The vulnerability of Mediterranean forests to extreme heat and drought events: the case of Holm oak (Quercus ilex L.) dieback in Italy
Forest@ - Journal of Silviculture and Forest Ecology, Volume 22, Pages 56-65 (2025)
doi: https://doi.org/10.3832/efor4934-022
Published: Oct 14, 2025 - Copyright © 2025 SISEF
Research Articles
Abstract
Extreme heat waves coupled with severe drought episodes are increasing in frequency in the Mediterranean region. Such climatic events pose significant threats to evergreen sclerophyllous vegetation, despite their inherent resistance and adaptation to drought conditions. Over recent decades, several cases of dieback affecting Quercus ilex, Quercus suber, and other evergreen shrubs have been reported across the Mediterranean basin. In Italy, the first reports of Q. ilex dieback due to climatic events date back to 2012 in Tuscany, and with more extensive impacts observed following the extreme drought of 2017. During the summer of 2024, widespread dieback phenomena were reported across Southern Italian regions, including Apulia, Basilicata, and the islands of Sardinia and Sicily. The symptoms were consistent and recurrent in all the affected areas, characterized by partial or total foliar browning, followed by wilting and branch dieback. Notably, resprouting from dormant buds was observed in affected branches and leaves in the following rainy season. Affected areas showed ongoing symptoms of oak decline, often associated with the presence of Phytophthora spp. and other opportunistic pathogens, suggesting a complex interaction between biotic and abiotic stressors. This study aims to document and analyze the most relevant cases of Q. ilex dieback occurring in central and southern Italy, using also data collected through the SilvaCuore web application. The phenomenon of dieback and mortality of Q. ilex is still poorly understood in terms of the cause-and-effect relationships, the relative contribution of biotic and abiotic factors, the extension and severity of damage, and the ecological consequences. Although Mediterranean woodlands (forests and shrublands) are scarcely represented in the current scientific and monitoring programs (ICP Forests Level I network), the available results show an increase in dieback and mortality in Italy since 2012. We emphasize the urgent need for coordinated research efforts to study and map this phenomenon. Proposed actions include establishing a network of permanent monitoring plots that combine phytopathological and ecophysiological assessments over time, deploying comprehensive terrestrial and remote sensing monitoring techniques, and involving citizens in early detection efforts. Finally, the research should aim to identify the site-specific and management-related factors that have contributed to the extent and intensity of the dieback phenomenon. Understanding these drivers is critical for developing effective management strategies aimed at mitigating the impacts of these dieback events and enhancing the resilience of the most vulnerable forest stands.
Keywords
Climate Change, Crown Dieback, Forest Health Monitoring, Mediterranean Forest Decline, Tree Mortality
Introduzione
Negli ultimi decenni la vulnerabilità delle foreste ai cambiamenti climatici è aumentata notevolmente ([5], [6], [23], [45]). L’aumento delle temperature associato al cambiamento climatico ha incrementato il deficit di pressione del vapore e la domanda evaporativa, causando siccità più lunghe e più intense ([74]), con conseguente riduzione della produttività forestale e della crescita degli alberi ([38], [60]). Queste “siccità calde” rappresentano oggi una minaccia significativa per la conservazione delle foreste in varie regioni ([39], [40]). Le ondate di calore e i periodi di siccità stanno causando la morte di alberi nella maggior parte dei biomi dei diversi continenti (Nord America, Europa, Australia, Asia continentale e Russia), con un forte aumento degli eventi di mortalità a partire dalla fine del secolo scorso ([3], [4]).
In Italia il 37% del territorio è coperto da boschi che da diversi anni presentano fenomeni di mortalità di individui appartenenti soprattutto al genere Quercus. La sintomatologia che accompagna la morìa delle querce è rappresentata da: avvizzimento delle chiome e defogliazione, disseccamento dei rami, formazione di germogli epicormici, necrosi delle radici, riduzione della biomassa, ecc., fino ad arrivare al caso più estremo con la morte della pianta ([42]). Tali sintomi sono riconducibili al fenomeno noto come deperimento delle querce (oak decline), una sindrome complessa causata da molteplici fattori biotici e abiotici, che comporta una progressiva perdita di vitalità fino a provocare la morte delle piante ([59], [71], [29]). Fenomeni di deperimento sono stati riportati negli ultimi decenni anche su querce mediterranee, in particolare su quercia da sughero (Quercus suber L.) e leccio (Quercus ilex L.) ([32], [64], [47], [44], [31], [65], [19], [13]).
Nella nostra penisola, il disseccamento delle querce, imputabile principalmente a periodi di siccità, fu inizialmente rilevato nei boschi dell’Italia nord-orientale (Quercus petraea Matt. Liebl. e Quercus robur L.), in cui la farnia risultava essere la specie quercina maggiormente colpita ([27], [55]). Il fenomeno si è poi verificato nelle zone centrali e meridionali colpendo principalmente il cerro (Quercus cerris L.), la roverella (Quercus pubescens Willd.) e il farnetto (Quercus frainetto Ten.) nell’areale del Parco Nazionale del Pollino ([35], [24], [25], [26]), che mostrano evidenti sintomi di disseccamento causati dalla siccità, identificata come il principale fattore di disturbo.
Negli ultimi anni il fenomeno si è intensificato ulteriormente, e i casi segnalati sono più che triplicati. Guardando al primo ventennio del secolo appena iniziato, si potrà notare che nei paesi mediterranei si sono verificate ripetute annate di forte stress da siccità e calore. Nel 2017 in molte aree del Centro-Sud non si sono verificate precipitazioni per oltre 3 mesi, con temperature di ben 8 gradi sopra la media del periodo. Questi fenomeni portarono ad un indebolimento dei soprassuoli forestali coinvolti, innescando processi che hanno portato a estesi casi di mortalità degli alberi nell’arco di pochi anni ([62]). Le “siccità calde”, caratterizzate dalla concomitanza di stress termico e idrico, sono considerate tra le principali cause della mortalità attuale degli alberi ([17], [8], [34]). Questi eventi climatici inducono la cosiddetta “disfunzione idraulica” (hydraulic failure), che si manifesta attraverso embolie xilematiche, e la “fame di carbonio” (carbon starvation), determinata dalla riduzione della disponibilità di carbonio. Tale riduzione si verifica a seguito di prolungati periodi di arresto dell’attività fotosintetica quale strategia conservativa al fine di chiudere gli stomi per ridurre eccessive perdite d’acqua ([50], [51], [10], [9]).
I fenomeni di disseccamento e mortalità si presentano spesso in maniera puntiforme, essendo influenzati da condizioni locali (profondità e caratteristiche del suolo, esposizione, pendenza), e la loro conoscenza rimane frammentata nonostante i recenti progressi nel monitoraggio. Una più precisa conoscenza della loro intensità e distribuzione è di primaria importanza per comprendere le cause e le conseguenze e poter così indirizzare la gestione. Il problema degli impatti prodotti dagli estremi climatici sul leccio e sulla vegetazione sempreverde mediterranea è arrivato alla pubblica attenzione nel corso del 2024 attraverso numerosi interventi informativi, divulgati con diversi mezzi di comunicazione. In molte regioni dell’Italia centro-meridionale, l’estate 2024 è stata, infatti, caratterizzata da temperature estremamente alte, con una grande incidenza di notti tropicali, che hanno determinato disseccamenti estesi nelle leccete. Il fenomeno ha quindi assunto rilevanza nazionale, anche se l’estensione e l’intensità del problema rimangono poco conosciute, così come le implicazioni gestionali per la conservazione delle foreste mediterranee.
Gli impatti sui boschi mediterranei
L’ambiente mediterraneo, uno dei più preziosi sistemi ecologici della Terra, è caratterizzato da un’elevata biodiversità, ma è anche contraddistinto da una notevole fragilità e vulnerabilità agli impatti del cambiamento climatico ([53]). Numerosi studi scientifici e rapporti internazionali ([41]) identificano la regione mediterranea come uno dei principali hotspot del cambiamento climatico globale, con previsioni di condizioni sempre più calde e aride nel prossimo futuro ([36]).
La vegetazione sclerofilla sempreverde mediterranea, da sempre soggetta agli stress e alle pressioni ambientali tipiche delle regioni in cui si è evoluta, negli ultimi anni è chiamata ad affrontare nuove sfide legate all’intensificarsi del cambiamento climatico, in particolare a causa delle sempre più frequenti ondate estreme di calore e siccità. Casi più o meno estesi di disseccamento e moria nelle foreste mediterranee sono ormai ben documentati nelle diverse regioni del mondo a clima mediterraneo ([52]), come il Cile (Venegas-González et al. 2023), la California (Venturas et al. 2016) e l’Australia meridionale ([49]). Nel bacino del Mediterraneo i disseccamenti hanno riguardato formazioni sempreverdi (latifoglie e conifere) dell’Europa meridionale ([16]) e del Nord Africa ([54]), interessando sia la vegetazione di latifoglie sclerofille sempreverdi delle aree costiere che le conifere oro-mediterranee.
La sclerofillia non rappresenta un adattamento specifico alla siccità ([63]). Le sclerofille sempreverdi mediterranee possono soffrire della combinazione di alte temperature e bassa umidità atmosferica, che determinano alti valori di VPD (Vapour Pressure Deficit). L’alto VPD induce fenomeni di cavitazione xilematica e rende difficile il recupero dalla cavitazione stessa. Le conseguenze ecologiche, per quanto riguarda la persistenza della lecceta (e più in generale della copertura vegetale), dipendono dalla durata, dall’intensità e dal ripetersi degli eventi estremi di calore e siccità. Questi eventi determinano la gravità dell’impatto in termini di perdita della chioma e mortalità del fusto. Le specie di latifoglie sempreverdi mediterranee possono rigenerare vigorosamente da gemme avventizie alla base del fusto dopo tali episodi, assicurando la persistenza del bosco. Tuttavia, la resilienza della vegetazione mediterranea potrebbe essere ridotta dal ripetersi di siccità estreme a causa della progressiva deplezione delle riserve di amido e zuccheri solubili, compromettendo la capacità delle piante di fronteggiare ulteriori condizioni di stress ([48]).
Il monitoraggio delle condizioni delle foreste mediterranee
In Italia, varie ricerche e attività di monitoraggio hanno evidenziato un aumento dei fenomeni di deperimento e mortalità di diverse specie sempreverdi tipiche della foresta mediterranea, in particolare il leccio e la quercia da sughero. L’alterazione del regime climatico, con ondate di calore e siccità prolungata ormai sempre più frequenti, è ritenuta uno dei principali fattori predisponenti al già ben noto deperimento ([35], [1]). A ciò si aggiunge la diffusione di agenti patogeni, come Phytophthora spp., considerati tra i principali fattori biotici coinvolti nel deperimento delle querce ([12], [64], [26], [66]) e agenti di cancro come Diplodia corticola e Biscogniauxia mediterranea ([47]). Al contrario, sono ancora limitate le conoscenze sui più recenti fenomeni di disseccamento che hanno interessato prevalentemente il leccio e altre specie della macchia mediterranea, con manifestazioni più acute e repentine in corrispondenza di periodi estremamente caldi e siccitosi. Le prime osservazioni sporadiche relative a questi comportamenti risalgono al 2012 in Toscana, ma evidenze più ampie sono state ottenute a seguito dell’ondata estrema di calore e siccità del 2017 nell’Italia centrale ([56], [58]), con un quadro sintomatologico coerente e costante (Fig. 1). I sintomi sono costituiti da: ingiallimento e imbrunimento totale o parziale delle foglie (Fig. 1E-F), avvizzimento e disseccamento delle stesse e dei germogli apicali e, nei casi più gravi, delle ramificazioni principali e del fusto fino alla morte dell’individuo (Fig. 1A-B). Quando il disseccamento ha interessato la parte superiore del fusto, si è osservata la comparsa di getti epicormici nella chioma e nella parte inferiore del fusto (Fig. 1C-D). Spesso gli esemplari di leccio disseccati presentavano anche la necrosi dell’apparato radicale riconducibile ad infezioni da Phytophthora spp. In qualche caso è stata osservata la presenza di cancri carboniosi sul fusto dovuti a infezioni di Biscogniauxia mediterranea e la mortalità di individui giovani e adulti. Nel periodo autunnale, sono state rilevate la schiusura di gemme dormienti e la comparsa di nuovi getti sulle piante meno compromesse.
Fig. 1 - Quadri sintomatologici osservati su leccio: disseccamento dell’intera chioma - Sardegna (A); disseccamento dei getti apicali - Sardegna (B); disseccamento dei rami e presenza di getti epicormici - Toscana (C); germogli epicormici lungo il ramo - Sardegna (D); imbrunimento disseccamento delle foglie - Sicilia (E); clorosi fogliare - Toscana (F).

Le condizioni delle foreste in Italia, in termini di defogliazione e mortalità degli alberi, sono oggetto di monitoraggio nell’ambito del programma CON.ECO.FOR. (ICP Forests), a cura dei Carabinieri Forestali ([15]). I risultati generali indicano un pronunciato e progressivo peggioramento delle condizioni dei boschi a partire dal 2010, in linea con quanto osservato anche a livello europeo ([57]). In Italia le formazioni a leccio, sughera e macchia mediterranea ammontano a circa il 10% della superficie forestale; tuttavia, questo tipo fisionomico risulta sottorappresentato nel programma CON. ECO.FOR. (11 aree permanenti di monitoraggio delle 261 a livello nazionale). I risultati, pur suggerendo un incremento del fenomeno di deperimento (espresso come defogliazione e mortalità) anche nella vegetazione mediterranea ([15]), non sembrano sufficientemente rappresentativi della reale entità e tendenza degli impatti sulla vegetazione mediterranea.
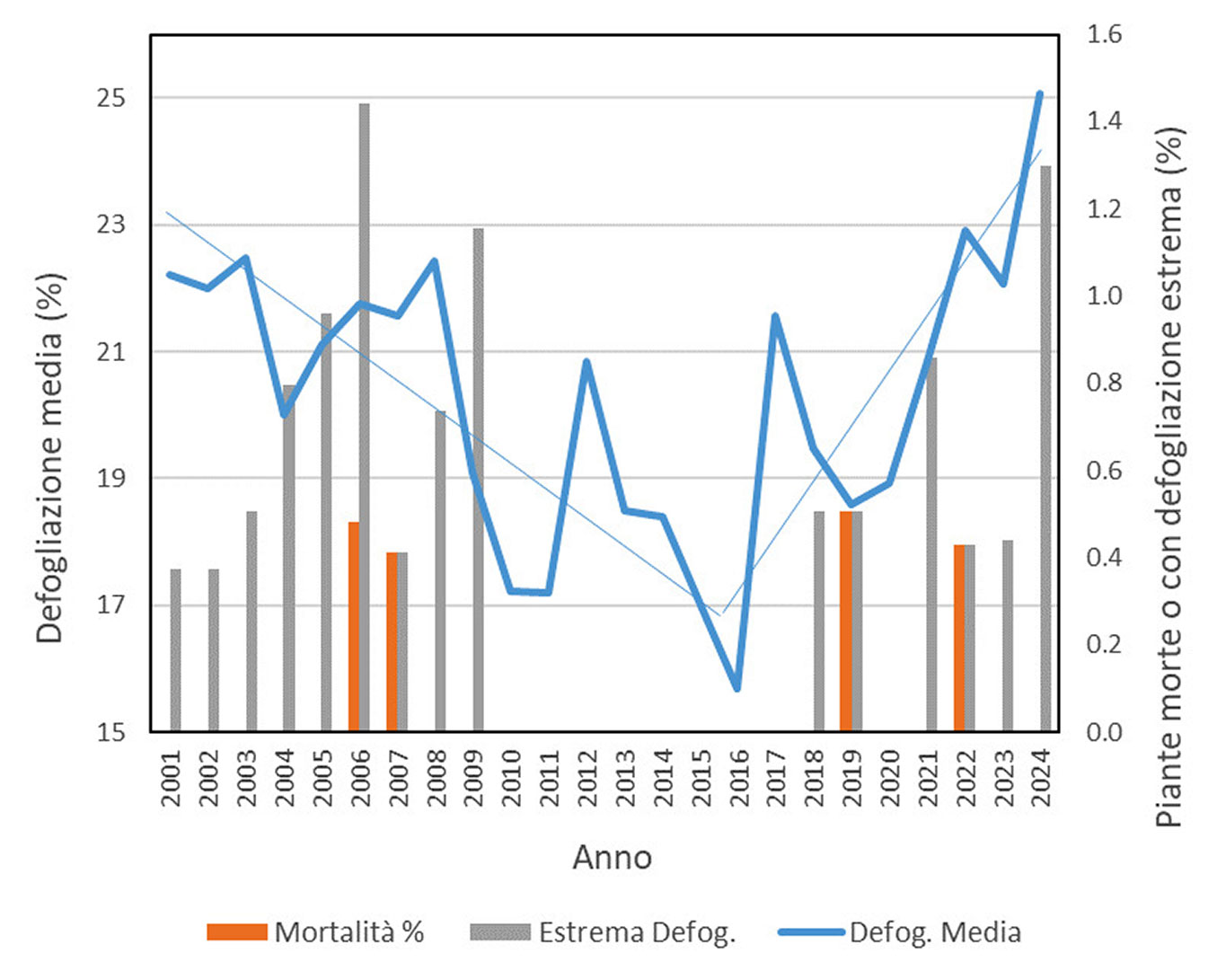
La Fig. 2 mostra i risultati dell’indagine CON.ECO.FOR. (ICP Forests, Livello I) sulle condizioni di defogliazione e mortalità in Quercus ilex a livello nazionale (Italia) dal 2001 al 2024. L’indagine ha preso in considerazione circa 250 piante distribuite in 11 aree di monitoraggio permanenti (per dettagli, vedi [15]). Per quanto riguarda la defogliazione media, si possono distinguere due andamenti distinti: dal 2001 al 2016 si osserva una riduzione della defogliazione, mentre dopo quell’anno si osserva un deciso aumento, con valori massimi nel 2025 (andamenti significativi, p<0.05, test Mann-Kendall). Dal 2016 il trend di defogliazione mostra dei picchi nei valori medi in corrispondenza degli anni caldi e siccitosi, ovvero il 2017, il 2022 e il 2025. La mortalità è sporadica e di entità molto limitata. Tuttavia, se insieme alle piante morte si considerano anche quelle con defogliazione estrema (>85%), si osserva un aumento nell’ultimo anno. Piante con defogliazione maggiore all’85% sono diffuse anche dopo il 2003, anno in cui durante l’estate sono state registrate temperature estreme per intensità e durata. Questo comportamento è attribuibile alla grande capacità di rigenerazione di questa specie attraverso gemme soppresse o avventizie, che rende l’effettiva morte della pianta difficile da diagnosticare.
Fig. 2 - Defogliazione e mortalità su Quercus ilex in Italia, secondo l’indagine CON.ECO.FOR. La linea blu mostra il livello di defogliazione media percentuale. Gli istogrammi indicano la percentuale di piante morte e con defogliazione estrema (>85%). La defogliazione è espressa secondo una scala percentuale che va da 0 (pianta con chioma intatta) a 100 (pianta morta) (Fonte: Arma dei Carabinieri - Comando Unità Forestali, Ambientali e Agroalimentari).
La situazione nelle regioni
Di seguito vengono riportate le osservazioni effettuate dagli autori in alcune regioni dell’Italia centro-meridionale e insulare. Il quadro che ne risulta non ha la pretesa di essere esaustivo, ma costituisce una prima rappresentazione di un fenomeno in evoluzione, che verrà arricchita da ulteriori contributi che ne consentano l’interpretazione degli agenti causali e della diffusione. Nella Fig. 3 sono illustrate alcune situazioni panoramiche nelle singole regioni.
Fig. 3 - Panoramica sui fenomeni di disseccamento riscontrati in boschi di quercia nelle diverse regioni italiane: Sicilia (A, B) lecceta di Monte Tre Frati (Etna) prima dei fenomeni di disseccamento (2022) (A) e nel mese di luglio 2024 (B); Sardegna (C) versante con disseccamenti diffusi in Ogliastra, 2024; Toscana (D) versante con disseccamenti nella Riserva di Monterufoli-Caselli (Pisa), 2017; Puglia (E) versante con disseccamenti nel Gargano, 2024; Basilicata (F) versante con disseccamenti nelle Piccole Dolomiti Lucane, 2017.

Basilicata
In Basilicata, fenomeni di deperimento delle foreste causati dalla siccità vengono riportati già da alcuni decenni, in particolare quelli innescati tra la fine degli anni ’90 e l’inizio del 2000, in diverse aree interne del territorio regionale, in particolare nel Parco Nazionale del Pollino. In queste aree, caratterizzate dalla quasi totale assenza di interventi di gestione da quasi mezzo secolo, sono state condotte analisi multi-proxy sugli anelli degli alberi (ad esempio Quercus frainetto Ten.), considerando aspetti relativi alla struttura dell’albero (crescita radiale e anatomia del legno) e a diversi indicatori fisiologici e funzionali, quali l’efficienza d’uso dell’acqua valutata tramite analisi isotopiche di δ13C e δ18O, nonché l’analisi dei carboidrati non strutturali, prima e dopo il disseccamento indotto dalla siccità ([24], [25], [61]). Inoltre, sono stati condotti studi per approfondire il possibile coinvolgimento di agenti patogeni, come Phytophthora quercina, nel complesso fenomeno di deperimento delle foreste miste di cerro e roverella ([27]). Da tali studi è emerso che la mortalità degli alberi legata alla siccità sta determinando una sostituzione parziale o totale delle tipologie di vegetazione in diversi biomi. Le foreste dominate da specie tipiche di ambienti più umidi stanno evolvendo verso popolamenti caratterizzati da specie più adattate a condizioni aride. In particolare, nei querceti della Basilicata si è osservata una tendenza all’autosostituzione, ovvero la sostituzione della specie dominante con la stessa specie, fenomeno che, sebbene non desti preoccupazioni immediate, assume un significato preoccupante alla luce di recenti studi sulla rinnovazione. Questi indicano una significativa riduzione della presenza di giovani alberi e di rinnovazione in alcune aree dei querceti lucani, suggerendo una perdita di vigore di queste foreste e sollevando preoccupazioni sul loro futuro. A seguire, un fenomeno di particolare rilievo che di recente ha attirato l’attenzione è quello relativo all’andata di calore e siccità avvenuti nel 2017 con sintomi di disseccamenti che hanno colpito i boschi lungo il versante del Parco Regionale di Gallipoli Cognato e Piccole Dolomiti Lucane (Fig. 3F). In quello stesso anno, analisi condotte tramite immagini satellitari hanno evidenziato che circa 50.000 ettari su un totale di 350.000 della superficie boscata in Basilicata presentavano un elevato stato di stress, caratterizzato da disseccamenti delle chiome e perdita anticipata delle foglie, in particolare nelle specie decidue.
Successivamente l’ondata di calore e siccità del 2024 ha causato disseccamenti e mortalità interessando anche formazioni mediterranee a leccio. Fenomeni di imbrunimento generalizzato delle chiome sono stati notati in superfici abbastanza estese nell’entroterra collinare e montano della provincia di Matera. Fenomeni ancor più estesi sono stati segnalati in alcune leccete rientranti nell’area del Parco Regionale di Gallipoli Cognato e Piccole Dolomiti Lucane soprattutto nei versanti che si affacciano sulla valle lungo il fiume Basento. Ulteriori segnalazioni sono arrivate anche dall’areale del Parco Nazionale del Pollino e lungo la valle del fiume Sinni, con molti siti interessati dal fenomeno innescatosi a seguito dell’intensa ondata di calore che ha coinvolto le aree nell’agosto del 2024, dove sono state registrate temperature elevate soprattutto durante le ore notturne.
Puglia
Già negli anni duemila, il deperimento delle querce era stato osservato soprattutto su cerro e roverella nell’Appennino meridionale ([18]), ed era stato attribuito a fattori abiotici, biotici ed antropici, questi ultimi relativi all’errata gestione dei boschi o al loro abbandono. Tra i fattori biotici coinvolti nel fenomeno sono stati identificati alcuni funghi endofiti, capaci di comportarsi da patogeni di debolezza, fra cui Diplodia corticola, B. mediterranea e Discula quercina ([7], [21], [33]).
Fattori biotici sono stati individuati anche per i casi di improvviso disseccamento del leccio nella regione Puglia. Questi fenomeni sono stati osservati per la prima volta nel 2016 nel Salento ([28]). Lo studio citato ha individuato sulle querce deperenti tre funghi patogeni appartenenti alla famiglia delle Botryosphaeriaceae, e precisamente Diplodia corticola, Diplodia quercivora e Neofusicoccum vitifusiforme. Le prime due specie sono noti agenti di cancro su fusto e rami in boschi deperienti di leccio e quercia da sughero ([46], [47], [11], [68], [30]). I funghi appartenenti alla famiglia delle Botryosphaeriaceae producono spore di facile dispersione ad opera del vento, della pioggia e degli insetti, i quali spesso peggiorano la situazione già compromessa delle querce. Buprestidi, cerambicidi e curculionidi sono parassiti emergenti della quercia nella regione mediterranea ([70]), che si aggiungono ai due fitofagi riscontrati spesso in associazione con il deperimento quercino in Puglia, gli Emitteri Kermes vermilio e Nidularia pulvinata. Non è escluso, anzi è molto probabile, che il disseccamento repentino delle querce sia dovuto a più fattori che agiscono in sinergia su piante fisiologicamente stressate dai cambiamenti climatici. Degno di nota è il rinvenimento di diverse specie patogene appartenenti al genere Phytophthora e alla famiglia delle Botryosphaeriaceae associate a gravi fenomeni di deperimento e mortalità di piante di leccio in diverse aree comunali della Provincia di Lecce ([31], [13]). Sempre nel Salento, su querce in stato di deperimento, Carluccio et al. ([19]) hanno recentemente individuato Diplodia seriata come fungo, Brenneria goodwinii, Gibsiella quercinecans e Rahnella victoriana come batteri.
Nel corso dell’estate 2024, in corrispondenza di una serie di ondate di calore estremo e siccità, si sono verificati disseccamenti estesi nelle leccete in varie zone della regione. Fra queste, si evidenziano quelle poste sui versanti calcarei esposti a sud presso il Gargano, fra Monte Sant’Angelo e San Giovanni Rotondo (Fig. 3E). Questa zona è di particolare interesse anche perché ospita un’area permanente di monitoraggio della rete di Livello I ICP Forests. Altre zone dove si sono verificati i disseccamenti sono: Acquaviva delle Fonti (BA), Gioia del Colle (BA), Massafra (TA, bosco Pianelle), Agro di Brindisi, Ostuni e comuni limitrofi, Agro di Lecce, Otranto, Nardò e comuni limitrofi, Agro di Altamura, Gravina in Puglia. Disseccamenti sono stati inoltre osservati a carico delle alberature cittadine e periurbane di Bari città e comuni limitrofi. Alla luce di queste numerose segnalazioni l’Osservatorio della Regione Puglia, con determina dirigenziale n. 86 del 24/08/2022 ha costituito un gruppo di lavoro per affrontare tale problematica.
Sardegna
I boschi a querce della Sardegna da diversi decenni sono interessati da fenomeni di deperimento, che negli ultimi anni si sono manifestati con maggiore intensità interessando superfici sempre più vaste. Il quadro sanitario delle formazioni boschive si è ulteriormente aggravato nel 2024 con il fenomeno del disseccamento, osservato in modo significativo nel settore orientale dell’Isola dalla Gallura fino al Sarrabus, e nell’area di Monte Arcosu - Gutturu Mannu (Fig. 3C). La prolungata siccità del 2024, con precipitazioni che sono state insufficienti a compensare la perdita d’acqua per evapotraspirazione, ha svolto un ruolo fondamentale nella manifestazione del fenomeno. Sebbene il disseccamento del leccio sia primariamente legato a fattori abiotici, in molte aree l’impatto della siccità è stato aggravato dalla presenza di agenti patogeni, in particolare specie appartenenti al genere Phytophthora, che compromettono l’apparato radicale delle piante colpite, riducendone la capacità di assorbire acqua e nutrienti ([43], [66]).
A seguito della diffusa e intensa manifestazione del disseccamento delle leccete nell’estate del 2024, l’Assessorato della Difesa dell’Ambiente della Regione Sardegna ha costituito un gruppo di lavoro finalizzato a studiare il fenomeno per individuarne le cause e le possibili implicazioni. Il gruppo di lavoro è stato istituito nell’ambito delle attività del Tavolo Tecnico Fitosanitario regionale e coinvolge l’Agenzia Agris, l’Università di Sassari, l’Agenzia Forestas, il Corpo Forestale e di Vigilanza Ambientale, il Dipartimento Meteoclimatico dell’Agenzia Regionale per la Protezione dell’Ambiente della Sardegna e la Protezione Civile regionale. Una delle prime attività ha riguardato l’analisi di immagini satellitari e di alcuni indici vegetazionali (EVI, NDVI e NGRDI) che hanno consentito di stimare sia l’estensione (circa 110.000 ettari) che l’intensità del disseccamento, sulla base della variazione degli indici vegetazionali rispetto allo stesso periodo dell’anno precedente. Sono stati inoltre effettuati sopralluoghi in campo che hanno permesso di verificare lo stato vegetativo delle piante colpite e in particolare del leccio. I risultati preliminari indicano una forte disomogeneità del disseccamento, anche all’interno della stessa area di rilevamento e a parità di condizioni termo-pluviometriche.
L’analisi dei dati climatici, effettuata dall’Agenzia Regionale per la Protezione dell’Ambiente della Sardegna - Dipartimento Meteoclimatico, ha evidenziato nel periodo compreso tra ottobre 2023 e luglio 2024 un sensibile aumento delle ondate di calore e un’alterazione del regime pluviometrico, con siccità prolungate soprattutto nel settore orientale dell’isola che possono aver costituito i principali fattori responsabili dei disseccamenti osservati. Tuttavia, la natura complessa del fenomeno suggerisce la necessità di un approccio olistico e multidisciplinare che consideri l’insieme dei fattori potenzialmente coinvolti: capacità idrica del suolo, variabilità della resistenza delle piante a fattori abiotici e biotici, presenza di patogeni o altre avversità.
Il fenomeno osservato nel 2024 si inserisce in una tendenza già segnalata negli ultimi decenni, ma si distingue per la rapidità e l’estensione del disseccamento. Dopo le piogge di agosto e settembre, è stata osservata una parziale ripresa vegetativa delle leccete. In particolare, nei lecci in cui il disseccamento non ha portato a morte le gemme principali, si sono osservati nuovi germogli apicali; laddove, invece, il disseccamento ha interessato anche la parte terminale dei rametti, la ripresa è avvenuta tramite germogli epicormici da gemme avventizie. È altresì importante rilevare come in diverse aree, verosimilmente già interessate da fenomeni di deperimento, è stato rilevato un tasso di mortalità del leccio intorno al 25% in seguito all’evento siccitoso.
La siccità ha avuto un impatto negativo anche sulla quercia da sughero, segnatamente nelle sugherete della bassa Gallura e dell’Ogliastra dove, contrariamente a quanto osservato sul leccio, le piante si presentavano completamente defogliate. La maggior parte delle piante di quercia da sughero ha mostrato una ripresa vegetativa nei mesi autunnali, ma nelle piante che manifestavano sintomi pregressi di deperimento l’evento siccitoso ha contribuito a peggiorarne le condizioni di salute con conseguenti disseccamenti e morìe.
La condizione fitosanitaria sempre più allarmante dei boschi esige un monitoraggio continuo delle avversità e la pianificazione di misure gestionali adeguate a mitigare gli impatti futuri. Alla luce di ciò e considerata l’importanza ecologica e socioeconomica dei boschi di querce nell’isola, la Regione Sardegna sta implementando una rete di monitoraggio fitosanitario permanente sull’intero territorio regionale.
Sicilia
In Sicilia, fenomeni estesi di disseccamento su popolamenti forestali causati da ondate di calore e prolungata carenza idrica sono stati osservati soprattutto nella parte orientale nei mesi estivi del 2024, prevalentemente su formazioni mature di Q. ilex L. e specie arbustive tipiche della macchia mediterranea e, con una minore incidenza, su individui e nuclei di Q. pubescens.
I casi più evidenti sono stati osservati in provincia di Catania dalla seconda metà del mese di luglio con la comparsa di estesi disseccamenti dei boschi di leccio, nelle aree di Monte Peloso, Monte Tre frati e Monte Minardo, a quote comprese tra 1200 e 1300 m s.l.m., nei territori di Adrano e Bronte, sul versante ovest dell’Etna (Fig. 3A-B, rispettivamente prima e durante l’evento siccitoso). Il fenomeno ha suscitato non poca preoccupazione in quanto ha coinvolto un territorio boscato di oltre 100 ha di notevole interesse naturalistico e paesaggistico, e in quanto già manifestatosi nel 2021. L’area di Monte Minardo presenta comunità vegetali e specie molto significative sotto il profilo fitogeografico ([22]).
Sintomi ascrivibili a stress di natura idrico-termica, a circa 300 m s.l.m., sono stati osservati su Q. virgiliana (Ten.) Ten., Q. congesta (C. Presl), Fraxinus ornus L. e Celtis tournefortii Lam. nel “Bosco di Santa Maria La Stella”, nel comune di Aci Sant’Antonio. In quest’ultimo, sono stati inoltre osservati casi di disseccamento di giovani piante di Q. pubescens. Situazioni similari a quelle prima descritte sono state segnalate anche nei boschetti relitti di Q. ilex di Monte Ilice (Trecastagni) e su nuclei di Q. pubescens s. l. e Q. ilex L. di Monte Ceraulo (Mascalucia), sul versante sud-est dell’Etna.
Segnalazioni di disseccamento e mortalità di un certo rilievo, ma comunque non superiori al 10%, sono state riscontrate a carico della vegetazione sclerofilla sempreverde mediterranea e sono pervenute anche dalla provincia di Siracusa. In località Mandredonne (Palazzolo Acreide) a circa 600 m. s.l.m., l’incidenza dei disseccamenti sui popolamenti di Q. ilex L. è stata del 60%. Nella stessa località, in forma sparsa e meno accentuata, anche individui di Pistacia lentiscus L. e Phyllirea angustifolia L. hanno mostrato sintomi similari. A quote più basse, all’interno della R.N.O. “Pantalica, Valle dell’Anapo e torrente Cavagrande”, sono stati rilevati disseccamenti su formazioni miste di Q. ilex L. e Q. pubescens s. l., interessando circa il 40% del soprassuolo boscato e su individui sparsi di P. angustifolia L. Infine, in località Frassino, comune di Buccheri (SR), a circa 300 m. s.l.m., formazioni a sughera hanno mostrato disseccamenti della chioma, diffusi fino al 60% del popolamento.
Nel resto dell’isola non sono stati riscontrati altri casi gravi di disseccamenti, ad eccezione di casi isolati a carico di individui di sughera nel bosco della Ficuzza e di Fagus sylvatica a Piano Battaglia (Parco regionale delle Madonie) in provincia di Palermo.
In linea generale, si può affermare che i casi più gravi sono stati riscontrati in corrispondenza di suoli di derivazione vulcanica, caratterizzati da una scarsa capacità di ritenzione idrica (Monte Etna e Monti Iblei). Nel caso della sughera, i danni sono stati osservati prevalentemente su piante recentemente decorticate. La persistente siccità associata all’aumento della frequenza delle ondate di calore, che stanno caratterizzando già da diversi anni la Sicilia, e una preliminare indagine che ha indicato l’assenza di patogeni ed insetti, fanno ipotizzare che le principali cause dei fenomeni di disseccamento e mortalità osservati siano da ascrivere principalmente allo stress di natura idrico-termica. Ulteriori indagini dovranno essere condotte per confermare l’origine abiotica del fenomeno e per verificare l’eventuale coinvolgimento di agenti patogeni nei fenomeni di disseccamento/deperimento, già evidenziati in precedenti studi ([37], [67]).
Toscana
In Toscana gli impatti delle ondate di calore e siccità hanno interessato estesamente le foreste e gli alberi dei diversi gruppi funzionali (conifere, latifoglie decidue, latifoglie sempreverdi) a partire dal 2017 ([14]). Nel corso di questi eventi climatici le latifoglie decidue (Fagus sylvatica L., Q. pubescens Willd., Q. cerris L.) abscidono le foglie e ricostituiscono la chioma più o meno completamente nell’anno successivo. Quercus cerris ha mostrato alti tassi di mortalità associata alla presenza di B. mediterranea. Le conifere, per lo più di impianto artificiale (Abies alba, Pseudotsuga menziesii, Pinus nigra), hanno mostrato sofferenza e mortalità soprattutto negli impianti al di fuori del loro optimum climatico.
Per quanto riguarda la vegetazione mediterranea, importanti ed estesi eventi di mortalità degli alberi e disseccamento di ampie parti della chioma causati da ondate di calore e siccità sono stati osservati già a partire dell’estate 2022 a carico di Q. ilex, e successivamente estesi nel corso dell’estate 2017, hanno interessato le zone costiere e nei versanti collinari esposti a mare della Toscana meridionale (Fig. 3D - [56]), interessando un fronte che, sia pure in maniera discontinua, si estende per una lunghezza di circa 130 Km, da Cecina (LI) al Parco Regionale della Maremma (GR), dove ha assunto carattere di notevole intensità ([2]). Sono state interessate in maniera sporadica e irregolare anche le aree interne, soprattutto in condizioni di suolo predisponenti allo stress idrico (suoli superficiali, rocciosi, calcarei). Sono state colpite prevalentemente le formazioni mature, provocando prima l’imbrunimento di ampie porzioni di chioma e poi la diffusa mortalità a livello della chioma e del fusto degli alberi. I lecci colpiti hanno reagito negli anni successivi per mezzo dell’attività delle gemme soppresse, producendo rami epicormici sia al colletto che all’interno della chioma. L’iniziale capacità di ricaccio di questa specie ha fatto ritenere possibile un parziale recupero vegetativo del soprassuolo danneggiato. Tuttavia, i getti prodotti fra il 2018 e il 2019 sono successivamente disseccati e morti in gran parte tra il 2021 e il 2022. La morte e il deperimento dei lecci sono accompagnati dalla diffusione del fungo patogeno di debolezza B. mediterranea. Anche i popolamenti di sughera (Q. suber L.) dell’alta Maremma toscana hanno subito un forte impatto, con numerose piante deperienti, branche disseccate e chiome fortemente diradate. Tali eventi hanno determinato nelle sughere una maggiore virulenza di patogeni, soprattutto del genere Phytophthora (oomicete), ed anche attacchi di insetti xilofagi quali Platypus cylindrus, che ne hanno accelerato il deperimento e determinato la morte.
Il disseccamento delle leccete, sia ad alto fusto che cedui invecchiati, e dei popolamenti di sughera, sta causando un cambiamento nella struttura del bosco mediterraneo, con la trasformazione del leccio ad alto fusto in forme arbustive e l’affermazione di strutture di bassa statura con prevalenza di specie arbustive della macchia mediterranea.
Le specie arbustive sempreverdi sclerofille della macchia mediterranea sono risultate essere vulnerabili, come il mirto (Myrtus communis L.) e il corbezzolo (Arbutus unedo L.). La fillirea (Phyllirea latifolia L.), considerata resistente ([16]), è apparsa anch’essa vulnerabile nel corso dei più recenti eventi di siccità. Il lentisco (Pistacia lentiscus L.) è invece risultata la specie meno danneggiata. Le specie arbustive mediterranee sono risultate particolarmente vulnerabili sui substrati ofiolitici (Monterufoli-Caselli, provincia di Pisa), dove anche Juniperus oxycedrus ha mostrato segni di deperimento.
Le segnalazioni tramite l’app SilvaCuore
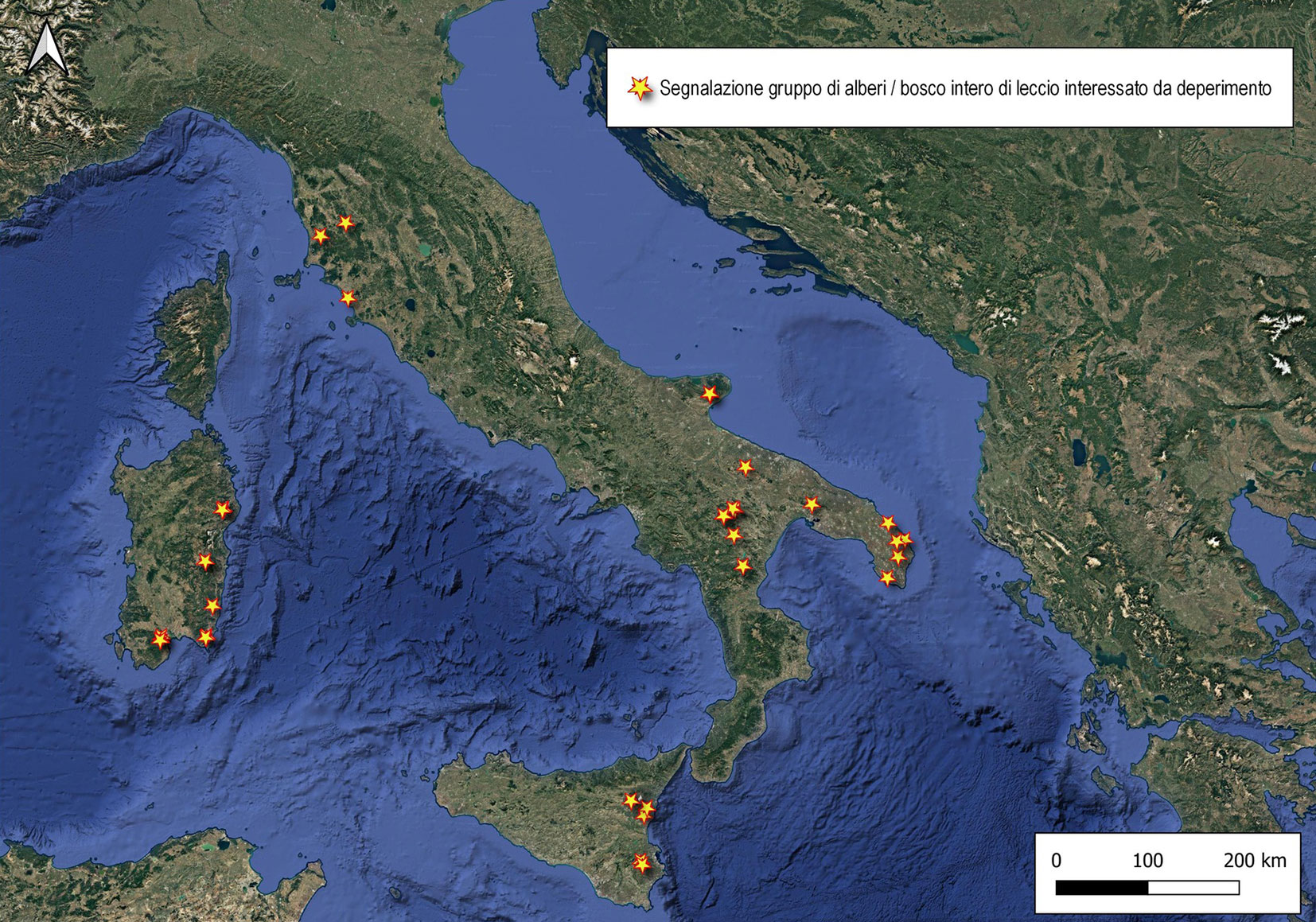
I diversi casi di deperimento e mortalità delle foreste segnalati su tutto il territorio italiano negli ultimi anni evidenziano un aumento della vulnerabilità delle foreste ai disturbi provocati da eventi climatici estremi, soprattutto ondate di calore e siccità. Questo è il presupposto che ha portato alla creazione di SilvaCuore, la prima web-application realizzata in Italia per censire lo stato di salute dei boschi. L’idea è stata quella di coinvolgere una comunità attiva di utenti, rappresentata non solo da professionisti del settore ma anche da appassionati, che vogliano contribuire attivamente al monitoraggio dei nostri boschi. Pertanto, diventa cruciale riuscire a rafforzare il legame tra il mondo della ricerca e la collettività (progetto di Citizen Science), favorendo la creazione di una banca dati condivisa e consultabile online tramite la pagina web collegata all’applicazione (⇒ https://www.silvacuore.org/). SilvaCuore, grazie ad un’interfaccia user-friendly, consente a tutti i cittadini di partecipare in modo intuitivo alla segnalazione. Il protocollo prevede l’inserimento di informazioni di base, come località, data dell’osservazione, tipologia e composizione del popolamento, specie interessata e parte della pianta coinvolta; è inoltre possibile aggiungere dati più dettagliati, quali il nome scientifico, la natura e l’entità dei sintomi, la percentuale di diffusione del fenomeno e l’eventuale presenza di alberi morti. Ogni segnalazione viene accompagnata da una o più immagini fotografiche e dalla posizione geografica, ottenuta tramite GPS integrato, posizionamento manuale su mappa o inserimento diretto delle coordinate. Tutte le informazioni confluiscono in un database dedicato (Google Firebase - Firestore®) e sono sottoposte a una verifica preliminare da remoto da parte del team SilvaCuore; nei casi più rilevanti, si procede con un sopralluogo in campo per confermare la tipologia e la gravità del fenomeno, integrando così le segnalazioni con un riscontro diretto. I dati raccolti e validati sono consultabili nella homepage dell’applicazione, dove per ogni popolamento forestale in declino sono riportati località, tipologia di problema, natura dei sintomi e una foto rappresentativa. Ad oggi, le prime segnalazioni riguardano principalmente querceti mediterranei, ma il database è in continuo aggiornamento. Le informazioni acquisite hanno già permesso di delineare un quadro preliminare della diffusione del deperimento nei boschi di leccio a livello nazionale, con 33 segnalazioni provenienti da Sardegna, Sicilia, Puglia, Basilicata e Toscana (Fig. 4).
Fig. 4 - Distribuzione geografica delle aree interessate dai fenomeni di deperimento e disseccamento del leccio, secondo le segnalazioni ricevute attraverso l’app. SilvaCuore.
L’obiettivo primario è quello di creare un network che, attraverso collaborazioni interdisciplinari con istituzioni pubbliche e/o private ed associazioni no-profit, possa fungere da strumento di supporto al monitoraggio delle foreste italiane. Tutte le informazioni raccolte, infatti, permetteranno di capire quali siano state le cause del declino e di valutare la vulnerabilità e la capacità di recupero delle formazioni forestali dopo eventi climatici estremi. SilvaCuore rappresenta un utile strumento per poter pianificare azioni di gestione che migliorino la resistenza e la resilienza delle nostre foreste: la gestione forestale adattativa, fondata su basi ecosistemiche, potrebbe infatti aumentare la possibilità delle foreste di fronteggiare lo stress climatico, al fine di conservarne e valorizzarne le funzioni primarie.
Conclusioni
Le osservazioni riportate, ancorché di tipo empirico, supportano l’estensione e la gravità del disseccamento del leccio nelle regioni dell’Italia centro-meridionale e insulare. Poiché il fenomeno appare connesso alle ondate di calore e siccità che si verificano a causa dei cambiamenti climatici, esso sembra destinato ad aggravarsi con l’aumento dell’intensità e della frequenza di tali eventi estremi, come suggeriscono i modelli climatici ([69]). Il ripetersi di questi eventi può innescare processi di degradazione della foresta verso forme di macchia e di gariga, con conseguenze economiche ed ecologiche che riguardano, fra le altre cose, la conservazione del suolo, della biodiversità e del paesaggio. Le conoscenze, tuttavia, sono ancora molto frammentarie e necessitano di un’azione coordinata fra gli enti amministrativi, enti di ricerca e gestori forestali per poter raggiungere i seguenti obiettivi:
- determinare l’estensione, l’intensità e lo sviluppo temporale degli impatti allo scopo di individuare i fattori ambientali connessi all’insorgere e alla dinamica dei danni;
- aumentare la rappresentatività della rete nazionale di monitoraggio nell’ambito della vegetazione mediterranea, tramite un’intensificazione locale della rete stessa, anche in collaborazione con enti territoriali, e l’integrazione con reti regionali di monitoraggio fitopatologico;
- promuovere il contributo dei cittadini (citizen science) tramite la diffusione dell’applicazione Silva Cuore ([20]);
- promuovere attività di monitoraggio intensivo su plot selezionati, relativamente ad aspetti ecofisiologici, fitopatologici e vegetazionali per stabilire relazioni causa-effetto;
- promuovere attività di ricerca per l’individuazione, la selezione e la propagazione di genotipi di leccio resistenti;
- l’attività di ricerca, infine, dovrà approfondire anche quali fattori stazionali e gestionali hanno influito sulla diffusione e sull’intensità del fenomeno, fondamentale per individuare buone pratiche gestionali capaci di garantire una maggiore resilienza dei soprassuoli più vulnerabili.
L’obbiettivo finale è quello di ottenere strumenti di conoscenza capaci di prevedere le dinamiche vegetazionali in atto e, in tal modo, definire le strategie di adattamento della vegetazione mediterranea per mantenere i servizi ecosistemici e l’economia su scala locale.
Ringraziamenti
Si desidera esprimere un sentito ringraziamento al personale del Servizio 13 - Servizio per il Territorio di Siracusa e a tutti i tecnici e operatori delle diverse regioni che hanno contribuito alla raccolta dei dati e delle informazioni, il cui supporto è stato determinante per la realizzazione del presente studio. Si ringraziano i colleghi dell’Agenzia Agris, dell’Università di Sassari, del CFVA Sardegna, dell’Agenzia Forestas, dell’Arpas Sardegna e della Protezione Civile Sardegna per il prezioso contributo e le competenze messe a disposizione durante i rilievi di campo e lo studio del fenomeno. Le foto sono state fornite dagli stessi autori dell’articolo. La foto 2E, relativa alla regione Puglia, è stata eseguita da drone ad opera dell’Appuntato Scelto Rocco Angelicchio del Nucleo Carabinieri Parco della Foresta Umbra.
References
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
Online | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar
CrossRef | Google Scholar