Ground beetles as indicators of past management of old-growth forests
Forest@ - Journal of Silviculture and Forest Ecology, Volume 14, Pages 162-174 (2017)
doi: https://doi.org/10.3832/efor2351-014
Published: May 15, 2017 - Copyright © 2017 SISEF
Research Articles
Abstract
Old-growth forests are terrestrial ecosystems with the highest level of biodiversity and the main environments for the study of conservation and dynamics of the forest system. In Mediterranean Europe, two millennia of human exploitation deeply altered the structural complexity of the native forests. Some animal groups, including insects, may be used as a proxy of such changes. In this paper we explored the possible effects of forest management on the functional diversity (species traits) of carabid beetle communities. Three old-growth forests of the Sila National Park were sampled by pitfall traps set up in pure beech, beech-silver fir and Calabrian black pine forests. In each forest, five managed vs. five unmanaged stands were considered. Managed sites were exploited until the sixties of the past century and then left unmanaged. More than 6000 carabid specimens belonging to 23 species were collected. The functional diversity in carabid groups is influenced by forest management especially in beech and beech-silver fir stands. Body size, specialized predators, endemic species and forest species were negatively affected by stand management. On the contrary, omnivorous ground beetles populations (or species) with a high dispersal power (macropterous) and large geographic distribution were positively influenced by stand management. In pine forests the old-growth community seems less sensitive to past management and more affected by soil evolution. Soil erosion and disturbance may reduce species diversity of ground beetles. Anyway, the composition of the carabid community shows that 50-60 years of forest restoration are enough for the reconstruction of a fairly diverse assemblage reflecting a “subclimax” situation.
Keywords
Functional Diversity, Species Traits, Forest Management, Old Growth Forests, Bio-indicators
Introduzione
Tra gli ecosistemi terrestri, le foreste sono quelli a maggior diversità genetica, specifica e sistematica ([24]). In passato, gran parte delle foreste europee sono state fortemente utilizzate ([17], [16]), con interventi - riconducibili a una selvicoltura intensiva - che hanno favorito una o poche specie a discapito di altre, provocando un cambiamento nella qualità degli habitat forestali, con conseguenze anche per la biodiversità ([9]). In tal senso un aspetto fondamentale per la definizione delle politiche di conservazione della biodiversità forestale riguarda la gestione delle aree forestali classificate come old-growth forests o boschi vetusti. Il termine foresta vetusta non ha una definizione univoca. Nell’ambito del progetto “Foreste Vetuste in Italia” ([3]) la definizione di foresta vetusta è la seguente: “Foreste in cui il disturbo antropico sia assente o trascurabile, caratterizzate da una dinamica naturale che determina la presenza, al loro interno, di tutte le fasi di rigenerazione, compresa quella senescente. Tale fase è caratterizzata da individui di notevoli dimensioni ed età; presenza di legno morto (alberi morti in piedi, rami e alberi caduti a terra); una flora coerente con il contesto biogeografico caratterizzata dalla presenza di specie altamente specializzate che beneficiano del basso grado di disturbo e di specie legate ai microhabitat determinati dall’eterogeneità strutturale”. Se nell’ambito forestale inizia ad avere una connotazione chiara, nello studio delle comunità e della biodiversità animale la nozione old-growth forest non ha una fisionomia ben precisa. Alcuni studi consentono di identificare in alcune caratteristiche di comunità indicatori di diverse situazioni e/o evoluzioni forestali ([5], [2], [28]). Più numerosi sono i lavori che mettono in relazione la struttura forestale con le caratteristiche e la composizione delle comunità animali e i problemi di conservazione ([19], [42], [1]). È noto in letteratura che le attività forestali influenzano la biodiversità complessiva dei boschi ([15]). Questi cambiamenti influenzano gli organismi positivamente o negativamente secondo le loro esigenze di habitat ([29]). I coleotteri carabidi sono insetti ampiamente utilizzati per valutare l’impatto dei cambiamenti degli habitat ([41]). In Europa, ad esempio, si hanno informazioni dettagliate sulle risposte dei carabidi nei confronti della gestione forestale nelle piantagioni di conifere e nelle foreste boreali ([10], [20], [28], [21], [36]). Per le foreste dell’area del Mediterraneo, storicamente modificate dall’uomo, non si conoscono invece gli effetti delle utilizzazioni forestali sulla carabidofauna ([44]). Inoltre, la maggior parte degli studi che hanno cercato di analizzare gli effetti della gestione forestale sui carabidi, ha spesso fornito risultati contrastanti. Parlando più in generale nel contesto europeo, la gestione delle foreste può far aumentare ([35]) o diminuire la ricchezza di specie di carabidi ([28]), oppure non avere alcun effetto su di essa ([27]). Effettivamente la gestione forestale, modificando svariati fattori come l’umidità, l’intensità luminosa, la lettiera, la necromassa, l’eterogeneità della composizione specifica e la struttura del bosco (vedi bibliografia in [46]) può influire sulla varietà di specie di coleotteri carabidi ([47], [32], [38]). Le caratteristiche ecologico-funzionali delle diverse specie possono spiegare la loro risposta alle variazioni ambientali ([22], [31]). La gestione delle foreste può infatti influenzare fortemente la presenza di specie di carabidi prettamente forestali e di quelle incapaci di volare, e di conseguenza poco mobili ([43]), mentre può favorire alcune specie onnivore ([23]). In questo contesto, il presente lavoro si propone di valutare gli effetti della gestione forestale sulle caratteristiche funzionali della carabidocenosi. Misurando la ricchezza delle specie presenti in diversi gruppi ecologici, lo studio ha inteso confrontare la diversità funzionale dei carabidi presenti in boschi vetusti e da tempo in condizione di abbandono colturale con popolamenti analoghi ma regolarmente sottoposti a interventi selvicolturali. Nello specifico si è voluto valutare se l’abbandono colturale del bosco favorisce la ricchezza funzionale dei carabidi; se le caratteristiche funzionali dei carabidi sono influenzate dai modelli colturali analizzati, riferibili alle foreste con caratteri di vetustà e quelle dove sono stati effettuati interventi di gestione; a quale livello di naturalità si trovano oggi le comunità di coleotteri carabidi associate a particelle gestite nel passato. Al riguardo si presume che nelle foreste con caratteri di vetustà, caratterizzate da un elevato grado di complessità strutturale, ci sia un maggior numero di specie a preferenze ecologiche forestali e di predatori specializzati con scarso potere di dispersione. Nelle foreste più o meno regolarmente sottoposte a interventi di utilizzazione potrebbero invece essere favorite specie generaliste o di spazi aperti, ad alto potere di dispersione, onnivore e xerofile ([4], [40], [34]).
Materiali e metodi
Aree di studio
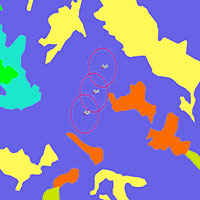
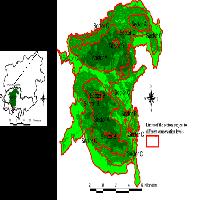
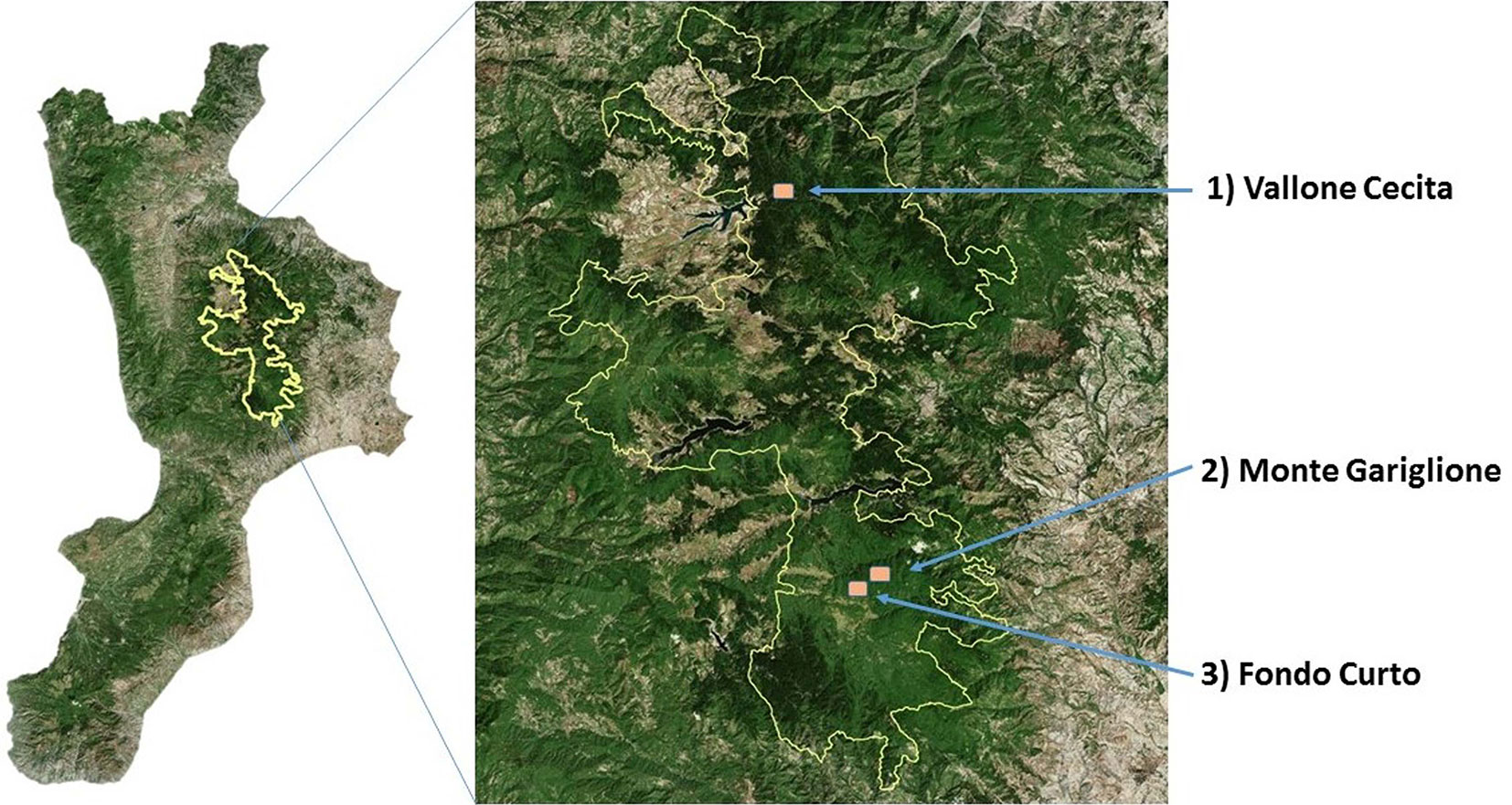
La ricerca è stata condotta in tre foreste del Parco Nazionale della Sila: Monte Gariglione (comune di Taverna, provincia di Catanzaro e comune di Mesoraca, provincia di Crotone), Fondo Curto (comune di Taverna, provincia di Catanzaro) e Vallone Cecita (comune di Longobucco, provincia di Cosenza - Fig. 1). In ciascuna foresta sono state individuate aree di bosco non gestito, ovvero bosco vetusto o con caratteri di vetustà in abbandono colturale, e aree di bosco gestito, ovvero bosco con analoghe caratteristiche stazionali e vegetazionali di quello vetusto, individuato in aree adiacenti ma sottoposto a interventi selvicolturali, utilizzato come confronto (Tab. 1).
Fig. 1 - Area di Studio. Parco Nazionale della Sila.
Tab. 1 - Siti campionati e loro caratteristiche generali.
| Località | Coordinate (UTM 33 S) |
Altitudine (m s.l.m.) |
Esposizione | Superficieconsiderata | Tipologia |
|---|---|---|---|---|---|
| Monte Gariglione Mg1,Mg2,Mg3,Mg4,Mg5 |
641756 m E 4332779 m N |
1500-1600 | Sud-Est | 8 - 10 ha | Abieti-Faggeta non gestita |
| Monte Gariglione Mg6,Mg7,Mg8,Mg9,Mg10 |
641874 m E 4332591 m N |
1500 | Sud-Est | 8 - 10 ha | Abieti-Faggeta gestita |
| Fondo Curto Fc1,Fc2,Fc3,Fc4,Fc5 |
640075 m E 4333190 m N |
1500 | Nord-Est | 8 - 10 ha | Faggeta non gestita |
| Fondo Curto Fc6,Fc7,Fc8,Fc9,Fc10 |
639923 m E 4333198 m N |
1500 | Nord-Ovest | 8 - 10 ha | Faggeta gestita |
| Vallone Cecita Vc1,Vc2,Vc3,Vc4,Vc5 |
633228 m E 4361320 m N |
1180-1250 | Sud | 10 - 15 ha | Pineta non gestita, suolo acclive e sabbioso |
| Vallone Cecita Vc6,Vc7,Vc8,Vc9,Vc10 |
635085 m E 4361009 m N |
1200 | Sud | 10 - 15 ha | Pineta gestita, suolo con strato umifero sviluppato |
Bosco misto faggio-abete di Monte Gariglione
Monte Gariglione (1765 m s.l.m.) è la cima più elevata della Sila Piccola ed è compreso nella Foresta del Gariglione che interessa prevalentemente i versanti in destra idrografica della valle del fiume Tacina, con macro-esposizione nord e, in minor misura, quelli in sinistra idrografica del bacino del Rio Gallina, un affluente del fiume Soleo. La foresta del Gariglione fu definita da Douglas ([14]) “un vero e proprio Urwald […], un impenetrabile groviglio di alberi […]” e appariva al visitatore come una foresta primigenia, mai sfiorata dalla mano dell’uomo. Nel 1907 la Foresta del Gariglione fu venduta alla Rueping, una società austro-tedesca, che iniziò l’utilizzazione del bosco fino alla prima guerra mondiale. Dopo il conflitto i lavori di utilizzazione furono ripresi dalla Società Forestale del Mezzogiorno d’Italia (So. Fo.Me. - [25], [13]). Il bosco del Gariglione è stato trattato con taglio raso e rilascio di riserve. Nel ventennio 1929-1949 l’utilizzazione forestale ha interessato circa il 90% della massa legnosa ([11]). Nonostante le utilizzazioni intense, il bosco Gariglione ha dimostrato una eccezionale capacità di recupero dovuta alle condizioni ecologiche favorevoli e oggi si presenta come una delle biocenosi meglio ripristinate dell’altopiano silano. Il bosco con caratteri di vetustà dove sono stati condotti i rilievi interessa un’area non particolarmente ampia risparmiata al taglio per la sua posizione immediatamente a monte dell’edificio dove avevano sede gli Uffici della So.Fo.Me., in seguito adibito, nella seconda metà del secolo scorso, a caserma del Corpo Forestale dello Stato. Si tratta di un bosco a struttura disetanea, pluristratificato, caratterizzato da piante di notevoli dimensioni di abete bianco e faggio, spesso nella fase di senescenza, e da gruppi misti di faggio e abete di diverse età, con elevata densità e grado di copertura. L’abete e il faggio raggiungono dimensioni massime, rispettivamente di 115 cm e 88 cm di diametro e rispettivamente 31.5 m e 29.5 m di altezza. In genere si nota un buon accumulo di lettiera al suolo, benché i processi di decomposizione procedano abbastanza rapidamente formando un suolo piuttosto soffice e ricco di sostanza organica. I boschi più o meno regolarmente sottoposti a interventi colturali di Monte Gariglione sono invece costituiti da cenosi miste faggio-abete, a struttura monoplana o bistratificata. Le ultime utilizzazioni risalgono alla fine degli anni sessanta del secolo scorso. È stato realizzato un intervento di forte intensità per cui si rilevano piante di notevoli dimensioni di faggio e di abete bianco sparse in modo irregolare e una fustaia monoplana che si è sviluppata in seguito all’ultima utilizzazione, con locale prevalenza dell’una o dell’altra specie. Questi soprassuoli si possono inquadrare nella fitocenosi Ranunculo-Fagetum sylvaticae (Bonin 1967). Il bosco è riferibile all’habitat d’interesse comunitario 9220* Faggeti degli Appennini con Abies alba Miller e faggeti con Abies nebrodensis (Lojac.) (Mattei, 1908). Il sottobosco è ricco di specie orofile a distribuzione meridionale quali Campanula trichocalycina Ten., Calamintha grandiflora (L.), Neottia nidus-avis (L.) Rich, Sorbus aucuparia L., Cardamine bulbifera (L.) Crantz. L’area è ricca anche di habitat rivulari ricchi di fitocenosi inquadrate nelle classi dei Galio aparines-Urticetea dioicae (Passarge ex Kopecký 1969) e dei Montio fontanae-Cardaminetea amarae (Br.-Bl. & Tüxen ex Klika & Hadac 1944); particolarmente importanti sono le comunità caratterizzate dalla presenza di Lereschia thomasii (Ten.) Boiss. e Soldanella calabrella Kress. Per l’entomofauna saproxilobionte significativa è la presenza di Cucujus cinnaberinus (Scopoli, 1763), Acanthocinus xanthoneurus (Mulsant & Rey, 1852), Sinodendron cylindricum (Linné, 1758) e Clinidium canaliculatum Costa, 1839.
Faggeta di Fondo Curto
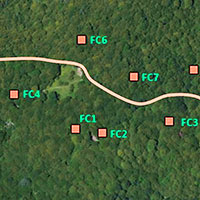
Il bosco si trova sul versante settentrionale della Foresta del Gariglione, nel tratto compreso tra la vetta di Cugno del Porazzo (1650 m s.l.m.), e il fiume Tacina (Fig. 2); l’area è posta a una quota di circa 1500 m s.l.m.; le pendenze non sono particolarmente elevate. Il soprassuolo con caratteri di vetustàè costituito da una faggeta con piante di diversa età in buone/ottime condizioni vegetative. Le caratteristiche di vetustà di questi popolamenti sono legate alla presenza di piante di dimensioni elevate - di cui la maggiore misura 113 cm di diametro e 34 m di altezza - e all’assenza di interventi colturali che ne hanno potuto condizionare le dinamiche evolutive. In alcuni tratti predomina la fustaia monoplana, con piante di faggio di notevoli dimensioni, sparse o a gruppi, rilasciate in seguito alle ultime utilizzazioni effettuate dalla So.Fo.Me. nel periodo 1929-1949. Successivamente non è stato effettuato nessun intervento. In altri casi la struttura è tendenzialmente pluristratificata, in quanto il soprassuolo è composto da piante vetuste rilasciate in modo sporadico, in parte da piante insediatisi dopo l’ultima utilizzazione e infine da piante nello stadio di perticaia o giovane fustaia. Sono sporadicamente presenti anche esemplari di pioppo tremulo e, nelle esposizioni più calde, di pino nero di Calabria. Sotto copertura è presente uno strato non eccessivamente denso di lettiera costituita in gran parte dalle foglie dell’anno. Il suolo è decisamente soffice, ricco di humus e gli orizzonti superficiali sono di colore scuro. Per quanto riguarda l’entomofauna saproxilobionte si segnala la presenza Acanthocinus xanthoneurus (Mulsant & Rey, 1852), Sinodendron cylindricum (Linné, 1758). Per i coleotteri carabidi è nota la presenza di Pterostichus ruffoi Sciaky, 1986, pterostichino endemico della Calabria. La faggeta è riferibile all’habitat d’interesse comunitario 9220* Faggeti degli Appennini con Abies alba Miller e faggeti con Abies nebrodensis (Lojac.) Mattei, 1908.
Fig. 2 - Foto aerea del Bosco Fondo Curto. Localizzazione delle sub-stazioni.
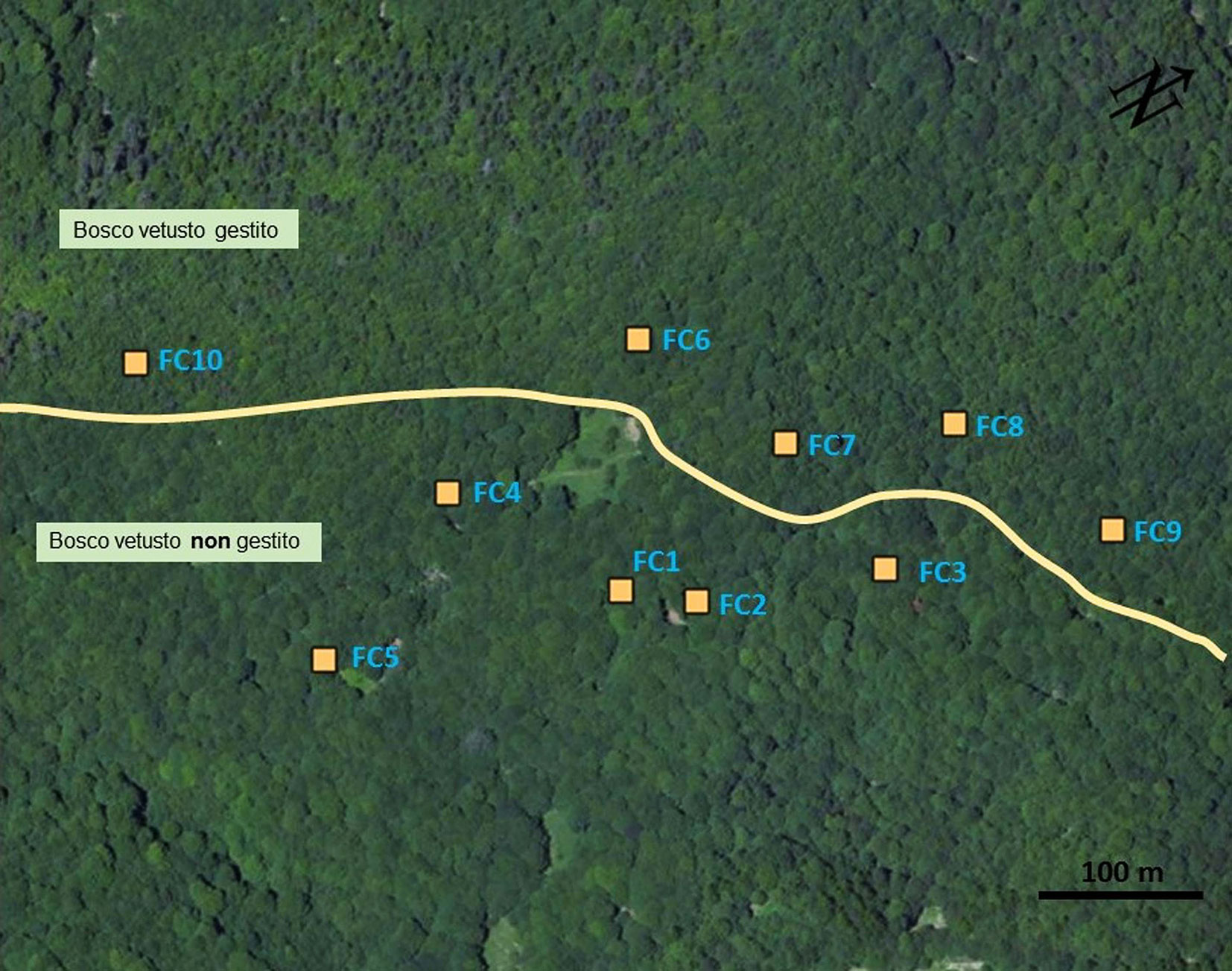
La faggeta sottoposta a interventi selvicolturali ha invece una struttura generalmente monoplana e un elevato grado di copertura. Le ultime utilizzazioni risalgono alla fine degli anni sessanta del secolo scorso. Si rilevano piante vetuste di faggio rilasciate sporadicamente che sovrastano la fustaia monoplana che si è sviluppata in seguito all’ultima utilizzazione.
Pineta di Vallone Cecita
Il bosco si trova sui versanti in destra idrografica del Vallone del Torrente Cecita e ricade nella Foresta Demaniale della Fossiata, in Sila Grande. Si estende a una quota media di 1200 m s.l.m., con una macro-esposizione a sud. Le pendenze sono in genere molto elevate, favorendo in una stazione fenomeni di erosione del suolo aggravati da un eccessivo calpestio da parte di visitatori (Vc3). Il bosco con caratteri di vetustàè una pineta disetanea costituita da esemplari di pino laricio di dimensioni ed età elevate con diametri fino a 190 cm, altezze che raggiungono 40 m ed età superiori ai 300 anni ([12]); vi sono gruppi di piante di differente età, posti spesso in prossimità degli esemplari più vecchi, ma al loro interno tendenzialmente coetaneiformi e distribuiti su superfici piuttosto limitate. Il profilo verticale è piuttosto articolato e il grado di copertura elevato. Nelle zone prive di copertura arborea o dove la densitàè scarsa sono presenti le specie caratteristiche dell’associazione Hypochoerido-Pinetum calabricae (Bonin 1978) quali Hypochoeris laevigata (L.) Ces., Pass. & Gibelli, Brachypodium sylvaticum (Huds.) P. Beauv., Festuca heterophylla Lam., Luzula forsteri (Sm.) DC., Melica uniflora Retz. La presenza delle specie arboree, erbacee e arbustive consente di attribuire questi popolamenti all’habitat d’interesse comunitario 9530* Pinete sub-mediterranee di pini neri endemici. Le caratteristiche di vetustà sono legate alla presenza di piante di pino nero di Calabria di grandi dimensioni e di età avanzata, distribuite in modo decisamente irregolare su tutta l’area, dall’assenza di interventi di utilizzazione a partire dalla prima metà del secolo scorso e dalla presenza di dinamiche evolutive non influenzate dall’attività antropica. Da segnalare la presenza di abbondante coleotterofauna saproxilobionte come: Cucujus cinnaberinus (Scopoli, 1763), Cucujus haematodes Erichson, 1845, Clinidium canaliculatum Costa, 1839, Chalcophora intermedia intermedia (Rey, 1890), Ergates faber opifex Mulsant, 1851, Prionus coriarius (Linné, 1758). La pineta sottoposta a interventi selvicolturali tra gli anni trenta e cinquanta del secolo scorso presenta oggi una struttura generalmente monoplana e una densità elevata. Localmente diviene bistratificata per la presenza di piante vetuste isolate e sporadiche di pino laricio all’interno della fustaia. È presente anche una discreta presenza di lettiera, che tuttavia non raggiunge mai spessori elevati. Il suolo è soffice e gli orizzonti più superficiali presentano un colore scuro.
Campionamento dei Carabidi
All’interno di ciascuno dei 6 boschi individuati sono stati scelti in maniera casuale cinque sub-plot di 400 m2 di superficie (20 × 20 m) in cui sono state installate due trappole a caduta, per un totale di 10 trappole per ogni bosco (Fig. 2). La trappola a caduta consiste in un bicchiere di plastica di 11 cm di profondità e 8.5 cm di diametro, innescata con una soluzione di aceto satura di sale e alloggiata a sfioramento nel terreno ([8]). Il campionamento è stato effettuato periodicamente ogni 20-25 giorni da giugno ad ottobre 2014. I coleotteri carabidi raccolti sono stati identificati a livello di specie e conservati in una soluzione alcolica al 60% o a secco in scatole entomologiche.
Classificazione ecologica
Le specie sono state classificate in relazione a dimensioni, dieta degli adulti, capacità di dispersione (conformazione alare), corologia e affinità con l’habitat (Tab. 2). Le dimensioni corporee delle specie trovate, ricavate da Porta ([39]), Brandmayr et al. ([8]), Mazzei et al. ([30]) e Mazzei et al. ([31]), sono state assegnate a sette classi dimensionali di 5 mm (da 0-5 mm a 30 mm) e integrate da misurazioni dirette di individui di entrambi i sessi. Quattro categorie di specializzazione alimentare, come definite da Brandmayr et al. ([8]), Mazzei et al. ([31]) e Talarico et al. ([45]) in zoofaga specializzata, zoofaga, spermofaga e onnivora, sono state integrate sulla base delle informazioni ricavate dalla banca dati a disposizione del gruppo di ricerca (una collezione entomologica integrata da matrici specie/stazione inedite). La capacità di dispersione è stata distinta in tre classi (brachitteri, pteridimorfi, e macrotteri), ricavate da Brandmayr ([6]) e Brandmayr et al. ([8]). Inoltre, in fase di determinazione delle specie è stata comunque verificata la conformazione alare di molti esemplari. La corologia è stata invece caratterizzata tramite 6 parametri, quali: endemiche calabresi, endemiche Italiane, Europee, Mediterranee, Euroasiatiche o -sibiriche e Paleartiche ([8]). Le preferenze ecologiche nei confronti dell’habitat sono infine state attribuite nel seguente modo: specie forestali, di ambienti aperti ed euritope, così come riportato in molti lavori sui coleotteri carabidi. Per molti taxa mediterranei poco noti le definizioni sono state verificate grazie alla già ricordata banca dati.
Tab. 2 - Quadro riassuntivo delle caratteristiche adattative delle specie come distribuzione percentuale nelle aree forestali campionate. (NG): non gestito; (G): gestito.
| Caratteristiche biologiche |
Codice numerico |
Categorie degli attributi |
Mg 1-5 (NG) |
Mg 6-10 (G) |
Fc 1-5 (NG) |
Fc 6-10 (G) |
Vc 1-5 (NG) |
Vc 6-10 (G) |
||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | n | % | n | % | n | % | |||
| Dimensioni | 1 | ≤ 5 mm | 1 | 7.7 | 2 | 12.5 | 1 | 7.1 | 1 | 9.1 | - | - | - | - |
| 2 | > 5 - ≤ 10 | 3 | 23.1 | 4 | 25.0 | 4 | 28.6 | 4 | 36.4 | 5 | 45.5 | 5 | 45.5 | |
| 3 | > 10 - ≤ 15 | 3 | 23.1 | 4 | 25.0 | 3 | 21.4 | 3 | 27.3 | - | - | - | - | |
| 4 | > 15 - ≤ 20 | 3 | 23.1 | 3 | 18.8 | 3 | 21.4 | 2 | 18.2 | 3 | 27.3 | 3 | 27.3 | |
| 5 | > 20 - ≤ 25 | 2 | 15.4 | 2 | 12.5 | 2 | 14.3 | 1 | 9.1 | 2 | 18.2 | 2 | 18.2 | |
| 6 | > 25 - ≤ 30 | 1 | 7.7 | 1 | 6.3 | 1 | 7.1 | - | - | 1 | 9.1 | 1 | 9.1 | |
| 7 | > 30 mm | - | - | - | - | - | - | - | - | - | - | - | - | |
| Alimentazione adulti |
1 | Zoofaga specializzata | 1 | 7.7 | 2 | 12.5 | 1 | 7.1 | - | - | 2 | 18.2 | 2 | 18.2 |
| 2 | Zoofaga | 11 | 84.6 | 12 | 75.0 | 13 | 92.9 | 10 | 90.9 | 7 | 63.6 | 7 | 63.6 | |
| 3 | Spermofaga | - | - | - | - | - | - | - | - | - | 0.0 | - | - | |
| 4 | Onnivora | 1 | 7.7 | 2 | 12.5 | - | - | 1 | 9.1 | 2 | 18.2 | 2 | 18.2 | |
| Conformazione alare |
1 | Brachitteri | 11 | 84.6 | 11 | 68.8 | 12 | 85.7 | 8 | 72.7 | 7 | 63.6 | 7 | 63.6 |
| 2 | Pteridimorfi | 1 | 7.7 | 1 | 6.3 | - | - | 1 | 9.1 | - | - | 1 | 9.1 | |
| 3 | Macrotteri | 1 | 7.7 | 4 | 25.0 | 2 | 14.3 | 2 | 18.2 | 4 | 36.4 | 3 | 27.3 | |
| Corologia | 1 | Endemiche calabrese | 1 | 7.7 | 1 | 6.3 | 1 | 7.1 | 1 | 9.1 | - | - | - | - |
| 2 | Endemiche Italiane | 7 | 53.8 | 7 | 43.8 | 8 | 57.1 | 5 | 45.5 | 3 | 27.3 | 3 | 27.3 | |
| 3 | Europee | 4 | 30.8 | 4 | 25.0 | 3 | 21.4 | 3 | 27.3 | 3 | 27.3 | 4 | 36.4 | |
| 4 | Mediterranee | 1 | 7.7 | 1 | 6.3 | 1 | 7.1 | - | - | 1 | 9.1 | 1 | 9.1 | |
| 5 | Euroasiatiche-sibiriche | - | - | 2 | 12.5 | 1 | 7.1 | 2 | 18.2 | 4 | 36.4 | 3 | 27.3 | |
| 6 | Paleartiche | - | - | 1 | 6.3 | - | - | - | - | - | - | - | - | |
| Habitat | 1 | Ambienti forestali | 11 | 84.6 | 11 | 68.8 | 11 | 78.6 | 8 | 72.7 | 8 | 72.7 | 7 | 63.6 |
| 2 | Eurecia | 2 | 15.4 | 4 | 25.0 | 3 | 21.4 | 2 | 18.2 | 1 | 9.1 | 2 | 18.2 | |
| 3 | Ambienti aperti | - | - | 1 | 6.3 | - | - | 1 | 9.1 | 2 | 18.2 | 2 | 18.2 | |
| Numero di specie totale | 13 | - | 16 | - | 14 | - | 11 | - | 11 | - | 11 | - | ||
Analisi statistiche
Le analisi statistiche sono state elaborate con il software STATISTICA® (StatSoft, Tulsa, OK, USA). I dati di cattura delle singole specie sono espressi in Densità di Attività (DA), ovvero il rapporto tra il numero di individui (nij) di una determinata specie catturati in ogni periodo di raccolta per l’unità di sforzo (us) mensile di campionamento (numero delle trappole attive per i giorni di permanenza delle stesse riferito all’unità di tempo di dieci giorni). Questo rappresenta il numero medio di individui attivi di una specie catturati in una trappola, riferito all’unità di tempo di dieci giorni. La DA, calcolata per ogni singola data di raccolta, corrisponde alla densità di popolazione attiva in un dato momento della stagione ([8]). Mentre la DAa (Densità Attività annua) è il rapporto tra il numero complessivo di individui (n.j) di un’unica specie per l’unità di sforzo totale (US, sommatoria delle us mensili). La matrice dei dati costruita con i valori di abbondanza delle specie è stata analizzata mediante l’Analisi delle Corrispondenze Multiple (MCA). Per analizzare l’eterogeneità dei dati è stata eseguita la Cluster Analysis. I siti di campionamento sono stati confrontati con il metodo della minima varianza (metodo di Ward) per la stima della similarità fra coppie di campioni tenendo conto soltanto dei dati di presenza/assenza. Le specie sono state infine raggruppate mediante il metodo legame completo con il calcolo della distanza euclidea.
Risultati
Nel corso del campionamento sono stati catturati complessivamente 6654 individui ripartiti su 23 specie di coleotteri carabidi (Tab. S1 - Materiale supplementare). Nel bosco non gestito e in quello gestito della foresta del Monte Gariglione sono state rinvenute rispettivamente 13 e 16 specie, mentre 14 e 11 nella foresta di Fondo Curto; nella foresta di Vallone Cecita sono invece state catturate 11 specie in entrambe le tipologie gestionali.
Caratteristiche funzionali delle comunità
Vengono di seguito illustrate le caratteristiche funzionali delle comunità campionate riportate come percentuale delle specie che presentano una data caratteristica rispetto al totale delle specie che compongono la comunità (Tab. 2).
Dimensioni. Nell’area forestale del Monte Gariglione il passaggio al bosco gestito comporta un lieve aumento della presenza di specie dalle dimensioni medio-piccole. Stesso andamento si osserva nell’area forestale di Fondo Curto, dove un carabide altamente specializzato, Carabus lefebvrei bayardi Solier, 1835, lungo circa 22-25 mm, scompare nel bosco gestito. Non ci sono effetti significativi fra bosco gestito e non gestito nell’area forestale di Vallone Cecita.
Caratteristiche alimentari degli adulti. Nel bosco gestito di Monte Gariglione si osserva la comparsa di specie opportuniste come Amara aulica (Panzer, 1796), specie con dieta onnivora in grado di nutrirsi sia di prede che di semi. Compare anche Badister bullatus (Schrank, 1798), con probabile specializzazione alimentare elicofaga, cioè a base di chiocciole. Nel bosco gestito di Fondo Curto si osserva la scomparsa di Cychrus italicus Bonelli, 1810 predatore elicofago, mentre compare l’onnivoro e opportunista Harpalus rufipalpis rufipalpis Sturm, 1818. Anche in questo caso nell’area forestale del Vallone Cecita non si evidenziano cambiamenti significativi della composizione faunistica.
Capacità di dispersione. Nell’area forestale del Monte Gariglione e di Fondo Curto la gestione del bosco comporta un notevole aumento delle specie macrottere, cioè in grado di volare. Nella foresta di Vallone Cecita in entrambe le tipologie di foresta si osserva lo stesso rapporto fra volatori potenziali e non, sebbene compaia lo pteridimorfo Harpalus rufipalpis Sturm, 1818 del quale sono note sia le forme brachittere che macrottere.
Spettro corologico. Nell’area forestale del Monte Gariglione il bosco gestito mostra un deciso aumento delle specie ad ampio spettro corologico come Amara aulica (Panzer, 1796) e Badister bullatus (Schrank, 1798). A Fondo Curto si verifica la medesima tendenza, con la scomparsa dal bosco gestito di Carabus lefebvrei bayardi Solier, 1835 e Platyderus neapolitanus neapolitanus (Reiche, 1855), specie endemiche dell’Appenino italiano. Nell’area forestale del Vallone Cecita non si osservano effetti così eclatanti, anzi la foresta gestita registra la scomparsa di Ophonus rufibarbis (Fabricius, 1792), specie di formazioni aperte a distribuzione Turanico-Europeo-Mediterraneo, e la comparsa di Abax parallelepipedus curtulus Fairmaire, 1856, un elemento a distribuzione Europea in Calabria presente esclusivamente in aree boschive del piano montano.
Preferenze ambientali. Sia sul Monte Gariglione che a Fondo Curto il bosco gestito mostra un deciso aumento delle specie di ambienti aperti, come ad esempio Amara aulica (Panzer, 1796) esclusiva del piano montano, e un aumento delle specie euriecie; al contrario mancano tipiche specie forestali come Carabus lefebvrei bayardi Solier, 1835 e Cychrus italicus Bonelli, 1810. Nell’area forestale del Vallone Cecita le differenze fra le due tipologie gestionali sono invece molto meno marcate.
Caratterizzazione delle comunità
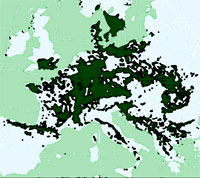
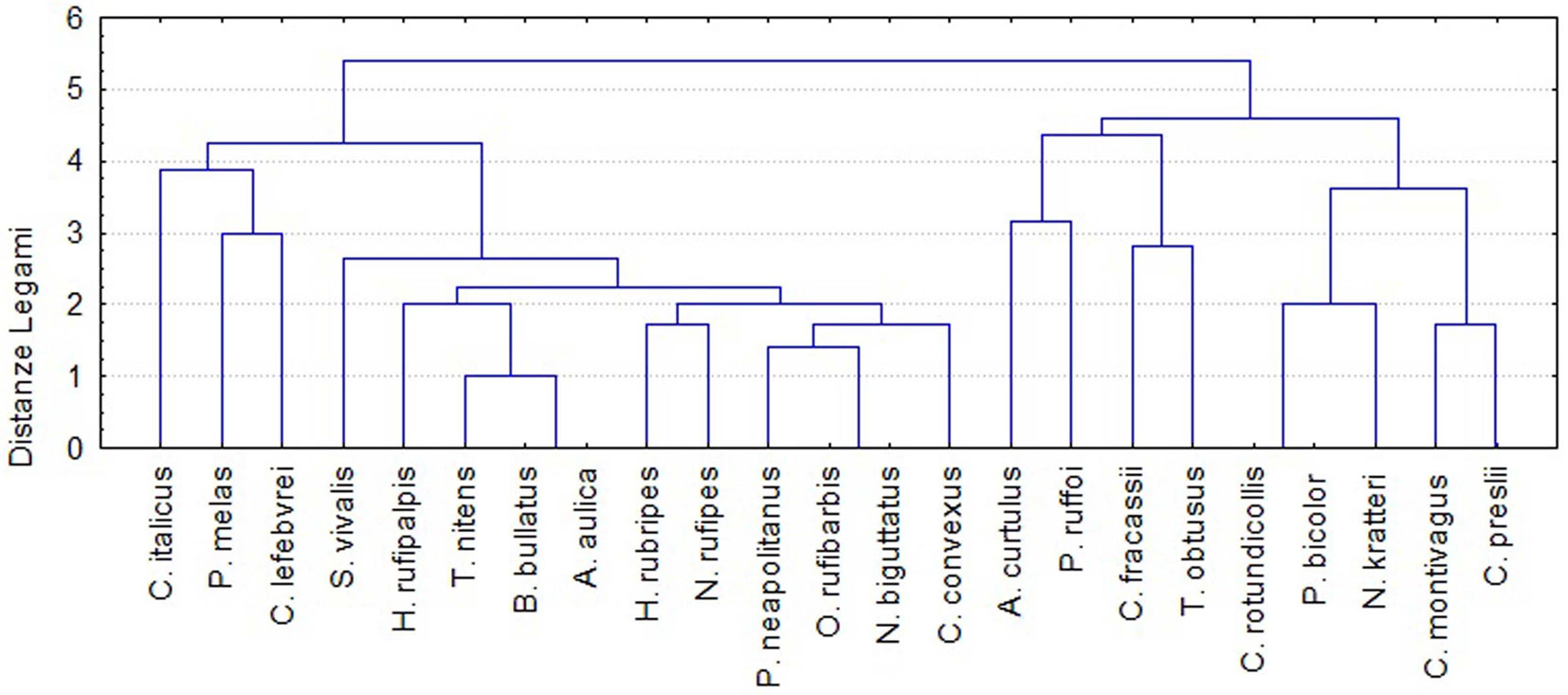
Nelle tre aree forestali campionate albergano due comunità ben diversificate, una legata alle faggete “fredde” e agli abieto-faggeti (Ranunculo brutii-Fagetum sylvaticae Bonin 1967), l’altra più termofila e tipica di minori altitudini che caratterizza le pinete. La prima comunità presenta sei specie igrofilo-microterme esclusive e fortemente caratterizzanti, quali Pterostichus bicolor amorei Ganglbauer, 1907, Calathus rotundicollis Dejean, 1828, Trechus obtusus lucanus Focarile, 1949, Pterostichus ruffoi Sciaky, 1986, Calathus fracassii Heyden, 1908 e Trichotichnus nitens (Heer, 1838), già individuate come caratteristiche di faggete appenniniche “fredde” ([5], [7]). Le pinete sono invece caratterizzate quantitativamente da una maggior abbondanza di tre carabidi più termofili Cychrus italicus Bonelli,1810 e Carabus lefebvrei bayardi Solier, 1835 entrambi predatori di chiocciole, e Pterostichus melas italicus (Dejean, 1828), nonché da due elementi in comune con i querceti e più frequenti in formazioni aperte, come Notiophilus rufipes Curtis, 1829 e Harpalus rubripes (Duftschmid, 1812). La classificazione ecologica delle specie in base alla loro abbondanza nelle 30 sub-stazioni campionate permette di distinguere abbastanza nettamente due gruppi (Fig. 3). Il primo comprende molte specie forestali termofile o mesofile, accanto a elementi delle formazioni aperte più frequenti nelle pinete grazie al loro maggiore soleggiamento. Il secondo include elementi pedologicamente più esigenti, igrofili e/o microtermi.
Fig. 3 - Dendrogramma di similarità tra le comunità di Carabidi (legame completo, distanze euclidee).
Classificazione delle comunità campionate
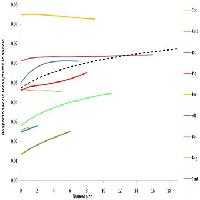
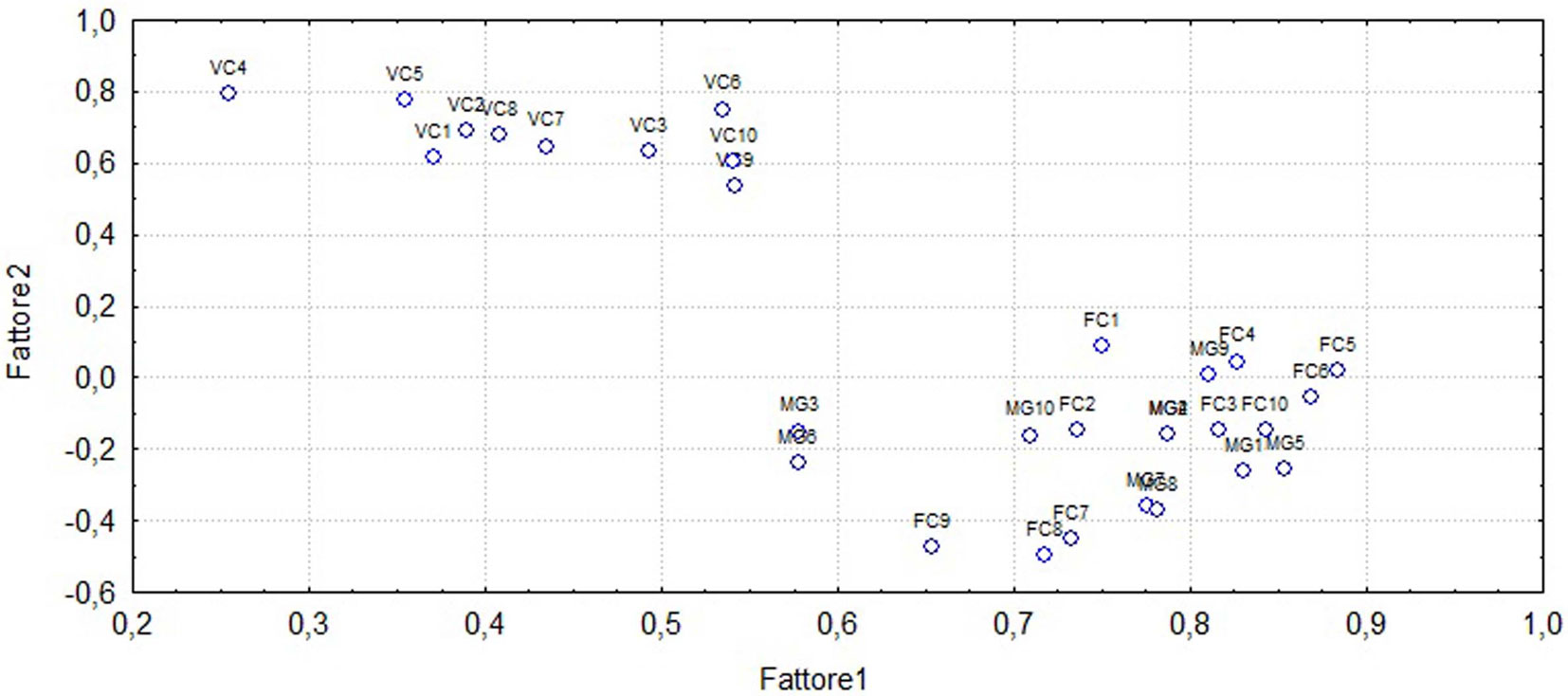
L’analisi PCA ha permesso di individuare due gruppi di stazioni (Fig. 4). Un primo gruppo è rappresentato dalle stazioni di Vallone Cecita che si separano dal secondo gruppo rappresentato invece dalle stazioni di Fondo Curto e di Monte Gariglione. Sulla base della tipologia forestale è quindi possibile affermare che complessivamente la distribuzione delle specie di Carabidi non è casuale, ma è influenzata dalle preferenze ambientali che hanno permesso di rilevare somiglianze fra faggete e differenze tra faggete e pinete.
Fig. 4 - Ordinamento della posizione dei siti lungo il primo e secondo asse della PCA. Al primo autovalore corrisponde il 46% della variabilità dei dati, mentre al secondo corrisponde il 20%, che sommati descrivono il 66% dell’informazione strutturale contenuta nei dati di partenza.
Analisi attraverso gli indici di similarità
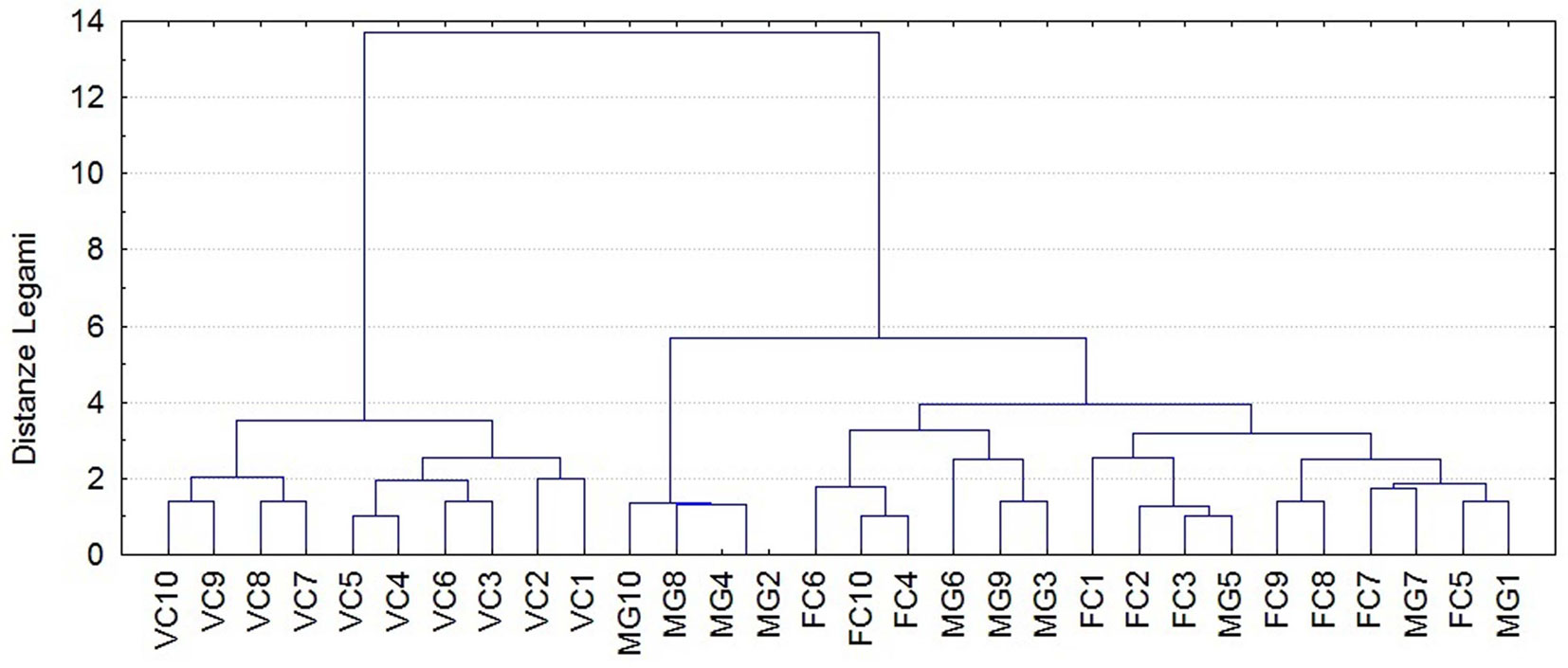
L’analisi multivariata delle strutture delle specie rinvenute (Tab. S1 - Materiale supplementare) mostra una struttura eterogenea (non continua) delineata su base strettamente numerica (Fig. 5), fermo restando che i valori delle DAa (Densità Attività annua) analizzati dall’analisi multivariata derivano dalle interazioni tra variabili biotiche e abiotiche. Ancora una volta l’insieme dei siti risulta aggregato principalmente in due gruppi, di cui uno è composto dalle pinete e l’altro le faggete e gli abieto-faggeti. Anche questa analisi conferma quindi quanto già evidenziato dalla PCA, e cioè che le pinete ospitano una comunità ben distinta da quella della faggeta o del consorzio misto abete-faggio.
Fig. 5 - Dendrogramma di classificazione dei siti campionati (metodo di Ward, distanze euclidee).
Discussione
Le tre aree forestali qui studiate rappresentano le tipologie forestali più diffuse sull’altipiano silano ([26], [33]). La diversa gestione a cui sono sottoposte e la composizione presentata hanno una ricaduta sull’entomofauna, in questo caso del suolo, che è direttamente verificabile. I risultati di questo studio dimostrano infatti che fra pinete, tendenzialmente termofile, e faggete e consorzi misti abete-faggio, formazioni forestali più fresche, esistono notevoli differenze nella comunità di specie e nella densità delle loro popolazioni. Le differenze osservate, già in parte note per l’Appennino ([5], [7]), inducono a trattare gli effetti della gestione del bosco rispetto alle condizioni più o meno prossime alla sua “vetustà” separatamente per ogni tipologia forestale. Le foreste poste a quote più elevate sono infatti caratterizzate dalla presenza esclusiva di un gruppo di specie igrofile e/o microterme (ad es., Pterostichus bicolor amorei Ganglbauer, 1907,Calathus fracassii Heyden, 1908, Calathus rotundicollis Dejean, 1828), in parte legate anche a suoli umidi (Nebria kratteri Dejean & Boisduval, 1830) o anche intrisi d’acqua (Pterostichus ruffoi Sciaky, 1986). Alcune di queste specie, come ad esempio Pterostichus bicolor amorei Ganglbauer, 1907 e Pterostichus ruffoi Sciaky, 1986, mostrano densità decisamente minori nei boschi gestiti rispetto a quelli non gestiti. Altre specie, come Nebria kratteri Dejean & Boisduval, 1830, mostrano invece decise preferenze per suoli più umidi indipendentemente dalla gestione forestale. L’analisi della biodiversità sembra dare risultati eterogenei se non addirittura discordanti fra loro. Al Gariglione si passa da una media di 8.6 specie per sub-stazione nei boschi non gestiti a 8.8 in quelli gestiti; parimenti nelle faggete di Fondo Curto si hanno valori rispettivamente di 9.8 e 7.6; nelle pinete di Vallone Cecita la densitàè invece di 6.6 e 6.4. Anche se il numero di repliche è esiguo, è chiaro che il numero di specie non sembra essere di per sé discriminante del tipo di gestione. Esso dipende, infatti, anche dalle condizioni di conservazione del suolo, che è spesso determinante per la diversità dei Carabidi. Non è quindi un caso che la stazione V3 di Vallone Cecita, caratterizzata da pendenze elevate e suolo degradato anche a seguito di un accumulo di materiale estraneo alla stazione, presenti una comunità di sole 4 specie (Tab. 2). La carabidofauna osservata presenta invece due comunità ben separate, quella dominata dal faggio e da Pterostichus bicolor amorei Ganglbauer, 1907, e quella delle pinete composta dagli elementi più termofili. L’asse orizzontale dell’analisi PCA rappresenta certamente un gradiente di crescente microtermia (da sinistra a destra), mentre il secondo asse è piuttosto da interpretarsi come un gradiente pedologico, ma anche di ombreggiamento del suolo. Spiccano qui le sub-stazioni di Fondo Curto sottoposte a interventi selvicolturali, ma situate sul versante Nord in situazione di minimo soleggiamento. L’analisi della composizione e delle caratteristiche delle comunità, per quanto indispensabili, sembrano però meno efficaci nella discriminazione degli effetti della gestione sulla comunità. Le caratteristiche funzionali esaminate permettono invece un certo grado di generalizzazione dei fenomeni osservati. Nelle comunità di faggete “fredde” e negli abieto-faggeti posti a quote elevate della Sila, la gestione del bosco tende infatti a spostare lo spettro dimensionale delle specie verso classi inferiori, comprese fra 5 e 15 mm di lunghezza corporea. Compaiono specie onnivore e aumentano tendenzialmente i predatori generici, crescono anche le specie in grado di volare, cioè con una maggiore capacità di dispersione. Aumentano vistosamente anche gli elementi a distribuzione geografica più ampia, euroasiatica, eurosibirica o paleartica. Tendono infine ad aumentare i carabidi di ambienti aperti, legati a pascoli o coltivi. Nelle pinete invece quasi tutte queste differenze sono più sfumate o del tutto inesistenti, mentre assume notevole importanza il grado di conservazione del suolo. Dove questo è sufficientemente evoluto può ospitare anche specie forestali esigenti, come Abax parallelepipedus curtulus Fairmaire, 1856 le cui larve lumbricifaghe prediligono suoli non troppo acidificati e ricchi di humus. Se invece le condizioni pedologiche sono caratterizzate da fenomeni di degrado o di più o meno accentuata erosione, specie più euriecie o comunque tipiche di boschi gestiti possono comparire anche in situazioni di vetustà del soprassuolo.
Conclusioni
Le foreste calabresi, e in particolare quelle della Sila, sono state per secoli interessate da utilizzazioni più o meno intense. In epoca romana il legname ricavato dallo sfruttamento di questi boschi era necessario per la costruzione delle navi e degli edifici pubblici. Utilizzazioni intense sono proseguite durante il Medioevo e il Rinascimento fino alla fine degli anni ’50 del secolo scorso, provocando una semplificazione della struttura e della composizione specifica dei boschi ([18]). La conservazione della biodiversità forestale rappresenta il fulcro delle politiche di tutela della natura, in quanto le foreste sono tra gli ecosistemi terrestri a maggior contenuto di diversità genetica, specifica e sistemica.
Gli studi dei boschi vetusti (o con caratteri di vetustà) consentono di integrare varie competenze disciplinari, quali quelle forestali, botaniche e zoologiche, con ricerche inerenti la struttura e la funzionalità dei sistemi biologici complessi da una parte e la valutazione quali-quantitativa delle funzioni ecosistemiche dall’altra. Questa ricerca ha messo in evidenza come la diversità funzionale dei coleotteri carabidi sia influenzata dalla gestione forestale, in particolare da modelli funzionali differenti e più articolati in sistemi forestali caratterizzati da elevata complessità strutturale rispetto a quelli semplificati come conseguenza dell’applicazione di interventi selvicolturali che fanno riferimento alla selvicoltura classica. Nei primi si osserva una dominanza di specie di maggiori dimensioni, una maggior specializzazione alimentare zoofaga, un basso potere di dispersione, una distribuzione geografica più ristretta e tendente all’endemicità, ma soprattutto un maggior numero di specie prettamente forestali. I risultati, anche se limitati all’altopiano della Sila, offrono un ulteriore contributo per la caratterizzazione delle foreste vetuste dell’Appennino meridionale. I risultati ottenuti sono pienamente concordanti con quanto già osservato in un confronto fra due ontanete del Parco Nazionale della Sila, di cui una con caratteri di vetustà e l’altra sottoposta a regolare ceduazione ([31]). Le pinete sinora studiate non hanno offerto dati di facile interpretazione, probabilmente perché le sub-stazioni campionate sono molto eterogenee e poco comparabili dal punto di vista pedologico. Complessivamente i risultati conseguiti sono tuttavia esaustivi almeno per i sistemi forestali della faggeta pura o mista con abete bianco. Altre situazioni già studiate in Calabria, come le aree caratterizzate da soprassuoli vetusti o con caratteri di vetustà, pur non essendo esenti dagli effetti dell’azione dell’uomo e quindi non essendo espressione della sola evoluzione naturale, si trovano in una condizione di maggiore ricchezza di “elementi di vetustà”. Infatti non sono state raccolte specie strettamente dendrofile o con tendenze dendrofile ([8]) come Leistus sardous (Baudi di Selve, 1883), Leistus spinibarbis (Fabricius, 1775) e certi sfodrini (Laemostenus venustus Dejean, 1828) già segnalati in castagneti secolari ([37]) e forse utilizzabili anche in Sila come bioindicatori. È infine da rilevare come anche nelle particelle “gestite” un turno del bosco di oltre mezzo secolo permetta di conservare nell’attuale carabido-cenosi solo residui degli aspetti successionali delle medesime comunità. Un periodo di sessant’anni sembra dunque favorire in modo soddisfacente il recupero di una comunità entomologica edafica almeno a livello di subclimax.
Ringraziamenti
Il presente lavoro di ricerca si inserisce nell’Azione di Programma “Foreste Vetuste nei Parchi Nazionali italiani” promosso dal Ministero dell’Ambiente e della Tutela del Territorio e del Mare.
References
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar