTCTCCGTGTCTC-3’ 5’-GCTGTTTCAGGCGTAATGTG-3’ |
(CA)n(AG)m | 14 | 154-186 |
| FEM(SATL)10 | 5’-TTGAGCAACATGTAATTATG-3’ 5’-AAATATCCGGTGCTTGTGTA-3’ |
(CT)n | 67 | 154-410 |
| FEM(SATL)11 | 5’-GATAGCACTATGAACACAGC-3’ 5’-TTCTTGAAGTAGTAGAACTA-3’ |
(GA)n(TA)m | - | - |
| FEM(SATL)12 | 5’-TTTTTGGAACCCTTGATTT-3’ 5’-ATTAAGAATGCCCGTCCATC-3’ |
(GA)nCA(GA)m | - | - |
| FEM(SATL)16 | 5’-TTTAACAGTTAACTCCCTTC-3’ 5’-CAACATACAGCTACTAATCA-3’ |
(CA)nCG(CA)m | 13 | 176-230 |
| FEM(SATL)19 | 5’-CTGTTCAATCAAAGATCTCA-3’ 5’-TGCTCGCATATGTGCAGATA-3’ |
(CA)nCGGC(CA)m | 19 | 142-200 |
| Media | - | - | 28.25 | - |
La reazione polimerasica è stata effettuata modificando il protocollo descritto in Brachet et al. ([5]) e Lefort et al. ([20]). In particolare, l’amplificazione del DNA è avvenuta utilizzando la seguente miscela di reazione: reaction buffer 1X, cloruro di magnesio 2.5 mM, dNTPs mix 0.2 mM, primer forward 0.2 µM, primer reverse 0.2 µM, Taq Polimerasi (Promega) 1 U, DNA campione 20 ng, H2O bidistillata sterile quanto basta per giungere a un volume finale di 25 µl.
Le reazioni PCR sono state condotte in un termociclatore Perkin Elmer GeneAmp® PCR System 9600, adottando i seguenti cicli di reazione: fase iniziale di 5 minuti a 96 °C; 30-35 cicli composti da 1 minuto a 94 °C (denaturazione del DNA), 1 minuto a 52 °C (annealing, cioè aggancio dei primer sul DNA campione), 2 minuti a 72 °C (sintesi del DNA); fase finale di 10 minuti a 72 °C.
La sequenza forward di ciascuna coppia di primer è stata marcata con un colorante fluorescente nel suo terminale 5’ (M-Medical S.r.l. e MWG-Biotech AG): IR-Dye 700 e, in alternativa, IR-Dye 800.
I prodotti di reazione sono stati separati elettroforeticamente dopo opportuna diluizione e denaturazione a 96 °C. L’elettroforesi è stata effettuata su gel di poliacrilamide denaturante (6.5%) utilizzando un sequenziatore (DNA 4200 Sequencer - LI-COR Biotechnology). Le corse elettroforetiche sono state condotte a 2000 V per 2 ore in TBE buffer 1X.
La determinazione dei polimorfismi è stata ottenuta attraverso il confronto del peso delle bande di DNA con quelle di uno standard marcato (IR-Dye 700 e IR-Dye 800) a peso molecolare noto (50-350 bp) e l’uso del software e-Seq (DNA Sequencing and Analysis Software).
Elaborazione dei dati
L’analisi della variabilità genetica dei popolamenti è stata effettuata attraverso una serie di parametri: le frequenze alleliche e i valori di eterozigosi sono stati calcolati ricorrendo al software GENALEX versione 6 ([29]). I valori di eterozigosi sono stati stimati seguendo i suggerimenti di Nei ([25]). La struttura e la distribuzione della variabilità genetica dei popolamenti sono state stimate attraverso la statistica F di Wright ([42], [43]), applicando il metodo riportato da Weir & Cockerham ([41]). La variabilità genetica totale è stata scomposta nelle sue componenti tra popolamenti e interna ai popolamenti. Le matrici delle distanze genetiche ([24]) sono state calcolate tramite il software GENDIST 3.6 e poi utilizzate per costruire il dendrogramma UPGMA attraverso il software NEIGHBOR 3.6 ([10]). Anche l’analisi delle coordinate principali è stata condotta sulle matrici delle distanze genetiche, utilizzando i pacchetti statistici DCENTER e EIGEN del software NTSYS ([34]). È infine stato utilizzato anche il software STRUCTURE ([31]), il quale definisce il numero ottimale di raggruppamenti nell’ambito del materiale in analisi, indipendentemente dalla loro origine, e calcola per ciascun individuo analizzato la percentuale di probabilità di appartenenza ai vari gruppi.
Risultati
Variabilità dei marcatori microsatellite
L’analisi genetica condotta in laboratorio sulla variabilità riscontrata a livello dei marcatori oggetto di studio ha consentito di identificare numerose varianti alleliche. Sono state evidenziate 113 bande, distribuite tra i quattro microsatelliti nucleari che hanno fornito risultati leggibili (Tab. 4). Da notare che, nel caso degli altri due marcatori (FEM11 e FEM12), non è invece stato possibile ottenere risultati sufficientemente chiari e ripetibili, per cui i dati ad essi relativi non sono stati considerati ai fini delle successive elaborazioni statistiche.
Sono stati evidenziati 38 alleli privati, cioè varianti geniche che sono presenti in un unico popolamento e che quindi rivestono una significativa importanza ai fini della definizione dei livelli di biodiversità presenti nell’ambito del materiale in esame. Il popolamento che ha presentato il maggior numero di alleli privati è risultato FAR con 10 alleli privati, seguito da MES con 7. All’opposto, CLA e SVI hanno evidenziato la presenza di un solo allele privato, mentre a CES e LIS non ne sono stati individuati del tutto.
Variabilità genetica interna ai popolamenti
I valori di variabilità genetica interni alle popolazioni sono riportati nella Tab. 5, dove N indica il numero medio di alleli per locus, Ne il numero effettivo di alleli per locus (parametro che, a differenza di quello precedente, tiene conto anche della frequenza con cui ciascuna variante allelica è presente), Ho l’eterozigosi osservata, He l’eterozigosi attesa (quella calcolata ipotizzando che le popolazioni si trovino in equilibrio di Hardy-Weinberg), FIS l’indice di fissazione (che indica l’eccesso - valori positivi - oppure la carenza - valori negativi - di omozigoti). He rappresenta anche la probabilità che due alleli scelti casualmente nell’ambito di una popolazione siano diversi.
Tab. 5 - Valori di ricchezza allelica e di diversità genetica nell’ambito delle 12 popolazioni di frassino ossifillo oggetto di analisi. (N): numero medio di alleli per locus; (Ne): numero effettivo di alleli per locus, (Ho): eterozigosi osservata; (He): eterozigosi attesa; (FIS): indice di fissazione
| Popolamento | N | Ne | Ho | He | FIS |
|---|---|---|---|---|---|
| BFN - Bosco Fontana | 8.00 | 4.36 | 0.583 | 0.586 | 0.005 |
| SVI - S. Vitale | 6.75 | 4.33 | 0.719 | 0.704 | - 0.021 |
| MES - Bosco della Mesola | 6.00 | 2.79 | 0.488 | 0.556 | 0.122 |
| CLA - Pineta Classe | 4.50 | 3.25 | 0.688 | 0.576 | - 0.194 |
| CER - Isola Certosa | 7.75 | 4.76 | 0.563 | 0.588 | 0.043 |
| CES - Olmè | 5.25 | 3.73 | 0.673 | 0.627 | - 0.073 |
| BEL - Belfiore | 6.50 | 4.11 | 0.688 | 0.630 | - 0.092 |
| LIS - Lison | 6.25 | 4.95 | 0.479 | 0.541 | 0.115 |
| CIN - Laghi di Cinto | 9.00 | 5.81 | 0.667 | 0.632 | - 0.55 |
| MUZ - Muzzana | 7.00 | 2.53 | 0.583 | 0.527 | - 0.106 |
| FAR - Grotta | 10.00 | 6.24 | 0.611 | 0.770 | 0.206 |
| PSA - Palude Sablici | 8.00 | 5.65 | 0.554 | 0.622 | 0.109 |
| MEDIA (dev. standard) | 7.08 (0.77) | 4.37 (0.51) | 0.608 (0.043) | 0.613 (0.038) | 0.008 (0054) |
Dai dati della tabella si evidenzia come i popolamenti presentino valori di ricchezza allelica molto differenti: in particolare, FAR, CIN, BFN e PSA hanno manifestato la più elevata ricchezza allelica, mentre, al contrario, MES, CES, CLA e MUZ hanno presentato i valori più ridotti di variabilità genetica interna al popolamento. Per quanto riguarda invece il livello di eterozigosi i valori più alti sono stati trovati a FAR e SVI, mentre quelli inferiori a LIS e MUZ. L’indice di fissazione è risultato mediamente prossimo a zero, denotando così un generale rispetto dell’equilibrio di Hardy-Weinberg.
Differenziazione genetica tra i popolamenti
Per quanto riguarda la differenziazione genetica tra i popolamenti (HT), è stato riscontrato un valore totale pari a 0.684; poiché la componente di tale variabilità dovuta a differenze interne alle popolazioni (HS) è stata stimata in 0.613, ne consegue un livello di divergenza tra di esse (DST) di 0.071, equivalente a un grado di differenziazione genetica (GST) di 0.102. In altre parole, di tutta la variabilità genetica che è stata identificata, poco più del 10% è dovuta a differenze tra popolamenti. A risultati analoghi si può pervenire analizzando i valori dei parametri F di Wright: anche in questo caso FIT rappresenta una stima della variabilità genetica totale (0.096), FIS di quella interna ai popolamenti (0.002) e FST il peso della componente di variabilità tra popolamenti rapportata al totale (0.102).
Una conferma sulla presenza di una consistente quota di variabilità genetica che si colloca tra i popolamenti deriva dall’AMOVA (Analisi della Varianza Molecolare), la quale prevede un peso di quest’ultima additittura superiore rispetto ai casi precedenti (Fig. 7).
Fig. 7 - Risultati dell’AMOVA tra i popolamenti di frassino ossifillo oggetto di studio.
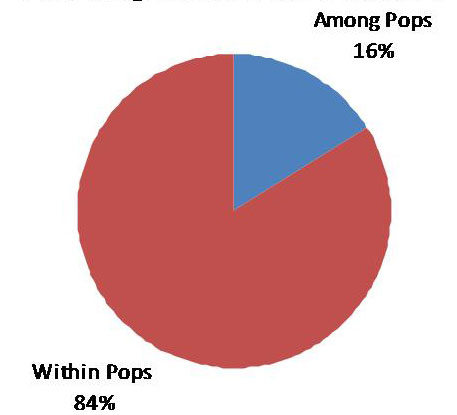
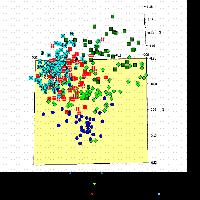
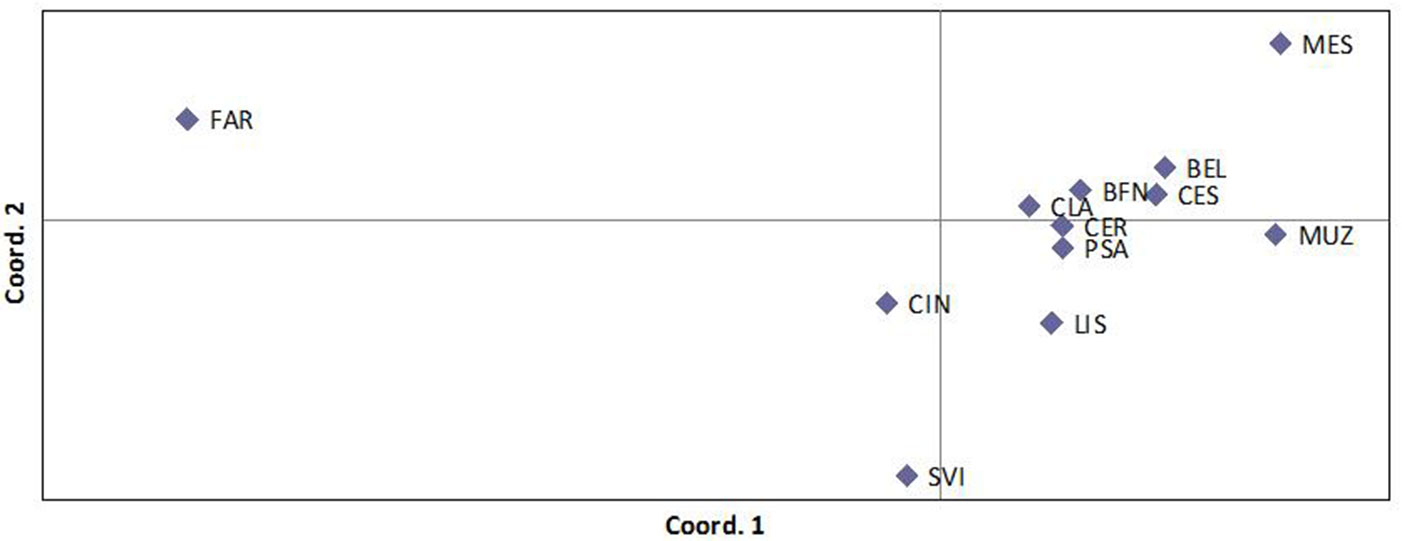
La distribuzione della variabilità genetica è stata valutata attraverso distinte metodiche. La prima è quella che prevede l’Analisi delle Coordinate Principali (PCoA), la quale raggruppa i popolamenti in base alle loro distanze genetiche. I risultati della PCoA sono riportati nella Fig. 8. Come si può notare, la situazione più evidente riguarda il popolamento friulano FAR, che è risultato nettamente differenziato da tutti gli altri. Questi sono invece risultati sostanzialmente simili e comunque non raggruppabili in funzione della loro localizzazione geografica. Ad esempio, i tre popolamenti emiliani si distribuiscono in modo irregolare nell’ambito della nuvola di popolamenti che si trova nella parte destra del grafico. All’opposto, i popolamenti di BEL, LIS e CIN, geograficamente molto vicini, non risultano geneticamente più simili rispetto agli altri.
Fig. 8 - Risultati dell’analisi delle coordinate principali relativa ai 12 popolamenti di frassino ossifillo analizzati nello studio e basati sulla variazione presentata a 4 loci microsatellite.
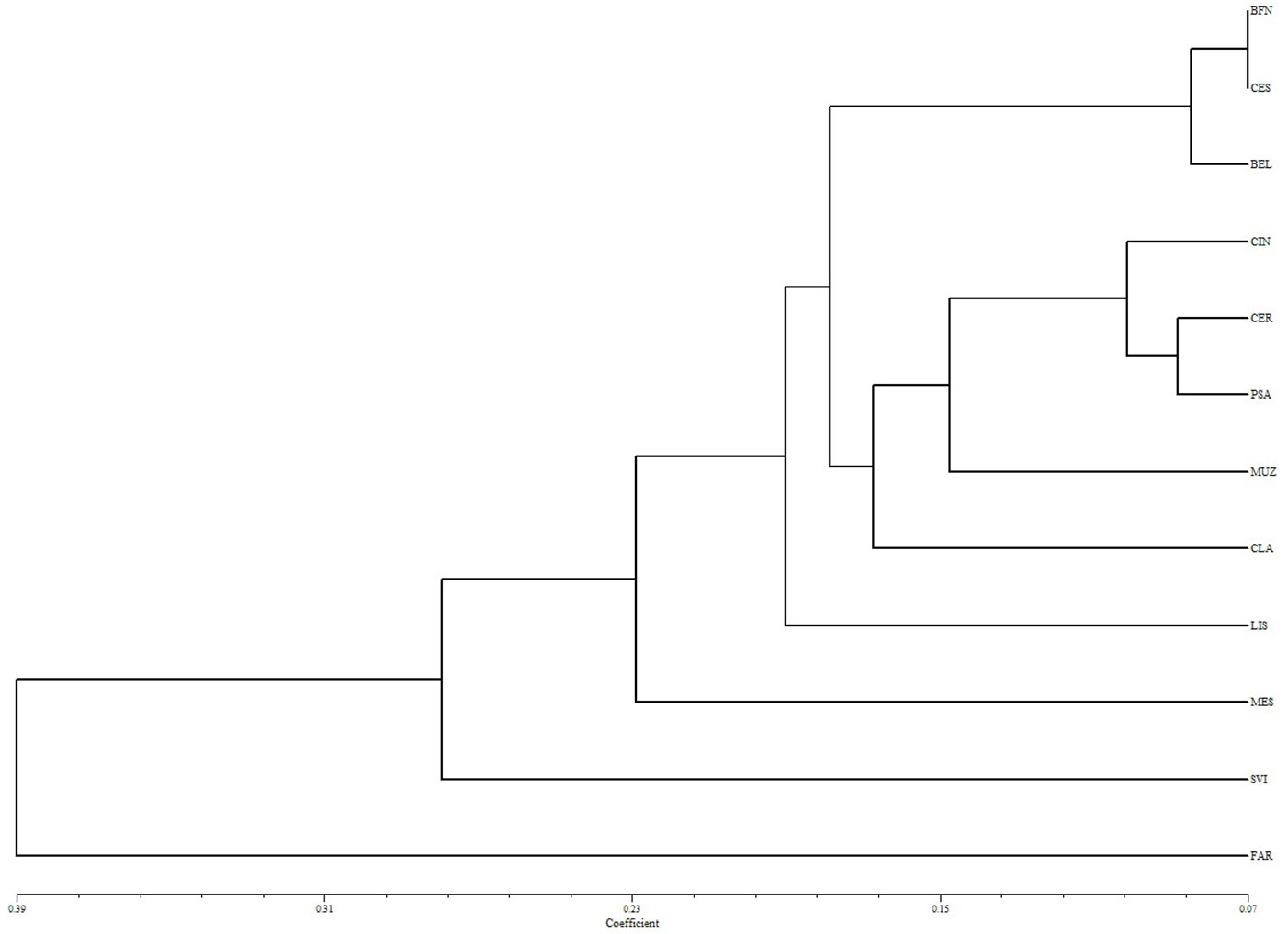
A risultati molto simili si perviene osservando il dendrogramma costruito sulla base delle distanze genetiche calcolate tra coppie di popolazioni (Fig. 9): anche in questo caso è evidente la netta differenziazione tra il popolamento di Grotta (FAR) e gli altri, nell’ambito dei quali invece non si ravvisano andamenti particolari.
Fig. 9 - Dendrogramma costruito sulla base delle distanze genetiche tra le popolazioni oggetto di studio.
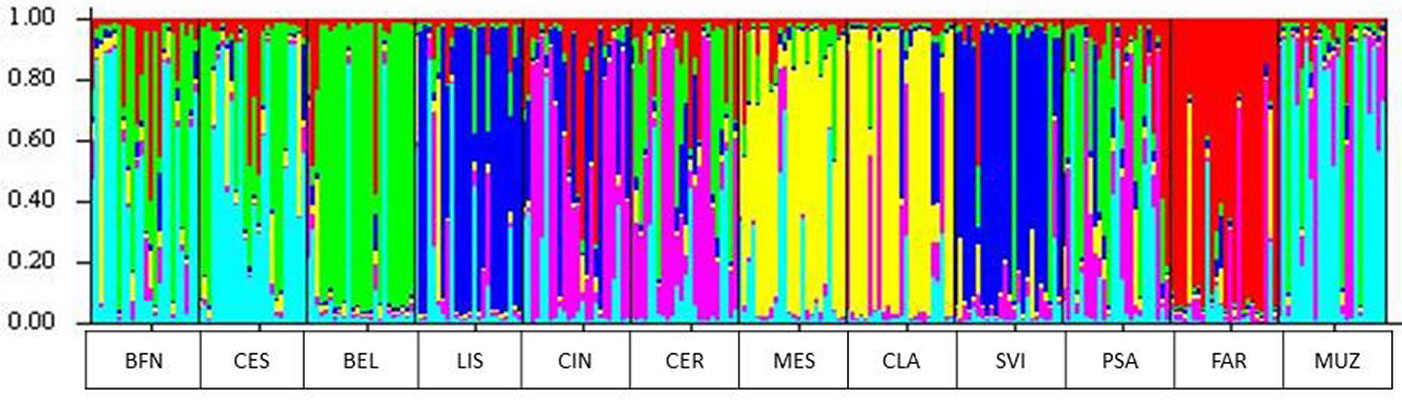
Dall’analisi STRUCTURE applicata alle popolazioni di frassino ossifillo è emerso come il numero ottimale di raggruppamenti genetici sia pari a 6. I vari individui si distribuiscono nell’ambito di tali gruppi come rappresentato in Fig. 10. Dalla figura emerge una situazione abbastanza confusa e poco correlata con parametri di natura geografica. Il popolamento lombardo (BFN), ad esempio, risulta simile a quello veneto di CES, ma anche a quello friulano di MUZ. Mentre FAR conferma la sua particolarità, i popolamenti veneti CIN e CER, presentano una certa similitudine, così come LIS con SVI e MES con CLA. Di più difficile definizione PSA, che presenta una situazione molto più complessa.
Fig. 10 - Risultati dell’analisi STRUCTURE effettuata sui 12 popolamenti di frassino ossifillo (K = 6).
Confronto tra Fraxinus angustifolia e F. excelsior
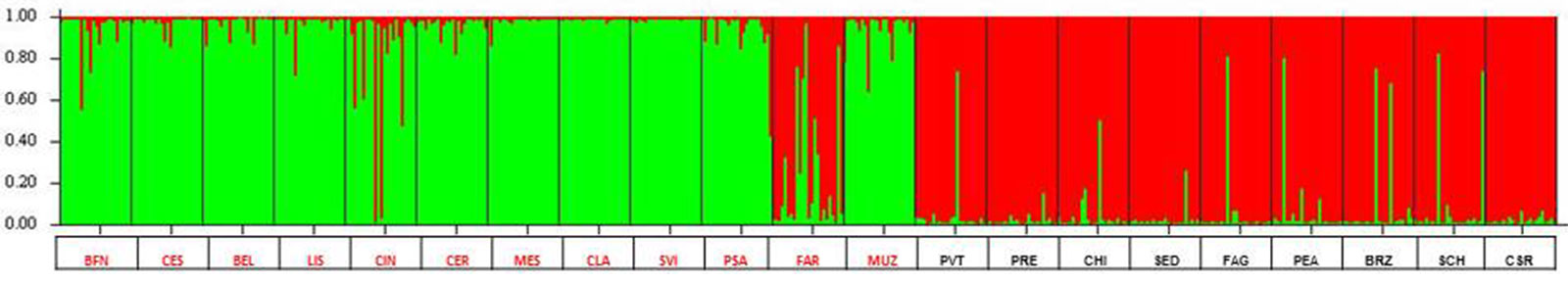
Per quanto riguarda le possibili interazioni genetiche tra le due specie di Fraxinus maggiormente diffuse nell’Italia settentrionale, appare significativo il risultato ottenuto elaborando i dati tramite la procedura STRUCTURE (Fig. 11). Dalla figura si evidenzia subito come le due specie risultino divise in modo abbastanza netto. L’eccezione più vistosa è quella del popolamento di Grotta (FAR), il quale, pur essendo originariamente stato identificato (e soprattutto campionato) come frassino ossifillo, presenta caratteristiche genetiche in gran parte riconducibili al frassino maggiore. Un certo numero di individui con caratteristiche intermedie si trovano anche in altri popolamenti, soprattutto BFN e CIN.
Fig. 11 - Risultati dell’analisi STRUCTURE relativamente a 12 popolamenti di frassino ossifillo e 9 di frassino maggiore dell’Italia nord-orientale. Il colore verde indica il pool genetico riconducibile alla specie Fraxinus angustifolia, quello rosso a F. excelsior.
Discussione e conclusioni
Il lavoro svolto ha consentito di approfondire le conoscenze sulle caratteristiche genetiche dei popolamenti di Fraxinus angustifolia dell’Italia nord-orientale e permette di trarre indicazioni che rivestono una certa significatività anche dal punto di vista pratico-applicativo. In particolare, si fa riferimento all’applicazione delle norme previste dal Decreto Legislativo 386/03, che recepisce la Direttiva CEE 105/1999 ed impone la certificazione del materiale forestale di propagazione. In dettaglio, la norma prevede l’istituzione delle Regioni di Provenienza, definite come “il territorio o l’insieme di territori soggetti a condizioni ecologiche sufficientemente uniformi e sui quali si trovano soprassuoli o fonti di semi sufficientemente omogenei dal punto di vista fenotipico e, ove valutato, dal punto di vista genotipico, tenendo conto dei limiti altimetrici ove appropriato” (art. 2, lettera g, D. Lgs 386/03). Nella definizione delle Regioni di Provenienza occorre tenere conto sia dei fattori di tipo ecologico in senso lato (climatici, pedologici, vegetazionali, ecc.) che di quelli genetici propri dei popolamenti che crescono spontaneamente nelle aree in esame ([3], [6]).
Come verificato per la maggior parte delle specie forestali ([14]), anche le popolazioni esaminate hanno presentato elevati livelli di variabilità genetica. Tuttavia, in un analogo studio effettuato in Grecia ([27]), si sono riscontrati valori di diversità genetica leggermente superiori: tale fatto può essere spiegato con la maggior dimensione dell’area di studio (che ha coinvolto l’intera penisola ellenica) e con una maggior prossimità di questa all’area balcanica, ritenuta la principale area di rifugio della specie durante l’ultima epoca glaciale ([13]). Va tuttavia ricordato come alcuni autori propongano anche le penisole iberica ed italiana quali possibili aree di rifugio, dal momento che vi sono stati identificati aplotipi di microsatelliti plastidiali praticamente assenti nell’area balcanica ([13]). Va inoltre tenuto presente che taluni parametri ricavati nel presente studio potrebbero essere influenzati da fenomeni di introgressione genetica tra il frassino ossifillo e quello maggiore: l’esempio più evidente è rappresentato dal popolamento di FAR, laddove gli elevati valori di alleli privati, ricchezza allelica ed eterozigosi sono probabilmente dovuti a una consistente percentuale di incrocio tra le due specie.
In linea generale, i popolamenti esaminati hanno presentato un buon accordo con i dati attesi sulla base della legge di Hardy-Weinberg (indice di fissazione medio 0.008). Tuttavia, se analizziamo i valori che si riferiscono ai singoli popolamenti, la situazione diventa eterogenea: è infatti possibile identificare situazioni di evidente eccesso di omozigoti (MES, LIS e i due popolamenti friulani: FAR e PSA) ed altre in cui invece è presente un numero di eterozigoti superiore a quello atteso (CLA, CES, BEL e MUZ). L’eccesso di omozigosi si può spiegare in diversi modi: presenza di alleli nulli (che inducono un’errata interpretazione di individui eterozigoti come omozigoti), effetto della strutturazione del popolamento in gruppi all’interno del quale l’incrocio risulta più probabile (effetto Wahlund). Tuttavia la causa più comune per spiegare l’eccesso di omozigoti è la presenza di inbreeding (incroci tra individui imparentati o addirittura di autofecondazione), che tende a rendere più probabile la comparsa di un genotipo omozigote rispetto alle unioni casuali. Nel citato studio di Papi et al. ([27]) effettuato in 8 popolamenti di frassino ossifillo in Grecia, è stato stimato un tasso di autofecondazione pari a 0.157. A sua volta, l’inbreeding molto spesso dipende da un basso numero di individui che costituiscono la popolazione, tra i quali forzatamente si instaurano vincoli di parentela più stretti. In specie allogame come il frassino ossifillo, l’eccesso di omozigosi è un fattore negativo, che indebolisce le popolazioni, innescando anche fenomeni di erosione genetica (depressione da inbreeding). Di conseguenza, un popolamento caratterizzato da un indice di fissazione significativamente positivo dovrebbe essere valutato con grande cautela nell’ottica di una destinazione alla produzione di materiale propagativo.
La distribuzione della variabilità genetica, pur essendo significativa la differenziazione tra di essi (GST e FST entrambi pari a 0.102), non è risultata correlabile ad alcun fattore geografico. Tale fatto può spiegarsi con la relativa esiguità dell’area considerata e con l’assenza di barriere (quali catene montuose) in grado di limitare il flusso genico tra di esse.
Benché i pool genici del frassino ossifillo e del frassino maggiore siano risultati distinti, alcuni individui hanno presentato caratteristiche genetiche intermedie: tale fenomeno è risultato evidente soprattutto a FAR, dove la maggior parte degli individui campionati è risultata più o meno simile al frassino maggiore. La spiegazione di ciò può derivare dal fatto che il popolamento cresce ai margini dell’areale di diffusione delle due specie, quindi in una situazione ove la loro compresenza è molto comune. Di conseguenza, fenomeni di ibridazione possono risultare più frequenti rispetto ad altre situazioni. A conferma di ciò, va detto che i frassini del popolamento in esame presentano anche caratteristiche morfologiche non del tutto chiare, che spesso creano notevoli difficoltà di definizione della specie (Pier Paolo Merluzzi, comunicazione personale). Individui originati da incrocio potrebbero anche essere presenti, sia pure con frequenza molto minore, in altri popolamenti: CIN in particolare, ma anche BFN (ove in effetti la diffusione del frassino maggiore è più consistente) e in misura ancora inferiore tutti gli altri popolamenti del Veneto e del Friuli Venezia Giulia. Al contrario, i popolamenti emiliano-romagnoli (MES, CLA e SVI) sembrano risentire in misura del tutto trascurabile di possibili introgressioni genetiche da parte del frassino maggiore: risultato spiegabile con la presenza sporadica di questa specie in tali aree.
La presenza di individui presumibilmente derivati da incrocio tra le due specie deve essere tenuta in grande rilievo nel caso in cui si proceda alla raccolta di materiale propagativo. Se infatti lo scopo è quello di ottenere materiale sicuramente riconducibile al frassino ossifillo, i popolamenti in questione dovranno essere evitati. D’altra parte, si può anche escludere che l’eventuale introgressione di geni provenienti dal frassino maggiore possa migliorare l’adattabilità del frassino ossifillo. Quest’ultima specie, infatti, è più termofila e in una prospettiva di cambiamenti climatici caratterizzati da un incremento delle temperature medie i fenomeni di incrocio potrebbero addirittura risultare controproducenti.
Ringraziamenti
Il presente lavoro è stato svolto con il contributo finanziario della Regione Veneto, Direzione Economia e Sviluppo Montano. Si ringrazia il Centro Nazionale per lo Studio e la Conservazione della Biodiversità Forestale del Corpo Forestale dello Stato di Peri (Verona), per la cessione del DNA relativo a quattro dei popolamenti di frassino ossifillo esaminati. Si ringraziano inoltre i dott. Daniele Savio e Pier Paolo Merluzzi per le informazioni fornite e per il supporto garantito durante alcune fasi di campionamento
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Online | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar