Short and mid-term effects of artificial defoliation on the growth of euramerican poplar (Populus × canadensis clone I-214) in poplar stands in relation to the intensity and seasonal timing of defoliation.
Forest@ - Journal of Silviculture and Forest Ecology, Volume 14, Pages 206-217 (2017)
doi: https://doi.org/10.3832/efor2417-014
Published: Aug 01, 2017 - Copyright © 2017 SISEF
Research Articles
Abstract
The impact of artificial defoliations on the growth of euramerican poplar (Populus × canadensis clone I-214) was investigated in three field assays. Young poplar trees (1st-3rd cultivation year) were subjected to 25%, 50%, 75% and 100% defoliation either in the early or in the late growing season (or in both periods), and their growth was measured in the same year and during the following 2-3 years. Only trees treated to 75% and 100% defoliation exhibited a significant growth decrease compared to control trees: the diametric growth of early defoliated trees was reduced up to 20-40% in the year of defoliation, whereas trees defoliated at the same rates late in the season, or defoliated twice in a single year, suffered the heaviest damage, showing a growth loss up to 50% in the year following the defoliation events. In the latter cases, a delayed sprouting at spring and the death of branchlets were observed. A financial analysis showed that the economic damage caused by the highest defoliation rates, mainly when occurring late in the season, definitely exceeds the cost of treatments, thus suggesting the adoption of appropriate control strategies.
Keywords
Poplar, Wood Plantation, Hybrid Clone, Artificial Defoliation, Plant Growth, Damage Treshold
Introduzione
Come è noto, la perdita di gran parte o di tutta la superficie fogliare assimilante determina in un albero una riduzione di incremento legnoso del tronco oltre che, parallelamente, una complessa serie di modificazioni fisiologiche che possono portare ad accresciuta sensibilità verso altri stress di natura biotica e abiotica ([23], [27]). Ne consegue che la pioppicoltura specializzata italiana, attività agricola finalizzata alla produzione di legno di elevata qualità per l’industria, soffre un danno economico dalla riduzione quantitativa del prodotto legnoso causata da defogliazioni di varia natura, anche se è noto che i danni economicamente più importanti patiti dalla pioppicoltura sono legati allo scadimento qualitativo del legno causato da attacchi di parassiti lignicoli e corticali ([6]).
Le defogliazioni parassitarie, in particolare quelle ad eziologia fungina, hanno condizionato l’evoluzione e la composizione clonale della pioppicoltura italiana sin dalle sue origini ([13]). L’introduzione nel nostro Paese di Venturia populina (Vuill.) Fabr., agente della defogliazione primaverile, avvenuta nella prima metà del XX Secolo, determinò il primo radicale rinnovamento clonale, con la progressiva sostituzione dei cloni “canadesi”, molto sensibili alla malattia, da parte di cloni ibridi Populus × canadensis Moench (derivanti da incroci spontanei o controllati di Populus deltoides W. Bartram ex Marshall × Populus nigra L.) resistenti. Un successivo importante rinnovamento clonale, ancor oggi in corso di evoluzione, prese avvio intorno agli anni âÂÂ60 dello stesso secolo con l’introduzione in Italia di Marssonina brunnea (Ell. et Ev.) P. Magn., agente della bronzatura, che determinò l’abbandono dei cloni ibridi più suscettibili e la loro sostituzione con cloni di P. deltoides, oppure di cloni ibridi dotati di elevata resistenza o di sufficiente tolleranza alla malattia ([12]). La riduzione di reddito annuo subita dai pioppicoltori intorno agli anni âÂÂ30 a causa della defogliazione primaverile venne stimata da Castellani & Prevosto ([11]) intorno al 60%, mentre Castellani & Cellerino ([9]) calcolarono una perdita di produzione legnosa pari al 16% della produzione nazionale annua a causa delle prime epidemie di bronzatura nei pioppeti padani. Altri temibili patogeni fogliari del pioppo sono le ruggini (Melampsora alli-populina Kleb. e Melampsora larici-populina Kleb.), che stanno anch’esse provocando danni importanti ([19]) e limitando la diffusione in coltura dei cloni più sensibili ([3]).
I pioppeti possono subire importanti defogliazioni anche da parte di insetti, in particolare Lepidotteri, che ad intervalli più o meno lunghi a seconda delle specie e in periodi specifici dell’anno a seconda del loro ciclo biologico compiono gradazioni capaci di defogliare severamente le piante ([22]). Uno di questi parassiti è la Falena invernale (Operophtera brumata L.), che compare solo sporadicamente ma può provocare forti defogliazioni su diverse specie forestali in epoca precoce, poco dopo la germogliazione. Altri temibili defogliatori Lepidotteri, in questo caso specifici delle Salicacee, sono la Pigera (Clostera anastomosis L.), che compie fino a quattro generazioni nel corso dell’anno e può provocare danni in momenti diversi della stagione vegetativa ([5]), e la farfalla bianca (Leucoma salicis L.), che compie due generazioni e può defogliare i pioppi in maggio-giugno e in agosto ([2]) (Fig. 1). Negli ultimi tre decenni la specie più dannosa è stata tuttavia la polifaga Ifantria americana (Hyphantria cunea Drury), che è giunta in Italia intorno agli anni âÂÂ70 del secolo scorso ed è stata spesso presente con elevati livelli di popolazione nei pioppeti dei cloni ibridi Populus × canadensis durante i mesi di agosto-settembre, periodo in cui le larve della seconda generazione sono in grado di provocare defogliazioni anche totali ([1]). Occorre ricordare infine il Coleottero Chrysomela populi L., capace di provocare intense defogliazioni primaverili soprattutto sui pioppi giovani. Anche se nessuno di questi parassiti entomatici, diversamente dalle malattie fungine, ha condizionato la composizione clonale della pioppicoltura italiana, i loro danni sono stati talvolta economicamente importanti, come è testimoniato da Arru che nel 1968 stimò in oltre 400.000 € (valore riportato all’attualità) il danno annualmente causato in quello stesso periodo dalla farfalla bianca alla produzione pioppicola nazionale, e da Lapietra ([24]), che per il 1981 calcolò un danno diretto annuale causato dagli insetti defogliatori (riportato all’attualità), sempre riferito all’intera produzione pioppicola nazionale, di 382.000 €, pari al 2% del totale dei danni causati da tutti gli insetti, cui andava aggiunta una spesa (sempre riportata all’attualità) per i trattamenti insetticidi di 668.000 €.
Fig. 1 - Defogliazione in pioppeto causata dal Lepidottero Leucoma salicis.

Va tuttavia sottolineato il fatto che, in mancanza di saggi condotti con un’appropriata metodologia scientifica che si avvalga di un confronto con piante testimone, la reale entità dei danni causati dagli eventi naturali di defogliazione può facilmente sfuggire alla comprensione, poiché questi eventi si presentano spesso con un livello elevato di omogeneità nelle piantagioni e con ampia estensione sul territorio, impedendo in questo modo una stima della produzione potenziale di riferimento in totale assenza di danno (testimone di riferimento).
I primi a compiere osservazioni organiche e ad eseguire rilievi comparativi sugli effetti delle defogliazioni nei pioppeti padani furono Castellani & Cellerino ([9]) per quanto riguarda le fitopatie e Arru ([5]) per quanto riguarda gli insetti. Fu proprio Arru ([5]) a definire alcuni aspetti che rimasero in seguito riferimenti fondamentali per la gestione dei pioppeti in caso di attacchi di insetti defogliatori. Essi vengono riportati di seguito con le parole stesse dell’Autore (in corsivo sono riportate alcune annotazioni personali):
- L’importanza del danno è in relazione all’intensità della defogliazione e al periodo vegetativo in cui essa si verifica.
- Attacchi ripetuti in anni successivi, o peggio ancora in uno stesso anno, aggravano seriamente i danni.
- Gli attacchi non hanno grande importanza dal punto di vista economico se determinano una sola defogliazione in un anno e se il fogliame distrutto non supera il 50%.
- Le conseguenze sulla produzione di legname [nell’anno stesso della defogliazione] sono più gravi quando la defogliazione è precoce (maggio-giugno). Tuttavia anche defogliazioni tardive (agosto) possono avere gravi conseguenze perché debilitano le piante e le espongono all’azione di altri agenti e fungilli parassiti da debolezza… con un danno valutabile soltanto nell’anno seguente.
- Nel caso di una defogliazione completa [in maggio-giugno] si ha una perdita di incremento legnoso del 10-12% e, come conseguenza secondaria, la morte di parte dei rametti. Se la perdita totale delle foglie si verifica due volte in uno stesso anno, oltre a una diminuita produzione di legname che di solito supera il 30% si può avere la morte di tutti i rametti e anche di grosse ramificazioni.
Partendo da queste e da successive osservazioni condotte in campo venne in seguito pubblicato uno schema che illustra le perdite di incremento legnoso in funzione dell’intensità e dell’epoca di defogliazione ([21]).
Non erano tuttavia mai stati realizzati, almeno in ambiente padano, saggi scientifici ad hoc mirati a valutare quantitativamente gli effetti causati da intensità ed epoche diverse di defogliazione su pioppi in piantagione, ovvero in condizioni ordinarie di coltivazione del pioppeto, con metodologie statistiche appropriate, idonee a misurare la variabilità del fenomeno e a valutare correttamente le perdite rispetto a piante testimone non defogliate. Ciò è senza dubbio legato alla difficoltà di realizzare defogliazioni artificiali di entità nota, per un adeguato numero di replicazioni e in diversi momenti dell’anno, su piante che al secondo anno di coltivazione in pioppeto presentano un’altezza di circa 10 metri e una superficie fogliare già pari ad alcune decine di metri quadrati.
Anche la letteratura internazionale è molto scarsa riguardo allo studio degli effetti di defogliazioni artificiali indotte su pioppi in piantagione. I principali contributi provengono da autori nordamericani tra cui Reichenbacker et al. ([28]), che hanno defogliato artificialmente giovani pioppi in campo simulando il danno causato dal Coleottero Chrysomela scripta F. (asportando porzioni di foglia) e hanno valutato la crescita e la produzione di biomassa, oppure Tucker et al. ([30]), che hanno studiato gli effetti di intensità diverse di defogliazione sulla crescita di pioppi (P. deltoides) al secondo anno di coltivazione, sempre simulando un attacco di C. scripta, o ancora Moulinier et al. ([26]) che hanno defogliato per più anni consecutivi giovani piantagioni di Populus tremuloides Michx. simulando il danno da Malacosoma disstria Hübner. Molti studi sono stati invece condotti, per motivi di praticità, su giovani piante in ambiente controllato o in vivaio, tra i quali si cita a titolo di esempio quello di Bassman & Dickmann ([7]), che hanno studiato gli effetti delle defogliazioni sulla fotosintesi e la crescita di giovanissimi pioppi ibridi euramericani.
In questo articolo verranno presentati e discussi i risultati di tre saggi sperimentali condotti in pieno campo a Casale Monferrato (AL) e finalizzati a mettere in evidenza alcuni effetti di defogliazioni artificiali eseguite con intensità e in epoche dell’anno diverse, in alcuni casi anche per più anni consecutivi, su pioppi di Populus × canadensis clone I-214 al primo, secondo e terzo anno di coltivazione in pioppeto. In particolare sono stati studiati gli effetti sugli accrescimenti dei tronchi nello stesso anno delle defogliazioni e in quelli successivi, sullo sviluppo del germoglio apicale durante il primo anno di coltivazione e infine sulla fenologia della germogliazione delle piante negli anni successivi a quello della defogliazione. Ulteriori osservazioni sono state condotte sullo sviluppo di un fungillo considerato come un tipico parassita da debolezza dei pioppi, Discosporium populeum (Sac.) Sutton ([20]), che è stato allevato in vitro su estratti acquosi ottenuti dalla spremitura della corteccia di alberi sottoposti a livelli diversi di defogliazione, ma questi risultati costituiranno l’oggetto di un ulteriore specifico contributo.
Non è stato purtroppo possibile affrontare altri aspetti delle innumerevoli conseguenze che le defogliazioni possono provocare sulla fisiologia dei pioppi. Il problema è infatti molto complesso, con infinite interazioni tra le diverse componenti in gioco (pianta - insetto/malattia - ambiente). Un fattore che sicuramente incide in modo significativo sulle dinamiche di accrescimento dei pioppi è l’alterazione dei processi di accumulo delle sostanze di riserva e di trasferimento di queste ultime dalle foglie al fusto e alle radici prima del riposo vegetativo ([8], [14]); a questo proposito le defogliazioni tardive erano già state indicate quale causa di alterata fisiologia e di gravi deperimenti dei pioppi in Italia settentrionale ([4]). Non vanno inoltre trascurate le possibili interferenze negative delle defogliazioni sui processi, recentemente scoperti, di azotofissazione batterica sulla chioma di specie del genere Populus ([16]).
Tuttavia i risultati sperimentali dei saggi oggetto di questo articolo possono già fornire un contributo di chiarezza sull’entità dei danni conseguenti ad eventi di defogliazione e contribuire a indirizzare le future ricerche e le scelte colturali in pioppicoltura. Una migliore conoscenza della soglia di danno dei parassiti defogliatori permette infatti una razionalizzazione delle strategie di difesa dei pioppeti e un minor impatto ambientale della pioppicoltura che, come è noto, nell’ambiente padano viene sovente condotta in aree sensibili perifluviali dove è importante ridurre al minimo la distribuzione dei fitofarmaci ([15]).
Materiali e metodi
Sono stati realizzati tre saggi in due piantagioni sperimentali costituite con pioppi di Populus × canadensis clone I-214 all’interno dell’Azienda Sperimentale Mezzi, annessa al CREA-LF sede di Casale Monferrato (Alessandria). I terreni dell’Azienda, compresi nella golena del fiume Po, sono tipicamente alluvionali e presentano tessitura sabbio-limosa con limitata presenza di sostanza organica. Il clima della zona è di tipo temperato umido con estate molto calda (“Cfa” secondo la classificazione Köppen-Geiger) e presenta una temperatura media annua di 12 °C e precipitazioni pari a circa 700 mm/anno con due massimi, uno in primavera e uno in autunno ([29]).
La prima piantagione (A), nella quale sono stati realizzati i saggi n. 1 e n. 2, è stata messa a dimora nel febbraio 1997 su un appezzamento della superficie di 0.7 ettari caratterizzato da condizioni pedologiche omogenee; sono state utilizzate 271 pioppelle di due anni di vivaio collocate alla spaziatura di m 4.5 × 5.5. La seconda (B), nella quale è stato realizzato il saggio n. 3, è stata invece messa a dimora nel marzo 1998 utilizzando 211 pioppelle di un anno di vivaio collocate alla stessa spaziatura; l’appezzamento, della superficie di circa 0.6 ettari, era caratterizzato anche in questo caso da condizioni pedologiche omogenee.
Saggio n. 1
Nello stesso anno dell’impianto (1997), nella piantagione A le pioppelle sono state sottoposte a 4 livelli di defogliazione artificiale (25%, 50%, 75% e 100%) in tre diverse epoche dell’anno (G = inizio giugno, S = inizio settembre; GS = inizio giugno + inizio settembre), attribuendo 10 piante (replicazioni) a ciascun livello e a ciascuna epoca di defogliazione, secondo uno schema a parcelle monoalbero completamente randomizzato, per un totale di 13 tesi a confronto: T (Testimone non defogliato), G-025, G-050, G-075, G-100, S-025, S-050, S-075, S-100, GS-025, GS-050, GS-075, GS-100.
Le defogliazioni sono state realizzate asportando meccanicamente, tagliandone il picciolo, 1 foglia ogni 4 presenti sul germoglio per riprodurre la defogliazione del 25%, asportandone 2 su 4 per riprodurre quella del 50%, e così via (Fig. 2). Tale modalitàè stata preferita a quella di asportare corrispondenti porzioni percentuali di superficie da ogni singola foglia (più simile al danno effettivamente causato dall’attività trofica degli insetti defogliatori) non soltanto per motivi pratici e realizzativi, ma anche perché le foglie danneggiate in tal modo possono andare incontro a necrosi estese o a filloptosi anticipata, modificando pertanto l’estensione della superficie assimilante rimasta e alterando i risultati. Il personale che ha realizzato manualmente le defogliazioni si è avvalso di cestelli elevatori idraulici che, collegati alla presa di potenza della trattrice, sono normalmente impiegati per interventi di potatura e sono in grado di raggiungere altezze pari a 10-12 metri (Fig. 3).
Fig. 2 - Pioppelle al secondo anno di coltivazione artificialmente defogliate al 100%.

Fig. 3 - Operazioni di defogliazione artificiale con l’impiego di cestelli elevatori.

Il pioppeto è stato altresì protetto da eventuali defogliazioni di tipo parassitario (che avrebbero interferito con le valutazioni sperimentali) mediante trattamenti anticrittogamici e insetticidi al fogliame ripetuti con cadenza circa mensile; sono state inoltre eseguite periodiche irrigazioni e discature del terreno per mantenere il pioppeto in buone condizioni di vigoria ed evitare possibili filloptosi conseguenti a stress idrici o nutrizionali.
Con cadenza decadale a partire dall’epoca della prima defogliazione sono stati misurati il diametro dei tronchi a m 1.30 da terra e la lunghezza del germoglio apicale di ciascuna pianta (per ragioni di brevità e di comprensibilità sono tuttavia riportati nelle tabelle dei risultati unicamente gli accrescimenti registrati a fine anno). Le misurazioni del diametro delle piante sono proseguite anche per i tre anni successivi a quello della defogliazione (1998, 1999 e 2000); non è stato purtroppo possibile proseguire oltre per i gravi danni riportati dal pioppeto durante l’esondazione del fiume Po nel novembre 2000. Al momento della seconda defogliazione (inizio settembre) sono stati anche misurati il peso verde e secco nonché la superficie totale della chioma di 10 piante testimone (non defogliate) e di altrettante piante sottoposte a defogliazione del 100% durante il mese di giugno; questo dato ha permesso di valutare le eventuali differenze di massa fogliare a fine stagione tra piante completamente defogliate e piante non defogliate nonché le eventuali differenze di rapporto tra sostanza verde e secca nei due casi.
Saggio n. 2
Considerando che il saggio n. 1 ha avuto per oggetto defogliazioni realizzate nello stesso anno dell’impianto, quando le pioppelle avevano da poco superato lo stress del trapianto e il loro equilibrio fisiologico era probabilmente ancora fragile per la presenza di un limitato apparato radicale, interamente costituito da radici avventizie di nuova formazione, poteva permanere il dubbio che gli effetti delle defogliazioni si fossero assommati a quelli degli eventuali stress fisiologici del trapianto. Per questo motivo nel 1998 è stato realizzato, nella stessa piantagione sperimentale (A), un secondo saggio di defogliazione artificiale in cui sono state defogliate pioppelle al secondo anno del turno che non erano state incluse nel saggio n. 1 (mai precedentemente defogliate), già ben affrancate e fisiologicamente più equilibrate. Questo secondo saggio è stato caratterizzato da metodologia sperimentale identica a quella del primo salvo che, per motivi di limitata disponibilità di piante, le replicazioni attribuite a ogni tesi erano soltanto 5 anziché 10. Sono state eseguite misurazioni decadali del diametro dei tronchi a m 1.30 da terra nel triennio 1998-1999-2000 (la raccolta dei dati è stata interrotta dalla già citata alluvione del novembre 2000). Come nel saggio n. 1, al termine del 1998 sono stati misurati il peso verde e secco nonché la superficie totale della chioma di 5 piante mai defogliate e di altrettante sottoposte a defogliazione del 100% nel mese di giugno; la stessa misura è stata effettuata al momento della prima defogliazione (giugno) in modo da ottenere una valutazione della massa fogliare assimilante artificialmente asportata.
Saggio n. 3
Nella piantagione B, messa a dimora a inizio 1998 con materiale vivaistico di un anno, è stato realizzato un terzo saggio che aveva lo scopo di valutare gli effetti di una defogliazione totale (100%) della chioma (Fig. 2), realizzata nelle tre epoche già considerate nei precedenti saggi (giugno, settembre e giugno + settembre) e subita dalle pioppelle soltanto durante il secondo anno di coltivazione (1999) oppure, limitatamente a un sottocampione di piante, per due anni consecutivi (1999 e 2000). Le tesi a confronto in questo saggio erano pertanto costituite da T (Testimone non defogliato), G-100[1], S-100[1], GS-100[1], G-100[2], S-100[2], GS-100[2], dove il numero tra parentesi quadre indica il numero di anni in cui le piante sono state sottoposte a defogliazioni totali. È stato adottato uno schema sperimentale a blocchi randomizzati con 3 replicazioni e 10 piante per blocco. Sono state eseguite misurazioni decadali del diametro dei tronchi a m 1.30 da terra nel biennio 1999-2000; non è stato purtroppo possibile proseguire i controlli oltre tale data per i gravi danni riportati dalla piantagione in seguito al citato evento alluvionale.
Tutte le misure degli accrescimenti sono state sottoposte ad analisi della varianza (ANOVA) e la significatività delle differenze tra le medie è stata calcolata attraverso il test di Duncan o il test di Newman-Keuls utilizzando il software STATISTICA® 5.0 (StatSoft Inc., Tulsa, OK, USA).
Risultati
Saggio n. 1
Al termine del primo anno di coltivazione del pioppeto A (1997) sono apparse evidenti in particolare le riduzioni di accrescimento causate dalle defogliazioni realizzate nel mese di giugno dello stesso anno, dal momento che le defogliazioni di settembre hanno fatto sentire i loro effetti soprattutto sugli accrescimenti dell’anno successivo. I pioppi che in giugno avevano subito il 100% di defogliazione (G-100 e GS-100) hanno fatto registrare a fine anno una drastica riduzione di incremento diametrico (-40-50%), così come è stata sensibile la riduzione (circa -30%) subita dai pioppi defogliati al 75% (G-075 e GS-075). La riduzione di accrescimento accusata dai pioppi defogliati al 50% (G-050 e GS-050) è stata mediamente del 15%, statisticamente non significativa rispetto ai pioppi defogliati al 25% (G-025 e GS-025), che non hanno praticamente subito conseguenze negative, e al testimone (Tab. 1).
Tab. 1 - Saggio n. 1. Impianto 1997; defogliazione 1997. Incremento diametrico (mm) cumulativo delle pioppelle (differenza tra la misura finale e quella iniziale del diametro) e sua riduzione percentuale rispetto al testimone al termine di ciascun anno di saggio. Le medie contrassegnate da lettere diverse differiscono significativamente tra di loro (P<0.05) secondo il test di Duncan (Dt).
| Trattamento | 1997 | 1998 | 1999 | 2000 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mm | Dt | % | mm | Dt | % | mm | Dt | % | mm | Dt | % | |
| Testim. | 19.3 | e | - | 72.4 | de | - | 110.5 | cd | - | 129.2 | cde | - |
| G-025 | 19.3 | e | 0.1 | 74.0 | de | 2.8 | 113.2 | d | 2.4 | 135.2 | de | 4.6 |
| G-050 | 15.8 | bcde | -18.2 | 68.6 | bcde | -4.7 | 105.4 | bcd | -4.6 | 123.1 | cde | -4.7 |
| G-075 | 13.1 | abc | -32.0 | 61.1 | bcd | -15.1 | 99.8 | bc | -9.7 | 116.1 | bc | -10.1 |
| G-100 | 11.2 | ab | -41.9 | 60.3 | bc | -16.3 | 96.8 | bc | -12.4 | 119.5 | bcd | -7.5 |
| S-025 | 18.4 | de | -4.3 | 70.6 | cde | -1.9 | 108.8 | bcd | -1.5 | 125.7 | cde | -2.7 |
| S-050 | 17.1 | cde | -11.1 | 67.5 | bcde | -6.3 | 107.8 | bcd | -2.4 | 119.1 | bcd | -7.8 |
| S-075 | 17.7 | cde | -8.2 | 66.3 | bcde | -7.9 | 108.8 | bcd | -1.5 | 126.5 | cde | -2.1 |
| S-100 | 19.0 | e | -1.5 | 59.7 | bc | -17.1 | 96.8 | bc | -12.4 | 109.8 | b | -15.1 |
| GS-025 | 19.0 | e | -1.5 | 71.5 | cde | -0.7 | 108.6 | bcd | -1.7 | 128.1 | cde | -0.9 |
| GS-050 | 16.8 | cde | -12.9 | 66.1 | bcde | -8.2 | 106.7 | bcd | -3.4 | 124.7 | cde | -3.5 |
| GS-075 | 13.7 | abcd | -29.1 | 57.8 | b | -19.8 | 93.5 | b | -15.4 | 115.4 | bc | -10.7 |
| GS-100 | 9.8 | a | -49.3 | 44.1 | a | -38.8 | 81.2 | a | -26.5 | 99.0 | a | -23.4 |
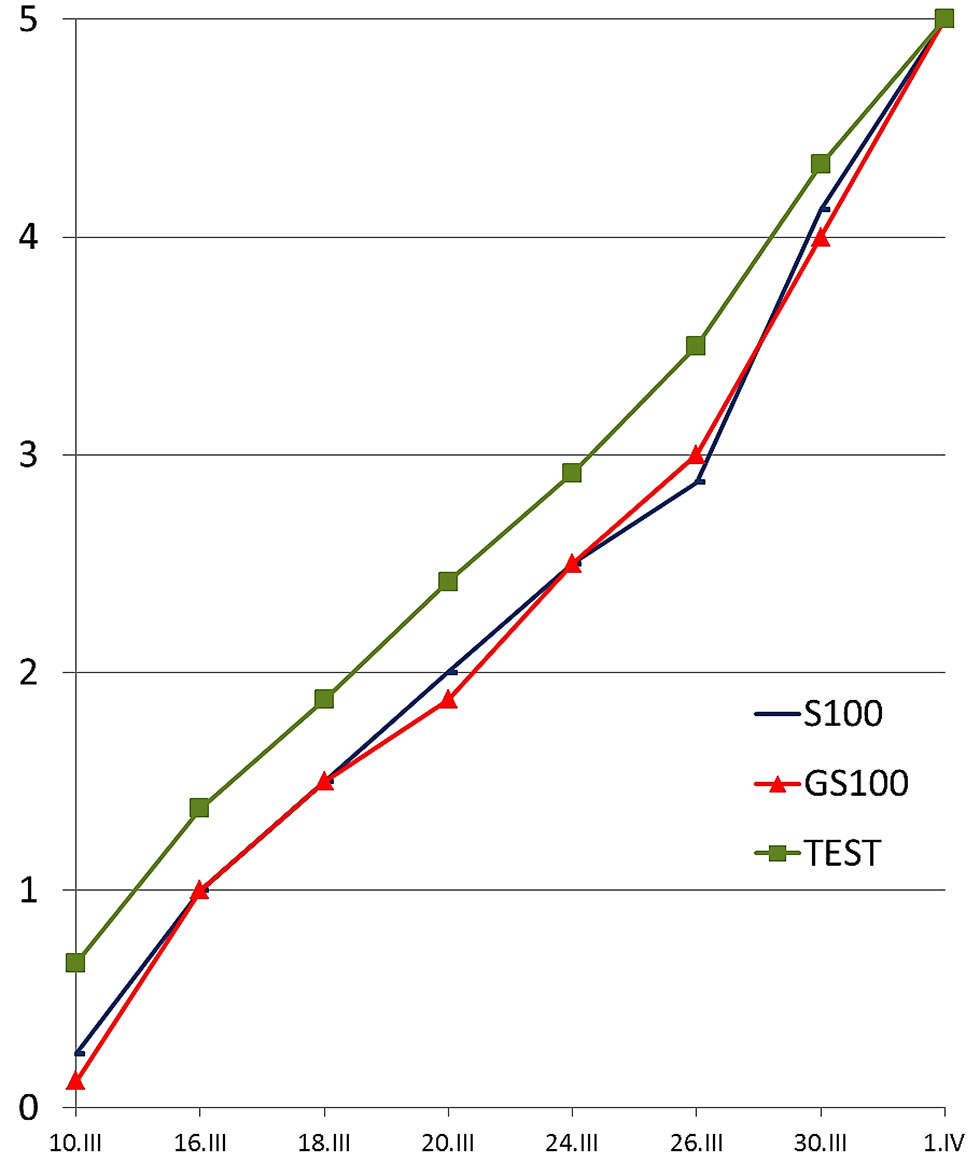
Al termine del primo anno di coltivazione (1997) la lunghezza del germoglio apicale delle pioppelle sottoposte a defogliazioni severe nel mese di giugno (G-075, G-100, GS-075, GS-100) è risultata pressoché dimezzata rispetto alle piante testimone (Fig. 4). Inoltre, le piante sottoposte a defogliazione totale nel mese di settembre (S-100) e quelle sottoposte a doppia defogliazione totale (GS-100) hanno accusato un ritardo di circa 3 giorni nelle prime fasi (1-4) della fenologia fogliare nella primavera successiva (1998), secondo la scala elaborata da Castellani et al. ([10]), arrivando tuttavia in contemporanea con le piante non defogliate alla fase 5 (Fig. 5). Le piante defogliate soltanto in giugno e quelle defogliate meno severamente in settembre non hanno invece accusato alcun ritardo nella germogliazione.
Fig. 4 - Test n. 1. Defogliazione 1997. Lunghezza del germoglio apicale a fine 1997 (termine della prima stagione vegetativa). Le medie contrassegnate da lettere diverse differiscono significativamente tra di loro (P<0.05) secondo il test di Newman-Keuls.
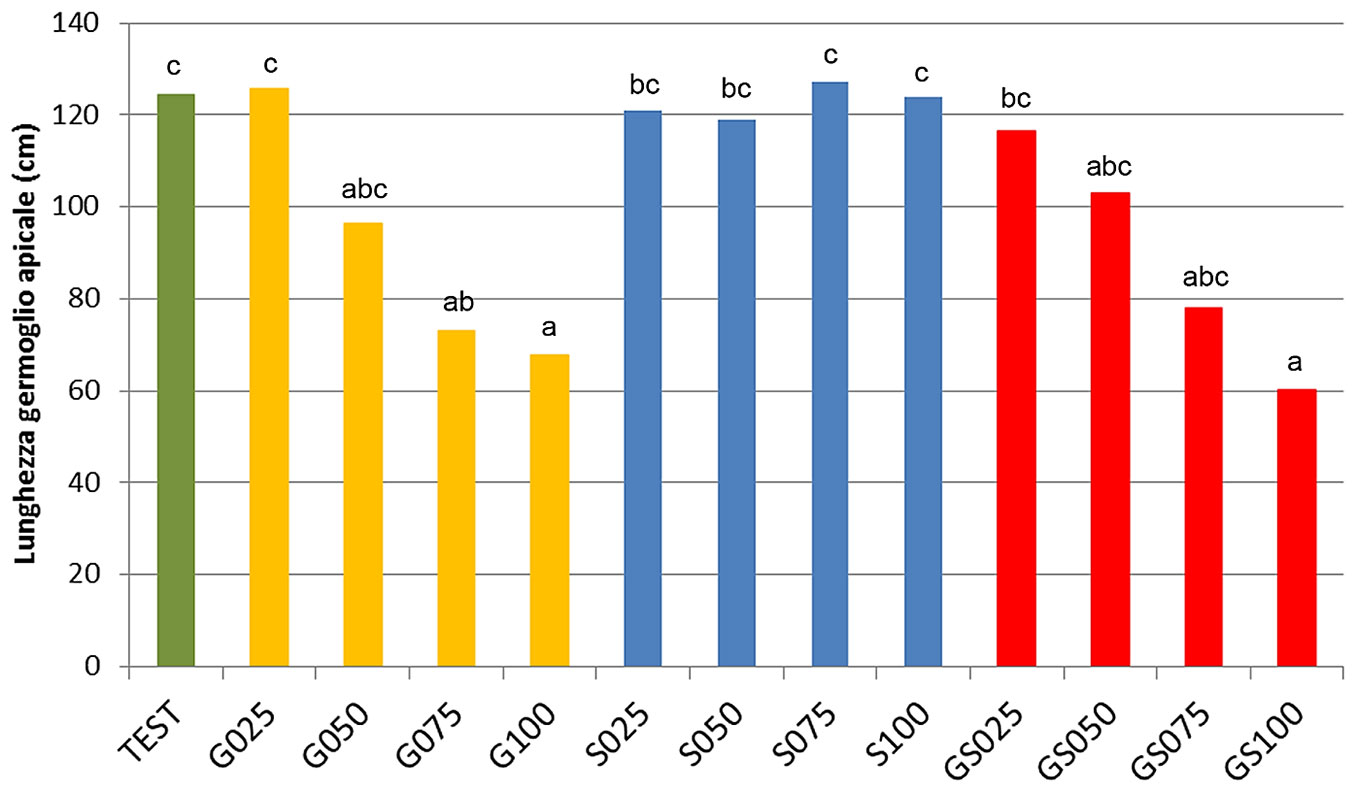
Fig. 5 - Test n. 1. Primavera 1998. Fasi fenologiche di pioppelle sottoposte a completa defogliazione nel settembre 1997 (S100) o a doppia completa defogliazione nel giugno e nel settembre 1997 (GS100) in confronto a pioppelle non defogliate (TEST).
La misurazione della superficie fogliare totale delle pioppelle a fine stagione 1997 ha messo in evidenza una riduzione della chioma del 64% nelle piante sottoposte a defogliazione totale nel mese di giugno rispetto a quelle non defogliate (Tab. 2). Non sono emerse differenze significative di rapporto tra i pesi di sostanza verde e secca nei due casi considerati (peso secco/peso verde = 33.4% e 34.5%, rispettivamente).
Tab. 2 - Saggi n. 1 e 2. Area fogliare totale (m2) di un pioppo al termine del primo anno (1997) e del secondo anno di coltivazione (1998) mai precedentemente defogliato (testimone) o completamente defogliato (G-100) nel giugno dello stesso anno, con le rispettive differenze percentuali. Le medie contrassegnate da lettere diverse differiscono significativamente tra di loro (P<0.05) secondo il test di Duncan (Dt).
| Trattamento | 1997 | 1998 | ||||
|---|---|---|---|---|---|---|
| m2 | Dt | % | m2 | Dt | % | |
| Testimone | 7.0 | b | - | 38.6 | b | - |
| G-100 | 2.5 | a | -63.7 | 23.9 | a | -38.1 |
Al termine del secondo anno di coltivazione (1998) le piante con maggiore riduzione di crescita (-39%) sono risultate quelle soggette alla doppia completa defogliazione dell’anno precedente (GS-100). Tra le altre tesi a confronto, le uniche a discostarsi significativamente dal testimone sono GS-075, S-100 e G-100, che presentano ancora riduzioni di accrescimento sensibili (-16-20% - Tab. 1).
Al termine del terzo anno di coltivazione (1999), pertanto a due anni di distanza dalle defogliazioni, le differenze di accrescimento tendono ad attenuarsi, ma è ancora sensibile la riduzione (-27%) accusata dalle piante sottoposte a doppia defogliazione completa (GS-100) e permangono le differenze significative rispetto al testimone nelle piante defogliate ai livelli più elevati (G-075, G-100, S-100 e GS-075), con riduzioni che vanno da -10% a -15% (Tab. 1). Per quanto riguarda i danni subiti dalla tesi GS-100, è importante considerare che, a fronte di un ridotto incremento diametrico del 27% rispetto al testimone alla fine del terzo anno, la differenza volumetrica del fusto, secondo la formula auxometrica definita da Frison ([17]), ammonta al 45%.
Alla fine del quarto anno di coltivazione (2000) le differenze rispetto al testimone sono ancora presenti, con le piante sottoposte a doppia defogliazione completa (GS-100) e quelle defogliate al 100% in settembre (S-100) che accusano ancora perdite significative di incremento diametrico pari al 23% e al 15% rispettivamente (Tab. 1). Rimangono significativamente diverse dal testimone anche le tesi G-075, G-100 e GS-075. La perdita in volume legnoso delle piante della tesi GS-100 rispetto al testimone ammonta al 41% secondo la già citata formula auxometrica.
I risultati di questo saggio evidenziano danni tendenzialmente superiori a quelli valutati da Arru ([5]) soprattutto al termine dell’anno stesso della defogliazione, anche se negli anni successivi del turno l’entità del danno tende a diminuire e ad avvicinarsi alle indicazioni di questo autore.
Saggio n. 2
In questo caso le defogliazioni artificiali sono state realizzate su piante al secondo anno di coltivazione (1998) mai precedentemente defogliate. Come nel saggio n. 1, al termine della stessa stagione di defogliazione (1998) sono state registrate perdite sensibili di incremento diametrico soltanto sulle piante più severamente defogliate a giugno, con riduzioni percentuali dal 20% circa (G-100, GS-075) fino al 33% (GS-100) rispetto al testimone (Tab. 3); tutte le altre tesi a confronto hanno subito riduzioni limitate con accrescimenti non significativamente diversi dal testimone. La superficie fogliare delle pioppelle al momento della defogliazione del mese di giugno era di circa 20 m2, mentre a fine stagione le piante che avevano subito una defogliazione totale in giugno presentavano una chioma con superficie ridotta del 39% rispetto alle piante non defogliate (Tab. 2). Come nel saggio precedente non sono emerse differenze di rapporto tra i pesi della sostanza verde e della sostanza secca nei due casi considerati.
Tab. 3 - Saggio n. 2. Impianto 1997; defogliazione 1998. Incremento diametrico (mm) cumulativo delle pioppelle (differenza tra la misura finale e quella iniziale del diametro) e sua riduzione percentuale rispetto al testimone al termine di ciascun anno di saggio. Le medie contrassegnate da lettere diverse differiscono significativamente tra di loro (P<0.05) secondo il test di Duncan (Dt).
| Trattamento | 1998 | 1999 | 2000 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| mm | Dt | % | mm | Dt | % | mm | Dt | % | |
| Testimone | 51.1 | c | - | 88.8 | cd | - | 125.5 | cd | - |
| G-025 | 52.0 | c | 1.8 | 92.5 | cd | 4.2 | 130.7 | cd | 4.1 |
| G-050 | 49.6 | c | -2.9 | 88.4 | cd | -0.5 | 126.3 | cd | 0.6 |
| G-075 | 48.8 | bc | -4.5 | 89.7 | cd | 1.0 | 122.2 | cd | -2.6 |
| G-100 | 41.0 | ab | -19.8 | 84.0 | bcd | -5.4 | 119.0 | bcd | -5.2 |
| S-025 | 52.2 | c | 2.2 | 94.8 | cd | 6.8 | 132.0 | cd | 5.2 |
| S-050 | 51.2 | c | 0.2 | 92.4 | cd | 4.1 | 129.3 | cd | 3.0 |
| S-075 | 50.4 | c | -1.4 | 92.7 | cd | 4.4 | 128.3 | cd | 2.2 |
| S-100 | 51.6 | c | 1.0 | 71.2 | b | -19.8 | 98.9 | b | -21.2 |
| GS-025 | 50.0 | c | -2.2 | 87.4 | cd | -1.6 | 126.9 | cd | 1.1 |
| GS-050 | 47.2 | bc | -7.6 | 87.9 | cd | -1.0 | 121.3 | bcd | -3.3 |
| GS-075 | 40.4 | ab | -20.9 | 76.6 | bc | -13.7 | 107.2 | bc | -14.6 |
| GS-100 | 34.4 | a | -32.7 | 47.9 | a | -46.1 | 62.9 | a | -49.9 |
Al termine del terzo anno di coltivazione (1999), secondo dalla defogliazione, si sono palesati anche gli effetti della defogliazione autunnale dell’anno precedente, con una forte riduzione di accrescimento (-46%) delle piante sottoposte a doppia defogliazione completa (GS-100) e con riduzioni sensibili anche a carico di S-100 (-20%) e di GS-075 (-14%). Tutte le rimanenti tesi non hanno invece presentato incrementi significativamente diversi dal testimone; si può notare inoltre che le tesi defogliate unicamente a giugno hanno recuperato buona parte delle perdite registrate nell’anno precedente (Tab. 3).
Alla fine del quarto anno di coltivazione (2000), la situazione è rimasta in larga parte identica a quella dell’anno precedente, con GS-100, GS-075 e S-100 che hanno evidenziato perdite significative di incremento rispettivamente del 50%, 15% e 21% rispetto al testimone (Tab. 3). Anche in questo caso è importante considerare che per quanto riguarda la tesi GS-100, a fronte di un suo ridotto incremento diametrico del 50% rispetto al testimone, la differenza volumetrica del fusto secondo la già citata formula auxometrica ammonta a circa il 60%.
I risultati di questo saggio sottolineano la elevata dannosità delle defogliazioni tardive più severe che, come aveva già osservato Arru ([5]), causano non soltanto ingenti perdite di produzione legnosa ma possono anche comportare danni qualitativi al legno per l’aumentata sensibilità delle piante verso parassiti da debolezza come Discosporium populeum o verso fisiopatie come l’alterazione delle “macchie brune”, capaci di provocare necrosi estese sul fusto e sui rami ([20]). La comparsa di queste avversitàè stata registrata nei due saggi descritti soprattutto su piante sottoposte a doppia defogliazione (GS) ai livelli di intensità più elevati (75% e 100%). Questi rilievi non hanno avuto carattere quantitativo, ma gli effetti delle defogliazioni artificiali sulla capacità germinativa e lo sviluppo dei conidi di D. populeum sono stati messi in evidenza attraverso saggi di laboratorio, i cui risultati verranno presentati in uno specifico articolo.
Saggio n. 3
In questo test sono stati studiati gli effetti di defogliazione totali (100%) della chioma realizzate nelle tre epoche (G, S e G+S) ma per un solo anno (1999) oppure per due anni consecutivi (1999 e 2000). Considerando le piante defogliate soltanto nel 1999, al termine di questo anno sono apparsi evidenti i danni causati dalla defogliazione totale di giugno (-46% e -39% di accrescimento diametrico per GS-100[1] e G-100[1] rispettivamente), mentre la defogliazione di settembre (S-100) ha fatto sentire i suoi effetti soprattutto nell’anno successivo (2000), al termine del quale tutte le tesi defogliate nel 1999 hanno accusato una riduzione di accrescimento diametrico significativamente diversa dalle piante testimone, con una perdita per la tesi GS-100[1] pari al 42%, mentre per S-100[1] e G-100[1] la perdita è stata del 20% circa. Nel corso del 2000 le piante della tesi G-100[1] hanno parzialmente recuperato la perdita di incremento subita nell’anno precedente.
Le piante sottoposte a defogliazioni complete in entrambi gli anni (1999 e 2000) hanno fatto registrare, a fine 2000, accrescimenti significativamente ridotti rispetto al testimone: le piante della tesi GS-100[2], che avevano subito 4 defogliazioni totali nel corso dei due anni, presentavano una riduzione del 46%, mentre le perdite ammontavano al 30% e al 25% per le piante delle tesi G-100[2] e S-100[2], che avevano subito due defogliazioni nei due anni (Tab. 4). Purtroppo l’interruzione del saggio a causa dell’alluvione del novembre 2000 ha impedito che potessero essere compiutamente valutati i danni causati dall’ultima defogliazione di settembre, che si sarebbero resi manifesti nell’anno successivo, ovvero nel 2001.
Tab. 4 - Saggio n. 3. Impianto 1998; defogliazione solo nel 1999 [1] o in entrambi gli anni, 1999 e 2000 [2]. Incremento diametrico (mm) cumulativo delle pioppelle (differenza tra la misura finale e quella iniziale del diametro) e sua riduzione percentuale rispetto al testimone al termine dei due anni di saggio. Le medie contrassegnate da lettere diverse differiscono significativamente tra di loro (P<0.05) secondo il test di Duncan (Dt).
| Trattamento | 1999 | 2000 | ||||
|---|---|---|---|---|---|---|
| mm | Dt | % | mm | Dt | % | |
| Testimone | 31.5 | b | - | 65.6 | d | - |
| G-100 [1] | 19.1 | a | -39.4 | 53.2 | c | -18.9 |
| S-100 [1] | 30.6 | b | -3.0 | 52.2 | c | -20.4 |
| GS-100 [1] | 17.2 | a | -45.5 | 37.9 | ab | -42.2 |
| G-100 [2] | - | - | - | 45.9 | bc | -30.1 |
| S-100 [2] | - | - | - | 49.0 | c | -25.2 |
| GS-100 [2] | - | - | - | 35.4 | a | -46.1 |
Pur considerando i limiti legati alla chiusura anticipata del test e alla mancata acquisizione dei dati relativi agli effetti cumulati di defogliazioni ripetute negli anni, anche da questo saggio emergono evidenze che confermano le importanti riduzioni di accrescimento delle pioppelle in seguito alla totale perdita delle foglie, soprattutto per eventi ripetuti più volte nel corso di uno stesso anno e, con i limiti imposti dalla breve durata del saggio, per eventi ripetuti per più anni consecutivi.
Discussione
I risultati dei tre saggi sperimentali hanno fornito le seguenti indicazioni:
- Un singolo evento di defogliazione totale determina, nello stesso anno o in quello successivo a seconda che si verifichi precocemente o tardivamente nella stagione vegetativa, una riduzione di incremento diametrico del 20% (in piante ben affrancate al secondo anno di coltivazione) fino al 40% (in piante da poco messe a dimora). Defogliazioni di entità minore hanno conseguenze limitate sulle piante ben affrancate.
- Due eventi di defogliazione al 75% o al 100% nel corso di una singola annata determinano gravi problemi fisiologici e una riduzione di accrescimento diametrico pari al 40-50%, valutabile appieno soltanto nell’anno successivo a quello delle defogliazioni, quando appaiono evidenti altre conseguenze come un ritardo della germogliazione, la comparsa di attacchi di parassiti da debolezza e la morte di parte dei rami. La perdita del 50% o meno delle foglie provoca invece danni limitati.
- I danni provocati da defogliazioni intense che si ripetono per più anni consecutivi tendono ad aggravarsi progressivamente nel tempo soprattutto se queste si verificano in epoca tardiva, probabilmente per un sempre maggior scadimento dell’equilibrio fisiologico delle piante (la durata forzatamente limitata del saggio n. 3 non ha tuttavia consentito un’approfondita valutazione di questo aspetto).
- Le defogliazioni precoci di elevata intensità provocano riduzioni di accrescimento (sia in altezza che diametriche) sensibili ma tendenti a essere compensate e ridimensionate con l’avanzare dell’età della piantagioni, mentre forti defogliazioni tardive (a fine stagione vegetativa) provocano perdite che perdurano negli anni e che vengono difficilmente riassorbite.
- Un pioppo al secondo anno di coltivazione che venga defogliato completamente in epoca precoce è in grado di formare una nuova chioma che, a fine stagione, presenta una superficie ridotta del 40% circa rispetto alla chioma di una pianta non defogliata. Una pioppella nell’anno del trapianto è invece in grado di riformare una minor superficie di chioma (-64% rispetto a una chioma normale), probabilmente in relazione allo stress del trapianto e a un equilibrio fisiologico più fragile. Anche questo dato concorre a spiegare perché i danni provocati dalle defogliazioni sulle pioppelle al primo anno di coltivazione siano più elevati.
Il quadro che viene così delineato conferma sostanzialmente le valutazioni di Arru ([5]), salvo che l’entità del danno causato dalle defogliazioni totali precoci, sulla base dei risultati dei tre saggi, al termine dello stesso anno della defogliazione appare significativamente più elevata rispetto alle stime di questo Autore. I saggi hanno tuttavia evidenziato il fatto che queste perdite vengono progressivamente riassorbite durante il turno di coltivazione. Un esempio di elevata capacità di crescita compensatoria è notoriamente fornito dalla betulla, che è in grado di recuperare le perdite di accrescimento subite a causa di defogliazioni severe ([31]).
In ogni caso, se consideriamo il costo di un singolo intervento antiparassitario alla chioma come pari mediamente a 80 €/ettaro (il costo può variare da 60 a 100 € circa a seconda della dimensione delle piante e del prodotto utilizzato), una produzione media legnosa del pioppeto pari a 20 m3/ettaro/anno (stima ampiamente superata dai pioppeti coltivati nelle aree padane maggiormente vocate) e un valore attuale di mercato del legno di pioppo pari a 50 €/m3 (D. Coaloa, comunicazione personale), ne consegue che il costo del trattamento incide sul valore della produzione dell’anno per l’8% (per lo 0.8% sul valore totale della produzione a fine ciclo considerando un turno di 10 anni). Ciò significa che, laddove sia da temere una defogliazione parassitaria di elevata intensità nel pioppeto, dal punto di vista strettamente economico (ovvero quando non prevalgano altri aspetti, per esempio di carattere ambientale) l’intervento antiparassitario a protezione delle chiome è ampiamente giustificato in considerazione del danno economico potenziale. Si potrebbe affermare inoltre che l’intervento è caldamente raccomandabile per impedire defogliazioni intense tardive, come potrebbero essere quelle causate dal Lepidottero Hyphantria cunea o dai funghi agenti di ruggini (Melampsora spp.), che tipicamente compaiono nell’estate e provocano defogliazioni intense sui cloni sensibili ([25], [1], [19]). È stato infatti dimostrato attraverso questi saggi che i danni causati dalle defogliazioni tardive incidono molto pesantemente sull’equilibrio fisiologico delle piante e causano perdite di accrescimento importanti che difficilmente vengono compensate nell’arco del turno, aprendo inoltre la strada a pericolosi parassiti da debolezza ([20]), causa di ulteriori danni di tipo qualitativo.
I pesanti danni economici che la pioppicoltura potrebbe soffrire a causa di attacchi da parte di parassiti defogliatori, ben evidenziati dai risultati precedentemente discussi, nonché le note controindicazioni di carattere ambientale (oltre che economico) legate ai relativi interventi di difesa suggeriscono l’importanza di attuare strategie di ricerca per un miglioramento genetico del pioppo mirato all’individuazione di genotipi dotati non soltanto di buone caratteristiche di accrescimento, di adattabilità e di qualità del legno, ma anche di elevata resistenza verso queste avversità. Già oggi molti cloni di pioppo tra quelli iscritti ai Registri nazionali dei Materiali di Base di vari Paesi dell’Unione Europea sono resistenti alle malattie fogliari ([25], [18]) e possono pertanto essere coltivati nelle aree maggiormente a rischio.
Ringraziamenti
Numerosi colleghi ricercatori, tecnici e operai hanno collaborato alla realizzazione dei saggi in campo: nell’impossibilità di citarli tutti, li ringrazio collettivamente, ma sento il dovere di ringraziare in modo particolare Bernardino Bianco, Giuseppe Deandrea, Gianni Facciotto, Achille Giorcelli, Giuseppe Negro, Franco Picco e Renzo Rossino, il cui impegno è stato determinante per il buon esito del progetto. Ringrazio ancora Pier Mario Chiarabaglio, Domenico Coaloa, Gianni Facciotto e Achille Giorcelli per la lettura critica del testo e i preziosi suggerimenti, Lucia Sebastiani per la revisione linguistica dell’abstract in inglese. Desidero infine ricordare e ringraziare i ricercatori entomologi che mi sono stati maestri e che per primi hanno affrontato lo studio dei danni causati dagli insetti defogliatori in pioppicoltura, Giovanni Arru, recentemente scomparso, e Gianfranco Lapietra.
References
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar